Article Text

Abstract
Background and aims: The mechanism of transformation to intestinal metaplasia in Barrett’s oesophagus has not been clarified. We previously reported that bile acids activate the Cdx2 promoter via nuclear factor kappa B (NF-κB) and stimulate production of Cdx2 protein in oesophageal keratinocytes with a resulting production of intestinal type mucin. In addition to Cdx2, Cdx1 may play an important role in the development of Barrett’s oesophagus. Therefore, we studied the direct effects of bile acids on the expression of Cdx1 as well as the precise mechanisms of Cdx1 expression in cultured oesophageal squamous epithelial cells. Furthermore, we investigated the relationship between Cdx1 and Cdx2 expression in cultured oesophageal squamous epithelial cells.
Methods: A rat model of Barrett’s oesophagus was produced by anastomosing the oesophagus and jejunum. The expression of Cdx1 was investigated by immunohistochemistry, while the response of that expression to bile acids was studied using a Cdx1 promoter luciferase assay. In addition, oesophageal squamous epithelial cells were transfected with a Cdx1 or Cdx2 expression vector, after which their possible transformation to intestinal-type epithelial cells was investigated.
Results: In our Barrett’s rat model, the metaplastic epithelium and adjoining squamous epithelium strongly expressed Cdx1. Further, the bile acids mixture dose-dependently increased Cdx1 promoter activity and Cdx1 protein in oesophageal epithelial cells. Transfection of the Cdx1 expression vector in cultured oesophageal epithelial cells induced production of Cdx2 protein.
Conclusion: Bile acid-induced sequential expression of Cdx1 followed by Cdx2 may have an important role in the development of Barrett’s epithelium.
Statistics from Altmetric.com
Barrett’s oesophagus is an acquired condition in which squamous epithelium in the distal oesophagus is replaced by metaplastic columnar epithelium.1 Interestingly, Barrett’s oesophagus frequently develops into adenocarcinoma of the oesophagus, a type of cancer whose incidence has increased during the past decade.2 Barrett’s oesophagus is considered to be induced by chronic gastro-oesophageal reflux disease (GORD),3 during which the oesophageal epithelium attempts to adapt to its new environment and may undergo profound phenotypic changes, leading to a different type of epithelium that is more resistant to this novel environment.4 Several clinical studies have suggested that reflux of duodenal contents with bile acids contributes to the development of Barrett’s oesophagus.5–8 Although there is great interest in the pathogenesis of this condition, little is known regarding the mechanism of cellular metaplasia or precise cell origin of Barrett’s epithelium.
Cdx1 and Cdx2 are members of the caudal-related homeobox gene family, based on their sequence homology to the caudal gene of Drosophila melanogaster.9 10 They are intestine-specific transcription factors, and are important in the early differentiation and maintenance of intestinal epithelial cells during gastrointestinal development.11 Cdx2 protein has been found in Barrett’s epithelium and oesophageal epithelium with oesophagitis.12 13 We previously reported a two-step mechanism that is involved in the development of Barrett’s epithelium, in which bile acids activate the Cdx2 promoter via nuclear factor kappa B (NF-κB) and stimulate the production of Cdx2 protein in oesophageal keratinocytes, with a resulting production of intestinal type mucin.14 In addition to Cdx2, Cdx1 is also known to play an important role in the development of intestinal metaplasia.15 When expressed in the stomach, Cdx1 can induce differentiation of gastric epithelial cells to intestinal-type cells, suggesting that it may play a fundamental role in generating intestinal metaplasia.16 Further, Cdx1 protein has also been found in Barrett’s epithelium.15 17
In the present study, we investigated the stimulatory effects of bile acids on Cdx1 expression in oesophageal epithelial cells. Further, we investigated the complicated mechanisms related to Cdx1 and Cdx2 in the development of Barrett’s epithelium.
MATERIALS AND METHODS
Protein extraction and western blot analysis
Protein extraction and western blot analysis were performed as described previously.14 The membranes were incubated with anti-Cdx1 (1:100), anti-Cdx2 (1:100; BioGenex, San Ramon, California, USA), anti-MUC2 (1:100; Santa Cruz Biotechnology, Santa Cruz, California, USA), or anti-β-actin (1:3000; Sigma Chemical, St Louis, Missouri, USA) antibodies, followed by horseradish-peroxidase-conjugated anti-rabbit or anti-goat immunoglobulin (DAKO, Carpinteria, California, USA).
