Article Text

Abstract
Background and aims Interruption of the enterohepatic circulation of bile acids (BAs) may protect against BA-mediated cholestatic liver and bile duct injury. BA sequestrants are established to treat cholestatic pruritus, but their impact on the underlying cholestasis is still unclear. We aimed to explore the therapeutic effects and mechanisms of the BA sequestrant colesevelam in a mouse model of sclerosing cholangitis.
Methods Mdr2−/− mice received colesevelam for 8 weeks. Gene expression profiles of BA homeostasis, inflammation and fibrosis were explored in liver, intestine and colon. Hepatic and faecal BA profiles and gut microbiome were analysed. Glucagon-like peptide 1 (GLP-1) levels in portal blood were measured by ELISA. Furthermore, Mdr2−/− mice as well as wild-type 3,5-diethoxy-carbonyl-1,4-dihydrocollidine-fed mice were treated with GLP-1-receptor agonist exendin-4 for 2 weeks prior to analysis.
Results Colesevelam reduced serum liver enzymes, BAs and expression of proinflammatory and profibrogenic markers. Faecal BA profiling revealed increased levels of secondary BAs after resin treatment, while hepatic and biliary BA composition showed a shift towards more hydrophilic BAs. Colonic GLP-1 secretion, portal venous GLP-1 levels and intestinal messenger RNA expression of gut hormone Proglucagon were increased, while ileal Fgf15 expression was abolished by colesevelam. Exendin-4 treatment increased bile duct mass without promoting a reactive cholangiocyte phenotype in mouse models of sclerosing cholangitis. Microbiota analysis showed an increase of the phylum δ-Proteobacteria after colesevelam treatment and a shift within the phyla Firmicutes from Clostridiales to Lactobacillus.
Conclusion Colesevelam increases faecal BA excretion and enhances BA conversion towards secondary BAs, thereby stimulating secretion of GLP-1 from enteroendocrine L-cells and attenuates liver and bile duct injury in Mdr2−/− mice.
- primary sclerosing cholangitis
- bile acid metabolism
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Bile acid (BA) sequestrants (resins) are used to treat cholestatic pruritus; in single cases, improvement of cholestasis was observed.
Resin-bound BAs stimulate the G-protein-coupled BA receptor Takeda G protein-coupled receptor 5 (TGR5) which increases glucagon-like peptide 1 (GLP-1) secretion.
GLP-1 signalling was shown to stimulate cholangiocyte proliferation and to protect bile duct epithelia against apoptosis.
What are the new findings?
Colesevelam treatment improved hepatic inflammation, fibrosis and ductular proliferation in the Mdr2−/− mouse model of sclerosing cholangitis.
Colesevelam feeding reduced colonic farnesoid X receptor messenger RNA and protein expression in Mdr2−/− mice.
Colesevelam feeding result in detoxification of the hepatic as well as the biliary BA composition.
Colesevelam feeding changed faecal BA profiles towards TGR5 signalling favouring increased colonic expression of GLP-1 and increasing its levels in portal blood, potentially contributing to the improvement of cholestatic liver injury in Mdr2−/− mice.
How might it impact on clinical practice in the foreseeable future?
Beyond their current use in pruritus BA sequestrants (with a longstanding and excellent safety track record) may represent a cheap and readily available treatment strategy for underlying cholestatic liver injury.
Introduction
Bile acid (BA) sequestrants such as colesevelam are anion exchange resins that complex BAs in the intestinal lumen, thus decreasing BA reuptake and enterohepatic circulation. BA sequestrants have been primarily used for lowering serum cholesterol and treatment of cholestasis-associated pruritus1 as well as BA-induced diarrhoea.2 Interestingly, colesevelam has been reported to improve serum BA levels3 and colestyramine to improve cholestatic liver enzymes in patients with primary sclerosing cholangitis (PSC),4 suggesting potential anticholestatic and bile duct-protective properties. Resins stimulate BA signalling via the Takeda G protein-coupled receptor 5 (TGR5)5 while repressing colonic farnesoid X receptor (FXR),6 thereby promoting secretion of glucagon-like peptide 1 (GLP-1) from enteroendocrine L-cells.6 Notably, the GLP-1 analogue exendin-4 has also cholangioprotective effects in primary cholangiocytes as well as in bile duct ligated rats,7 8 in addition to its beneficial metabolic effects.9 Although both inhibitions of apical sodium-dependent bile salt transporter (ASBT) as well as administration of BA sequestrants result in interruption of enterohepatic circulation of BAs and improvement of cholestatic features,4 10 11 their mechanism of action may differ. In contrast to complex binding of BAs through resins, the recently developed inhibitors of ASBT enhance the excretion of unbound BAs into intestine, where they can signal intracellularly via FXR, thereby inhibiting GLP-1 expression/secretion.6 We hypothesise that BA resin treatment results in reduced liver and bile duct injury in the Mdr2−/− mouse (as a model of sclerosing cholangitis) by modulating faecal as well as biliary BA content and composition, favouring formation of endogenous BAs that stimulate TGR5, which in turn induces cholangioprotective GLP-1 production.
Materials and methods
Animals
FVB/N wild-type (WT) and Mdr2−/− mice obtained from Jackson Laboratory (Bar Habor, Maine, USA) were housed in a 12 hours light/dark house facility with water and standard chow diet (SSNIFF, Soest, Germany) ad libitum.
Feeding protocols
Colesevelam (BA sequestrant) was a generous gift from Dr Alan F Hofmann. Eight weeks old male Mdr2−/ − mice received either control diet or a diet supplemented with 2% (w/w) colesevelam for 8 weeks.
In another series, Mdr2−/ − mice were injected once daily with exendin-4 (Sigma-Aldrich, Vienna, Austria) (0.1 µg/kg) for 2 weeks. Eight-week-old male FVB/N WT mice receiving 0.1% 3,5-diethoxycarbonyl-1,4-dihydrocollidine (DDC) for 17 days were injected with exendin-4 (0.1 µg/kg) once daily. The experimental protocols were approved by the local Animal Care and Use Committee (BMWFW-66.009/0315- WF/3b/2014).
Routine serum biochemistry and histology
Serum biochemistry and histological staining was performed as described previously.10
Immunohistochemistry
Detection of hepatic cytokeratin (CK) 19, F4/80, Ki67, vascular cell adhesion protein 1 (VCAM-1) and osteopontin (OPN) was performed as described previously.12 13 Immunohistochemistry (IHC) was also performed for GLP-1 in colon slices. In Mdr2−/− mice, CK19, VCAM-1, OPN and GLP-1 IHC positive areas were quantified by ImageJ software V.1.47v (National Institute of Health). In WT DDC-fed mice, gene expression profiles were used for quantification of these markers since DDC feeding results in accumulation of protoporphyrin plaques in bile ducts that give false positive areas with computational quantification. Ki67 positive cells were counted manually in five different power fields in all experimental settings.
Hepatic hydroxyproline content
To quantify liver fibrosis, hepatic hydroxyproline was measured from a standardised liver lobe as described previously.14
Hepatic, biliary and faecal bile acid analysis
Hepatic, biliary and faecal BA profiles were acquired using ultraperformance liquid chromatography tandem mass spectrometry as described previously.15
16 s rRNA microbiome analysis
16 s rRNA microbiome analysis was performed as described previously.16
Messenger RNA analysis and PCR
RNA isolation from liver and intestine, complementary DNA synthesis and real-time PCR were performed as described previously.17 Oligonucleotide sequences are available on request.
Western blot analysis
Protein expression was quantified as described previously.18
GLP-1 ELISA
Blood was collected from portal vein. The GLP-1 (Active 7–36) ELISA (Catalogue Number: 43-GP1HU-E01, ALPCO) was conducted according to the manufacturer’s instructions.
Statistical analysis
Results were evaluated using SPSS V.20.0. Statistical analysis was performed using student`s t-test. Data were reported as means of five to seven animals per group±SD A p value ≤0.05 was considered significant.
Results
Colesevelam treatment improves cholestatic liver injury in Mdr2−/− mice
Colesevelam was well tolerated in Mdr2−/− mice without affecting animal behaviour, food intake or body weight after 8 weeks of administration (see online supplementary figure 1). Importantly, mice did not develop diarrhoea. Colesevelam treatment even reduced the daily stool volume from 1.52 g/mouse/day to 1.03 g/mouse/day (cumulative output). Liver histology revealed complete reversion of bile duct injury (pericholangitis and onion skin type of biliary fibrosis) in colesevelam-treated Mdr2−/− mice (figure 1A, all mice shown in online supplementary figure 3). Additional experimental series confirmed that colesevelam treatment in Mdr2−/− mice led to complete reversion of the liver phenotype similar to that of WT mice (online supplementary figure 2). Accordingly, serum parameters ALT, AST and BAs as well as liver to body weight ratios of treated Mdr2−/− mice were in the same range as in WT mice (figure 1 and online supplementary figure 2). Importantly, resin treatment reduced bile duct proliferation as determined by immunohistochemistry (IHC) (figure 2A) and messenger RNA (mRNA) expression of CK 19 (figure 2B). Together, these findings implicate a beneficial role of BA sequestrant feeding on cholestatic liver and bile duct injury in a mouse model of sclerosing cholangitis (figures 1 and 2).