Rat models of Barrett’s oesophagus
Seven-week-old male Wistar rats were used for the experiments. To induce Barrett’s oesophagus, we employed Levrat’s model with minor modifications, as described previously.14 18–20 In brief, the gastro-oesophageal junction was cut and the oesophageal end separated. The distal end of the oesophagus was then re-implanted 2 cm beyond the ligament of Treitz in an end-to-side fashion into a loop of the jejunum and the proximal end of the stomach was ligated. Two and 6 months after formation of the oesophageal–jejunal anastomoses, the rats were euthanised and the oesophagi removed.
Immunohistochemistry
Five-micrometre thick sections from each specimen were stained with haematoxylin–eosin for light microscopy examinations. To identify Cdx1- or Cdx2-expressing cells, the tissue sections were incubated with anti-Cdx1 (1:100) or anti-Cdx2 (1:100) antibodies, followed by incubation with secondary biotinylated anti-mouse immunoglobulin or anti-rabbit immunoglobulin (DAKO). Bound antibodies were detected using an avidin–biotin peroxidase method (ABC Elite Kit; Vector Laboratories, Burlingame, California, USA). The sections were counter-stained with haematoxylin.
Double fluorescent immunostaining for Cdx1 and Cdx2 was performed. Deparaffinised sections were incubated with anti-Cdx1 (1:50) and anti-Cdx2 (1:50) antibodies. Binding of the primary antibodies was detected using fluorescein isothiocyanate (FITC)-conjugated anti-rabbit immunoglobulin or rhodamine-conjugated anti-mouse immunoglobulin (DAKO).
A polyclonal rabbit anti-Cdx1 antibody was raised in our laboratory by immunising a synthetic peptide corresponding to the carboxy terminus of mouse and rat Cdx1 (PSPVPVKEEFLP). This antibody is reactive to mouse, rat and human Cdx1 protein, and the specificity was confirmed by western blot analysis.
Cell lines and culture
Two cell lines, OE33 (a human oesophageal adenocarcinoma cell line; European Collection of Cell Cultures, Salisbury, UK) and Het-1A (a human normal oesophageal cell line immortalised by viral SV40 transfection; American Type Culture Collection, Manassas, Virginia, USA) were used in this study. For investigation of Cdx1 and Cdx2 promoter activation, OE33 cells were used to elucidate possible organ specific effects. For analysis of the direct effects of Cdx1 and Cdx2 on their expression, Het-1A cells were used, as possible transformation mechanisms of normal oesophageal keratinocytes were investigated.
Vector construction for reporter gene assays
For construction of a reporter vector of the Cdx1 promoter, accession no. M80463 was used. A length of 1429 bp of the Cdx1 promoter was amplified by polymerase chain reaction (PCR), then cloned into the SacI and BglII sites of a pGL3-basic luciferase vector (Promega, Madison, Wisconsin USA) to generate pCdx1/1429-Luc, and the cloned promoter sequences were confirmed by sequencing. Furthermore, 1014 bp of the Cdx2 promoter vector, pCdx2/1014-Luc, was constructed as described previously.14 As an internal control for the dual luciferase assay, pRL-TATA-Renilla-Luc, which expressed Renilla luciferase under a minimal TATA promoter, was also constructed.21
Expression plasmid
cDNA encoding full-length mouse Cdx1 was amplified by PCR using the following primers: 5′-CACCATGTACGTGGGCTATG-3′ and 5′-CTAGGGTAGAAACTCCTCCT-3′ (NCBI database NM-009880; National Center for Biotechnology Information, Department of Internal Medicine, Izumo City General Medical Center, Izumo, Japan). The amplified DNA samples were cloned into a pcDNA3.1/V5-His-TOPO vector (Invitrogen, Carlsbad, California, USA) and the plasmid clones were sequenced to confirm the constructs. Vector DNA without Cdx1 sequences was used as a negative control. A Cdx2 expression vector was also constructed as reported previously14 and vector DNA without Cdx2 sequences was used as a negative control.