Supplemental material
Supplemental material
Supplemental material

Colesevelam feeding improves liver injury in Mdr2−/− mice. (A) Representative H&E images (10× magnification) with markedly improved liver histology with reduced pericholangitis and onion skin type of biliary fibrosis in colesevelam fed Mdr2−/− mice. (B) Serum biochemistry reflects improved levels of transaminases (ALT, AST) as well as AP and bile acids (BAs). (C) Liver to body weight ratio (LW/BW) improved due to colesevelam feeding. *Significant difference from Mdr2−/− control group; p<0.05. ALT, alanine aminotransferase; AP, alkaline phosphatse; AST, aspartate aminotransferase; BA, bile acid.

Ductular proliferation is improved by colesevelam treatment. (A) Representative CK19 immunohistochemistry pictures (10× magnification) show reduced cholangiocyte proliferation in colesevelam-treated Mdr2−/− mice. (B) Real-time PCR was used to assess the mRNA expression of cholangiocyte proliferation marker CK19 which was reduced by colesevelam feeding. Expression values were normalised against 18sRNA levels and are shown relative to expression level in Mdr2−/− control mice. *Significant difference from Mdr2−/− control group; p<0.05. mRNA, messenger RNA.
Colesevelam treatment attenuates hepatic inflammation and fibrosis in Mdr2−/− mice
Because the reactive cholangiocyte phenotype in cholangiopathies is associated with development of hepatic inflammation and fibrosis, markers for both key processes in the progression of liver injury were investigated (figures 3 and 4). Representative images of F4/80 IHC displayed reduced numbers of macrophages in livers of Mdr2−/− mice fed with colesevelam (figure 3A). Accordingly, F4/80 mRNA levels were reduced in the treatment group in comparison to Mdr2−/− control mice (figure 3B). Gene expression profiles of Mcp1 and Vcam-1 (two proinflammatory markers of liver injury in Mdr2−/− mice) also revealed profound reduction of these genes in colesevelam fed mice (figure 3B). Sirius red staining (figure 4A) displayed a marked reduction of fibrosis in Mdr2−/− mice subjected to colesevelam. Accordingly, transcription of profibrogenic genes such as Col1a1 and Col1a2 were considerably reduced in the treatment group (figure 4B). In line, hepatic hydroxyproline content as well as α-SMA protein levels were attenuated in the colesevelam fed Mdr2−/− mice (figure 4C and online supporting figure 4A).
Supplemental material

Colesevelam feeding improves hepatic inflammation in Mdr2−/− mice. (A) Representative F4/80 immunohistochemistry images (10× magnification) showing reduced numbers of macrophages in the livers of Mdr2−/− mice after colesevelam treatment. (B) Real-time PCR was used to assess the mRNA expression of F4/80, Mcp1 and Vcam-1 as markers of inflammation which were reduced due to treatment with colesevelam. Expression values were normalised against 18 sRNA levels and are shown relative to expression level in Mdr2−/− controls. * Significant difference from Mdr2−/− control group; p<0.05.

Biliary fibrosis is reduced in Mdr2−/− mice by colesevelam treatment. (A) Representative sirius red stainings (10× magnification) show improved biliary fibrosis in Mdr2−/− mice fed with colesevelam. (B) Real-time PCR was used to assess the mRNA expression of fibrotic markers Col1a1 and Col1a2 which were reduced by colesevelam feeding. Expression values were normalised against 18sRNA levels and are shown relative to expression level in Mdr2−/− controls. (C) OH-proline levels were assessed and turned out to be improved in Mdr2−/− mice subjected to colesevelam treatment. *Significant difference from Mdr2−/− control group; p<0.05.
Colesevelam feeding alters bile acid homeostasis in Mdr2−/− mice
In line with effective BA depletion, colesevelam feeding reduced hepatic FXR protein expression (figure 5A) and increased mRNA expression of Cyp7a1 (the rate limiting enzyme in BA synthesis) sevenfold in Mdr2−/− mice (figure 5B). Notably, the sinusoidal BA uptake machinery Na+/taurocholate cotransporting polypeptide was increased at protein level by colesevelam feeding while expression of the canalicular BA export pump bile salt export pump did not differ between the groups (see online supporting figure 4B). Ileal mRNA expression of Fxrα (figure 5C), its target hormone Fgf15 and Proglucagon was also assessed (figure 5D). Ileal Fxrα mRNA expression was reduced by colesevelam and in line, Fgf15 mRNA was completely abolished under resin treatment, while Proglucagon was induced fivefold (figure 5D), indicating that colesevelam feeding may have differential impact on FXR and TGR5-dependent regulation of gene expression. Notably, gene expression of the inflammatory marker Mcp1 was reduced in the terminal ileum of colesevelam fed Mdr2−/− mice (figure 5D). Of particular interest, Proglucagon mRNA expression is also increased in the colon of colesevelam-treated Mdr2−/− mice (figure 5E).

Colesevelam feeding impacts on BA homeostasis in Mdr2−/− mice. (A) Hepatic FXR protein expression is reduced in colesevelam fed Mdr2−/− mice (Histone 3 was used as internal control), while (B) Cyp7a1 (key enzyme for BA synthesis) mRNA expression is profoundly increased. (C) Ileal Fxrα mRNA expression is reduced in colesevelam-fed Mdr2−/− mice. (D) Ileal gene expression of Fgf15 was completely abolished in colesevelam-fed mice, while Proglucagon levels were induced fivefold. Expression of inflammatory marker Mcp1 is reduced in the terminal ileum of colesevelam-fed Mdr2−/− mice. (E) Colonic gene expression of Proglucagon was increased due to colesevelam feeding. Expression values were normalised against 18sRNA (liver) and Gapdh (ileum) and 18sRNA (colon) levels and are shown relative to expression level in Mdr2−/− controls. *Significant difference from Mdr2−/− control group; p<0.05. BA, bile acid; FXR, farnesoid X receptor.
Bile flow as well as biliary HCO3 - and BA output were not influenced by colesevelam feeding (figure 6A-C). Rather, hepatic and biliary BA concentrations increased, 8-fold and 1.4-fold (data not shown), respectively, while their profiles showed a shift towards a more hydrophilic BA composition with taurine-conjugated muricholic acids (MCAs) consisting 51% and 56%, respectively (figure 6D and E).

Colesevelam treatment does not influence bile flow but changes bile composition. (A) Hepatobiliary bile flow (B) biliary HCO3 - output and (C) biliary BA output remain unchanged under colesevelam treatment. (D) Hepatic and (E) biliary BA profile showed a shift towards a more hydrophilic BA composition with taurine-conjugated MCAs being the most prominent species. BA, bile acid; MCA, muricholic acid; TCA, taurine-conjugated cholic acid; TCDCA, taurine-conjugated deoxycholic acid; TαMCA, tauro-alpha-muricholic acid; TβMCA, tauro-beta-muricholic acid; T ω MCA, tauro-omega-muricholic acid; TUDCA, taurine-conjugated ursodeoxycholic acid.
Colesevelam treatment modifies faecal BA output and composition
Resin feeding profoundly increased faecal BA output (1276.8 µmol/day in Mdr2−/− control mice and 6656.3 µmol/day in Mdr2−/− colesevelam-fed mice; cumulative faeces output). Importantly, in spite of a fivefold increase in faecal BA output Mdr2−/− mice subjected to colesevelam did not develop diarrhoea. At the same time, the resin significantly altered faecal BA composition by increasing concentrations of primary BAs α/βMCA, cholic acid (CA) and chenodeoxycholic acid as well as of secondary BAs lithocholic acid (LCA) and deoxycholic acid (DCA) (table 1). Of note, the two most potent endogenous TGR5 ligands, LCA and DCA,19 represented 51% of total BAs in the faeces of colesevelam fed Mdr2−/− mice, compared with 19% in the untreated group. Moreover, colesevelam-fed Mdr2−/− mice had 91% unconjugated BAs in the faeces compared with 83% in Mdr2−/− control mice (table 1).