Effects of bile acids on transcriptional activation of Cdx1
For evaluating the effects of bile acids on transcriptional activation of Cdx1, OE33 and Het-1A cells were cultured separately in 24-well plates (5×104 cells/well), then transfected with 0.5 μg of the promoter pCdx1/1429-Luc and 0.02 μg of pRL-TATA-Renilla-Luc in each well, using FuGENE 6 Transfection Reagent (Roche Diagnostics, Mannheim, Germany). A bile acids mixture (Sigma Chemical), which included cholic acid, glycocholic acid and taurocholic acid, was used as a stimulant. Twelve hours after transfection of the luciferase vectors, the cells were stimulated with various concentrations of the bile acids mixture or the vehicle (0.1% ethanol) alone for 12 h and cell lysates were used for determination of luciferase activity with a PicaGene Dual luciferase kit (Toyoinki, Tokyo, Japan), as described previously.21 The data are expressed as n-fold increases in luciferase activity of the bile acid-stimulated samples over that of the vehicle-stimulated samples.
Effects of bile acids on primary cultures of rat oesophageal keratinocytes
Primary cultures of oesophageal keratinocytes from normal rat oesophagi were established, as described previously,14 and used in experiments after the second passage. Following a 12 h incubation with the bile acids mixture, the cellular localisation of Cdx1 was investigated by Cdx1 immunofluorescence cytochemistry. After a 12 h incubation with various concentrations of the bile acids mixture, protein was extracted from cultured keratinocytes for measurement of Cdx1 expression by western blot analysis.
Homologous auto-regulations of Cdx1 and Cdx2
To investigate the homologous auto-regulation mechanism of Cdx1, the Cdx1 promoter was stimulated by transfection of a Cdx1 expression vector. OE33 cells were cultured in 24-well plates (5×104 cells/well) and transfected with 0.2 μg of the promoter pCdx1/1429-Luc, a total of 0.2 μg of various amounts of the Cdx1 expression vector or an empty vector, and 0.02 μg of pRL-TATA-Renilla-Luc in each well. To investigate the homologous auto-regulation mechanism of Cdx2, the Cdx2 promoter was stimulated by transfection of the Cdx2 expression vector. OE33 cells were cultured and transfected 0.2 μg of the promoter pCdx2/1014-Luc, a total of 0.2 μg of various amounts of the Cdx2 expression vector, a green fluorescent protein (GFP) expression vector (pEGFP-N1, unrelated protein control; BD Biosciences, San Jose, California, USA), or an empty vector, along with 0.02 μg of pRL-TATA-Renilla-Luc in each well.
Heterologous inter-regulation of Cdx1 and Cdx2
To investigate the heterologous inter-regulation mechanisms of Cdx1and Cdx2, the Cdx2 promoter was stimulated by the transfection of the Cdx1 expression vector. OE33 cells were cultured in 24-well plates (5×104 cells/well) and transfected with 0.2 μg of the promoter pCdx2/1014-Luc, a total 0.2 μg of the various amounts of the Cdx1 expression vector, a GFP expression vector, or an empty vector, along with 0.02 μg of pRL-TATA-Renilla-Luc in each well.
Similar to the method used to investigate the heterologous inter-regulation mechanisms of Cdx2 and Cdx1, the Cdx1 promoter was stimulated by transfection of the Cdx2 expression vector. OE33 cells were cultured and transfected with 0.2 μg of the promoter pCdx1/1429-Luc, a total 0.2 μg of various amounts of the Cdx2 expression vector, a GFP expression vector, or an empty vector, along with 0.02 μg of pRL-TATA-Renilla-Luc in each well.
To investigate the effects of the induced expression of Cdx1 or Cdx2 on Het-1A cells, the cells were transiently transfected with the Cdx1 or Cdx2 expression vector. The expressions of Cdx2 and Cdx1 in cultured Het-1A cells were investigated 48 h after transfection by immunofluorescence cytochemistry.