Faecal bile acid levels
Colesevelam treatment modifies gut microbiome
16srRNA microbiome analysis revealed an increase in the phylum Proteobacteria in Mdr2−/− mice treated with colesevelam (9% vs 5% in the control group) (see online supplementary figure 5A). More specifically, this increase resulted from elevated abundance of the class δ-Proteobacteria (order Desulfofibronales, family Desulfovibronacae) (see online supplementary figure 5B). Despite unchanged Firmicutes levels (Mdr2−/− control group 38%; Mdr2−/− colesevelam group 36%), in the colesevelam fed group a shift within the phylum Firmicutes was seen, from the class Clostridiales to the class Bacilli, more specifically from the order Clostridiales to the order Lactobacillales, known to consist several species exerting bile salt hydrolase (BSH) activity, such as Lactobacillus 20 (see online supplementary figure 5B). Also the phylum Bacteriodetes remained unchanged in Mdr2−/− control and colesevelam fed mice (56% vs 54%) (see online supplementary figure 5A). Of particular interest, although we see increased levels of secondary BAs in the faeces of colesevelam-fed Mdr2−/− mice, the class Clostridia (containing bacteria exhibiting 7α dehydroxylase activity) was predominant in untreated Mdr2−/− control mice (see online supplementary figure 5B).
Supplemental material
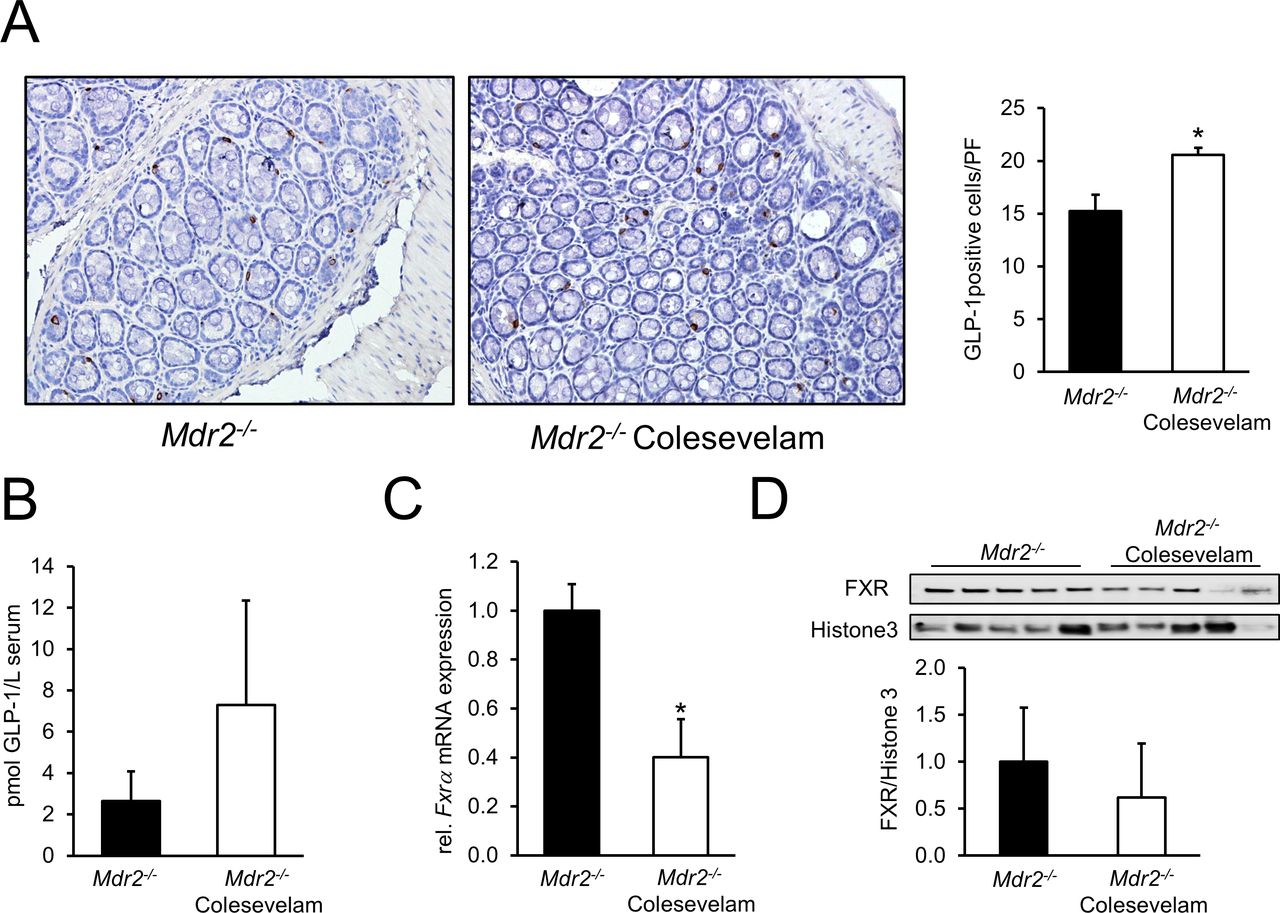
Colesevelam treatment increases colonic GLP-1 signalling in Mdr2−/− mice
Since increased mRNA levels of small intestinal and colonic Proglucagon and elevated secondary BAs LCA and DCA in colesevelam-fed Mdr2−/− mice point towards an augmented TGR5–GLP-1 signalling, colonic GLP-1 expression was assessed by IHC. Mdr2−/− mice fed with colesevelam exhibited increased numbers of GLP-1 secreting enteroendocrine L-cells (figure 7A). Accordingly, GLP-1 concentrations in portal blood of BA resin fed animals tended to be increased (figure 7B). Importantly (given the differential impact of FXR vs TGR5 on GLP-1 secretion), mRNA (figure 7C) as well as protein expression levels of (proximal) colonic FXR were reduced in colesevelam-fed Mdr2−/− mice compared with controls (figure 7D). Also, the expression of the FXR downstream target Ibabp was reduced at mRNA level (data not shown).

![[SP1.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC1/embed/inline-supplementary-material-1.jpg?download=true){kind=link}
![[SP2.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC2/embed/inline-supplementary-material-2.jpg?download=true){kind=link}
![[SP3.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC3/embed/inline-supplementary-material-3.jpg?download=true){kind=link}
{kind=link}
{kind=link}
![[SP4.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC4/embed/inline-supplementary-material-4.jpg?download=true){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![[SP5.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC5/embed/inline-supplementary-material-5.jpg?download=true){kind=link}
{kind=link}
Colesevelam treatment modulates intestinal GLP-1 secretion. (A) Representative GLP-1 IHC images (10× magnification) reflect increased numbers of GLP-1 secreting enteroendocrine L-cells in the colon of Mdr2−/− mice fed with colesevelam. (B) GLP-1 levels in portal blood were measured by ELISA as described in material and method section. GLP-1 concentration tended to be increased in the colesevelam-fed animals. (C) Colonic Fxrα mRNA expression is significantly reduced in colesevelam-treated Mdr2−/− mice. (D) Colonic protein expression of FXR was reduced in Mdr2−/− mice by colesevelam treatment. *Signficant difference from Mdr2−/− control group; p<0.05. GLP-1, glucagon-like peptide 1 ; mRNA, messenger RNA.
GLP-1 receptor agonist exendin-4 promotes bile duct formation
To further address a potential role of GLP-1 in improvement of liver and biliary injury,7 8 we treated Mdr2−/− mice and WT mice fed with DDC13 (as mouse models of sclerosing cholangitis) with exendin-4 as GLP-1 receptor agonist.7 In both models, exendin-4 increased expression of CK19 and Ki67 (see online supplementary figures 6 and 8), while at the same time VCAM-1 and OPN remained unchanged (see online supplementary figures 7 and 9).
Discussion
In this study, we uncover that the BA binding resin colesevelam reverses cholestatic liver and bile duct injury in the Mdr2−/− mouse model of sclerosing cholangitis (figure 1 and online supporting figures 2 and 3). Our findings provide strong evidence that resins such as colesevelam may have important implications as potential therapeutic strategies for cholestatic liver disease beyond their role in the treatment of pruritus. In this study, we now provide a potential mechanistic explanation for anecdotal clinical observations that resins may in addition to itch also improve cholestasis.1 Importantly, these mechanisms appear to be different from more costly and side effect prone ASBT inhibitors that recently were propagated for the treatment of cholestatic pruritus.21 22
The improvement of the cholangiopathy in the Mdr2−/− mouse subjected to colesevelam may at least in part be explained by interruption of the enterohepatic circulation with reduced output of potentially toxic BAs.10 11 Importantly, faecal BA analysis showed profound increases in BA excretion due to colesevelam treatment and enhanced conversion of primary to secondary BAs (table 1) with subsequent effects on BA signalling in the gut. As such, we observed markedly increased GLP-1 expression in enteroendocrine L-cells (figure 7A) and elevated GLP-1 levels in portal blood of colesevelam fed Mdr2−/− mice (figure 7B), pointing towards increased TGR5 activity as a result of increased formation of secondary BAs. Notably, enteroendocrine L-cell FXR activity and TGR5-mediated GLP-1 secretion appear to be inversely related.5 6 Mice lacking FXR showed elevated levels of Proglucagon mRNA and elevated serum GLP-1 levels.6 Moreover, colesevelam treatment of ob/ob mice was found to inhibit Fgf15 mRNA expression in ileum (further arguing for reduced FXR activity) and increased colonic Proglucagon expression.6 In line, in our study small intestinal and colonic FXR mRNA and protein expression was reduced in Mdr2−/− mice subjected to colesevelam feeding while at the same time colonic GLP-1 expression and portal blood levels of this hormone were increased (figures 5 and 7).