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) was performed using a Chromatin Immunoprecipitation Assay Kit (Upstate, Lake Placid, New York, USA) according to the manufacturer’s instructions. Briefly, Het-1A cells were incubated for 24 h after transfection with an empty or Cdx1 expression vector. The cells were cross-linked with 1% formaldehyde for 10 min then the nuclear fraction was isolated and sonicated to shear genomic chromatin. Chromatin was immunoprecipitated with anti-Cdx1 antibodies or without them, as a negative control, at 4°C overnight. Total DNA prior to immunoprecipitation was used as a positive control. Immunoprecipitated DNA–protein complexes were isolated and PCR was performed with the purified DNA using primers of the Cdx2 promoter region containing Cdx responsive elements (primers: 5′-TCCAGCCATTGGTGTCTGTG-3′, 5′-TTCCTTCTTTCCTCCCACCT-3′; product size, 151 bp; NCBI database Y13709). Twenty-five cycles of PCR were performed at 94°C for 30 s, 56°C for 30 s, and 72°C for 30 s.
Immunofluorescence cytochemistry
Primary oesophageal keratinocytes and Het-1A cells were grown separately on chambered glass slides, then fixed in 4% paraformaldehyde and permeabilised with 0.2% Triton X-100. The cells were labelled with anti-Cdx1 (1:50) and anti-Cdx2 (1:50). Binding of the primary antibodies was detected by FITC-conjugated anti-rabbit immunoglobulin, or rhodamine-conjugated anti-mouse immunoglobulin.
Statistical analysis
All data are expressed as the mean (with the SEM). Multiple comparisons were done with ANOVA followed by a Dunnett test. p Values of less than 0.05 were considered to be statistically significant.
RESULTS
Specificity of anti-Cdx1 antibody
To determine the specificity of the anti-Cdx1 antibody, Het-1A cells were transfected with a Cdx1 or Cdx2 expression vector. Results of western blot analysis showed that the anti-Cdx1 antibody reacted with the Cdx1 protein, whereas it did not cross-react with the Cdx2 protein (fig 1).

Specificity of the anti-Cdx1 antibody. Twenty-four hours after transfection of a Cdx1 or Cdx2 expression vector in Het-1A cells, protein was extracted and subjected to western blot analysis for Cdx1, Cdx2, and β-actin. The anti-Cdx1 antibody reacted with Cdx1 protein, but not with Cdx2 protein. Blots shown are representative of two separate experiments.
Immunohistochemistry examinations of rat oesophagus specimens
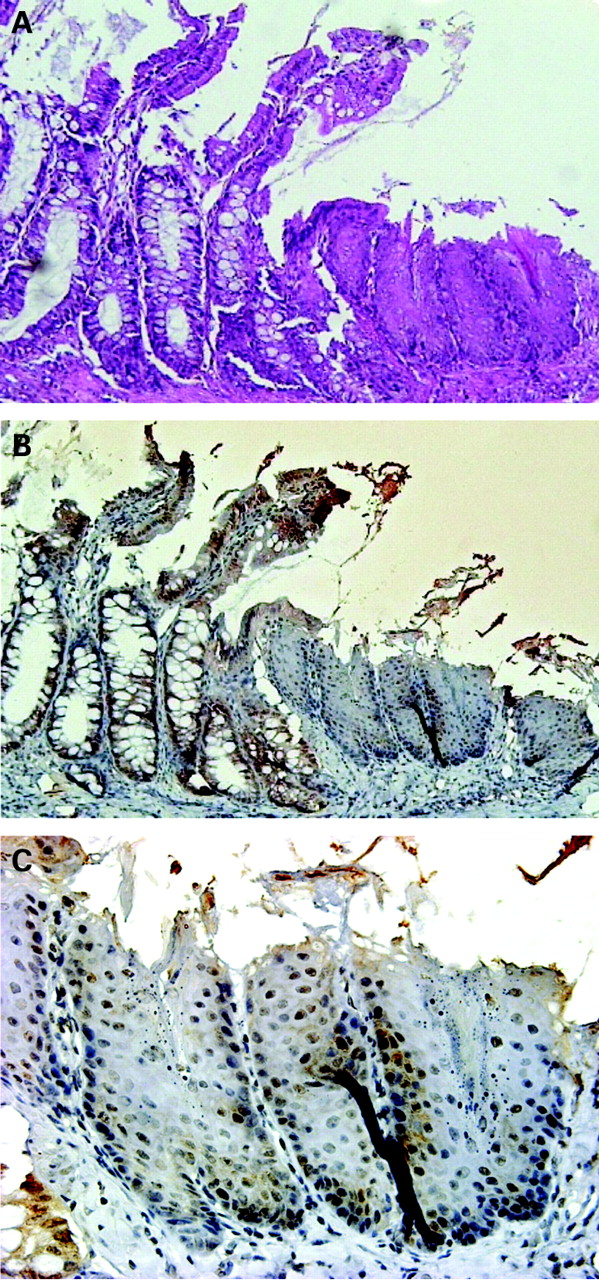
Two months after creation of the oesophageal–jejunal anastomoses, oesophagi in all rats were dilated and markedly thickened. The rats showed oesophagitis, which was histologically characterised by basal cell hyperplasia and elongation of papillae (fig 2A). Two months after the procedure, immunohistochemistry findings revealed Cdx1-positive cells with nuclear staining in squamous epithelia above the oesophageal-jejunostomy (fig 2B). No Cdx2-positive cells were observed in squamous epithelia above the oesophageal-jejunostomy (data not shown).