GLP-1 released from the colon, entering the liver via portal blood, may bind to its receptor at cholangiocytes and modulate their adaptive response to cholestasis, by reducing cholangiocyte apoptosis while at the same time favouring their proliferation, thereby preventing ductopaenia, a unifying event in cholangiopathies.7 8 23 This phenomenon would be predicted to protect cholangiocytes from BA-induced stress. GLP-1 docking to its receptor increases cAMP production, which results in an anti-inflammatory response due to the inhibition of NFκB and STAT1 expression, thereby reducing expression of inflammatory cytokines and chemokines as well as adhesion molecules such as Tnfα, Vcam-1 and Mcp1,24 known to contribute the reactive cholangiocyte phenotype.25 This hypothesis is at least partially supported by our findings in Mdr2−/− mice as well as in DDC-fed WT mice which were injected with the GLP-1 receptor agonist exendin-4. In both mouse models, bile duct mass (CK19 expression and staining) as well as cell proliferation (Ki67 staining) were increased due to exendin-4 injection while at the same time, markers for the reactive (proinflammatory/profibrogenic) cholangiocyte phenotype (VCAM-1 and OPN) remained unchanged (see onlinesupporting figures 6-9). Stimulation of cholangiocyte proliferation without promotion of a reactive cholangiocyte phenotype could be viewed as a potentially protective mechanism counteracting ductopaenia.
In addition to increased GLP-1 signalling, also changes in hepatic and biliary BA composition due to colesevelam feeding may add to the overall beneficial effect seen in livers of resin-treated Mdr2−/− mice. As such, colesevelam feeding led to a ‘hydrophilisation’ (thereby detoxification) of hepatic as well as biliary BA composition with predominance of taurine-conjugated α-MCAs, β-MCAs and ω-MCAs (figure 6). Interestingly, this ‘hydrophilisation’ was not seen in Mdr2−/− mice treated with an ASBT inhibitor,10 as reflected by the negligible concentration of biliary hydrophilic muricholic acid species while concentration of hydrophobic TDCA was increased.10 In addition to differences of GLP-1 signalling between ASBT inhibition and resin feeding, these divergent effects on biliary BA composition may significantly contribute to the mechanistic differences between these two treatment strategies.
The divergent effects on GLP-1 secretion might be explained by the fact that in ASBT inhibition, largely unmodified BAs enter the colon which can be reabsorbed and then activate intracellular FXR-dependent signalling pathways reported to suppress GLP-1 expression.6 Resin-bound BAs cannot be reabsorbed to the same extent and thus are still able to signal via the G protein-coupled receptor TGR5, thereby stimulating GLP-1 secretion. In line with this hypothesis, we found reduced colonic FXR expression in colesevelam-treated Mdr2−/− mice (figure 7C,D), which could contribute to increased GLP-1 expression,6 in addition to stimulation via TGR5.26 Therefore, in the context of cholestatic liver disease, inhibiting FXR and stimulating TGR5 in the colon seems to have beneficial effects. Interestingly, colonic FXR expression remained unaffected in Mdr2−/− mice treated with the ASBT inhibitor A4250 (data not shown). Together, these observations suggest that changes in colonic BA signalling are specific for the BA resin therapy in Mdr2−/− mice and explained at least in part the more pronounced improvement of cholestatic liver injury in Mdr2−/− mice in comparison to reported effects of ileal ASBT inhibitors.10 11
Since BAs are potential regulators of microbiota, we performed a 16S rRNA analysis to determine microbiota composition. Although elevated concentrations of secondary BAs may implicate a shift towards the phylum Firmicutes (since they include bacteria responsible for secondary BA production27), the abundance of Firmicutes remained unchanged under colesevelam treatment (see online supplementary figure 5A) and the class Clostridia (which contains bacteria exhibiting 7α-dehydroxylase activity) was even predominant in the untreated Mdr2−/− control group (see online supplementary figure 5B). However, 7α-dehydroxylation is restricted to taurine-deamidated BAs. Indeed, primary BAs need to be deconjugated before transformation to secondary BAs.28 In line, Mdr2−/− mice fed with colesevelam have more unconjugated BAs in faeces than Mdr2−/− control mice (Mdr2−/− control