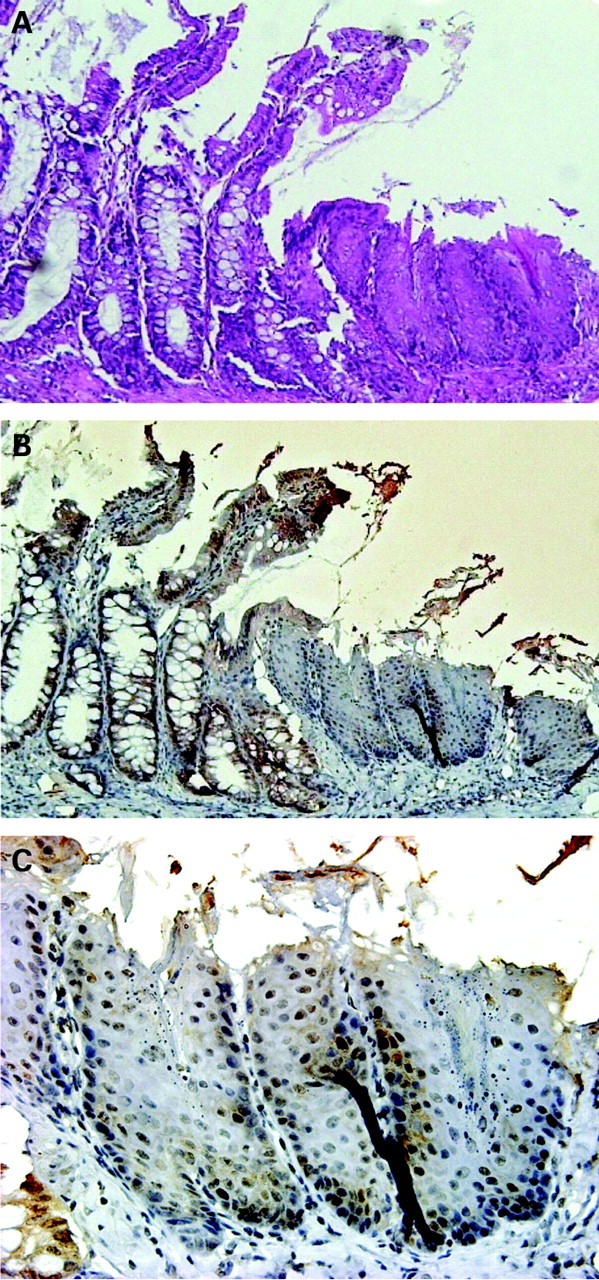
Oesophageal epithelium 2 months after formation of oesophageal–jejunal anastomosis. (A) Oesophageal–jejunal anastomosis (haematoxylin and eosin stain). (B) Results of immunohistochemistry for Cdx1 in oesophageal–jejunal anastomosis. Cdx1-positive cells were observed in the squamous epithelium. (C) Magnified view of immunohistochemistry results for Cdx1.
Six months after the procedure, columnar-lined epithelium consisting of absorptive cells and goblet cells was observed above the oesophageal-jejunostomy in 30% of the rats (fig 3A). Some islands of squamous epithelia remained in these epithelia, while areas of intermingled columnar and squamous epithelia with goblet cell formation were found both adjacent to and distant from the anastomosis. Basal cell hyperplasia and elongation of papillae with erosion and hyperkeratinisation were found in the most proximal epithelium (fig 4A). Cdx1-positive cells with nuclear staining were observed in the columnar epithelium above the oesophageal-jejunostomy as well as in squamous epithelia (fig 3B–D). Cdx1 positive cells were also diffusely expressed on the squamous epithelium with oesophagitis distant from the anastomosis (fig 4B).

Barrett’s epithelium 6 months after formation of oesophageal–jejunal anastomosis. (A) Metaplastic cells were formed adjacent to the site of anastomosis (haematoxylin and eosin stain). (B) Results of immunohistochemistry for Cdx1 in metaplastic epithelia and adjoining squamous epithelia. Cdx1-positive cells were observed in columnar and squamous epithelia. (C,D) Magnified views of immunohistochemistry results for Cdx1. Cdx1 positive cells were observed in metaplastic epithelia (C) and adjoining squamous epithelia (D).
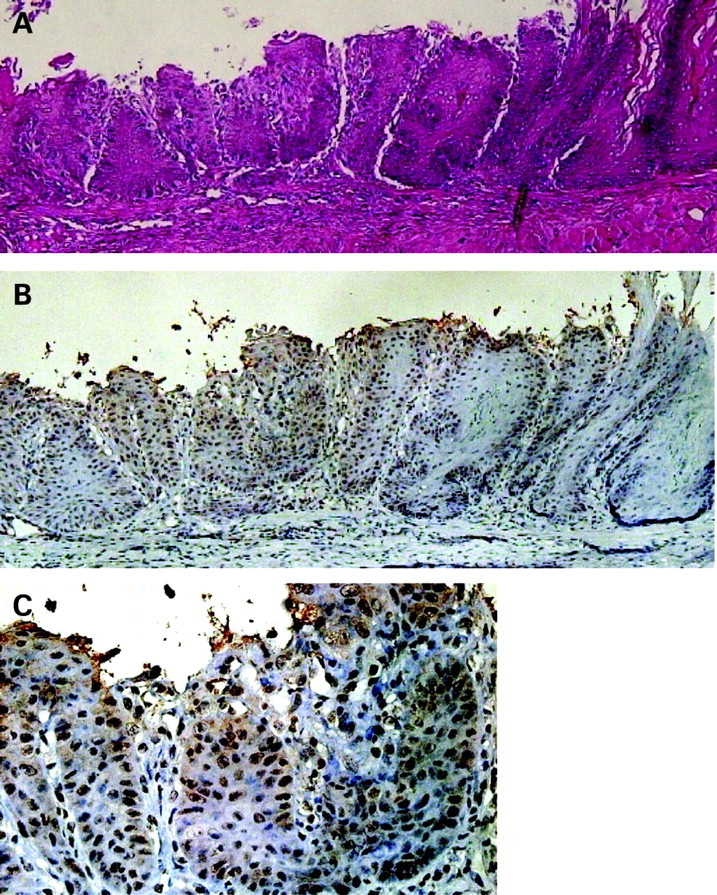
Squamous epithelia at 6 months after formation of oesophageal–jejunal anastomosis. (A) Squamous epithelia distant from the anastomosis (haematoxylin and eosin stain). (B) Results of immunohistochemistry for Cdx1. Cdx1 positive cells were observed in squamous epithelia distant from the anastomosis. (C) Magnified view of immunohistochemistry results for Cdx1.
To determine the co-localisation of Cdx1 and Cdx2 in Barrett’s epithelium, double-staining with Cdx1 and Cdx2 was performed. The pattern of expression of Cdx1 was similar to that of Cdx2 in the columnar cells of Barrett’s epithelium (fig 5).