mice 83% versus Mdr2−/− mice fed with colesevelam 91%). Interestingly, the family Lactobacillaceae, containing the genus Lactobacillus which is known to exert BSH activity20 was predominant in colesevelam-treated Mdr2−/− mice (see online supporting figure 5B). Furthermore, increased BA deconjugation may also explain the elevated amount of the class δ-Proteobacteria which consists of bacteria having desulfatation properties, since taurine contains a sulfonic moiety,20 which might represent a growth factor for these bacteria. In this context, δ-Proteobacteria produce hydrogen sulfide which was shown to have anti-inflammatory properties since it is relevant for the release of trefoil factor 3, which may play a key role in mucosal regeneration and repair processes,29 although this is discussed controversially.30 Another reason for increased concentration of faecal secondary BAs could be a prolonged transit time of resin-bound BAs in the colon, which would provide the bacteria more time for metabolism of BAs.
Extending the use of BA sequestrants that have a longstanding and excellent safety track record may represent a cheap and readily available approach in the treatment of cholestatic liver injury beyond pruritus. Cholestyramine has been shown to improve liver enzymes in PSC patients.4 Furthermore, BA sequestrants are the first-line treatment for BA-induced diarrhoea,31 a side effect that was seen both in phase I and phase II clinical trials22 with ASBT inhibitors.21 While ’older resins' formulated as powders were poorly tolerated because of considerably large doses and poor taste, the newer compounds can be formulated as pills or gels which may improve patients acceptance and compliance.
In conclusion, our study demonstrates that interruption of the enterohepatic circulation of BAs with colesevelam improves sclerosing cholangitis in the Mdr2−/− mouse by altering hepatic and biliary BA composition towards a more hydrophilic configuration. Moreover increasing faecal BA excretion and modulating the faecal BA composition towards more abundant TGR5 ligands results in increased colonic levels of GLP-1. Therefore, our results suggest that reduction of ileal BA reuptake by BA sequestrants may open new perspectives for pharmacological treatment of patients with cholangiopathies such as PSC.
Supplemental material
![[SP6.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC6/embed/inline-supplementary-material-6.jpg?download=true){kind=link}
Supplemental material
![[SP7.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC7/embed/inline-supplementary-material-7.jpg?download=true){kind=link}
Supplemental material
![[SP8.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC8/embed/inline-supplementary-material-8.jpg?download=true){kind=link}
Supplemental material
![[SP9.jpg]](https://gut.bmj.com/content/gutjnl/67/9/1683/DC9/embed/inline-supplementary-material-9.jpg?download=true){kind=link}
Supplemental material
References
Footnotes
Contributors CDF: study concept and design, writing of the manuscript, data collection, statistical analysis and interpretation of data. EH, VM, VK, AW, MS, NL, TS, AT: data collection, critical revision of the manuscript for important intellectual content. GP, KK, HUM: critical revision of the manuscript for important intellectual content. MT: study concept and design, interpretation of data, outlining and revising the manuscript; study oversight and acquisition of project funding.
Funding This study was funded by Austrian Science Fund. Grant number: F3517-B20.
Competing interests MT served as a consultant for Albireo, Falk, Genfit, Gilead, Intercept, MSD, Novartis and Phenex and is a member of the speakers’ bureau of Falk, Gilead, MSD and Roche. He further received travel grants from Falk, Roche and Gilead and unrestricted research grants from Albireo, Falk, Gilead, Intercept, MSD and Takeda. He is also coinventor of a patent on the medical use of norUDCA. H-UM served as consultant for Albireo, AstraZeneca and Intercept and received travel grants from Falk. CDF received travel grants from Gilead, Roche, Falk, Merck, Vifor, Abbvie and Böhringer Ingelheim. EH received travel grants from Falk, Roche and MSD. She also served as a consultant for Novartis and Intercept. AW consulted for Intercept and received travel grants from Falk.
Patient consent Not required.
Provenance and peer review Not commissioned; externally peer reviewed.