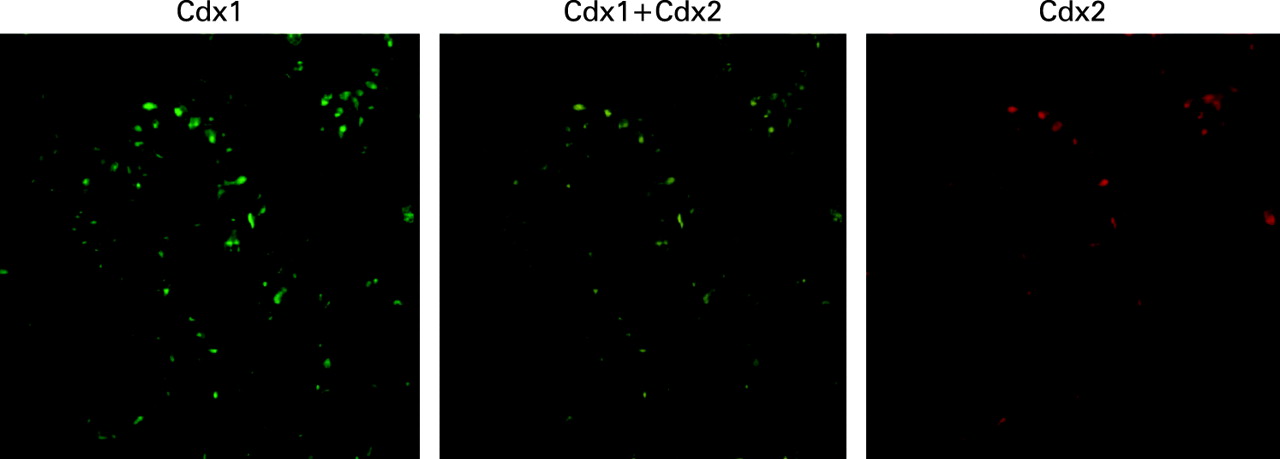
Barrett’s epithelium 6 months after formation of oesophageal–jejunal anastomosis. Shown are results of double staining of Cdx1 and Cdx2. The pattern of Cdx1 expression was similar to that of Cdx2 in columnar cells of Barrett’s epithelium.
Effects of bile acids on Cdx1 promoter activity
To investigate the possible effects of bile acids on Cdx1 gene expression, we evaluated Cdx1 promoter activity following stimulation with a bile acids mixture. To determine possible organ specific effects, the human oesophageal adenocarcinoma cell line OE33 and human normal oesophageal cell line Het-1A were used. The bile acids mixture had a strong stimulatory effect on Cdx1 promoter activity, with an approximately 1.5-fold increase in transcriptional activation in OE33 and Het-1A cells (fig 6A).

Effects of bile acids mixture on Cdx1 expression. (A) Effects of various concentrations of bile acids mixture on transcriptional activation of Cdx1 in OE33 and Het-1A cells. Cells were stimulated with various concentrations of the bile acids mixture or vehicle alone for 12 h, and cell lysates were used to determine luciferase activity. Data are expressed as the n-fold increase in luciferase activity in the bile acid-stimulated samples over that in the vehicle-treated samples. Results are expressed as the mean (with the SEM) of four experiments. *p<0.05 vs control. (B) Effects of bile acids mixture on Cdx1 protein expressions in primary cultured rat oesophageal keratinocytes. After incubation with 100 μmol/l of the bile acids mixture for 24 h, primary cultured cells were subjected to immunofluorescence cytochemistry for Cdx1. Cdx1 positive cells with nuclear staining were observed. (C) Effects of various concentrations of bile acids mixture on Cdx1 protein expression in primary cultured rat oesophageal keratinocytes. Cells were stimulated with various concentrations of the bile acids mixture or vehicle alone for 12 h, then protein was extracted and subjected to western blot analysis for Cdx1 and β-actin. Relative signal densities of Cdx1 to β-actin were quantified by densitometry. Blots shown are representative of three separate experiments. Results are expressed as the mean (with the SEM) of three experiments. **p<0.01 vs control.
Direct effects of bile acids mixture on Cdx1 expression in primary cultured oesophageal keratinocytes
To determine whether bile acids augment Cdx1 protein expression, we investigated the direct effect of the bile acids mixture on Cdx1 expression using primary cultured oesophageal keratinocytes and found that the bile acids augmented Cdx1 protein expression with nuclear staining (fig 6B). When examined by western blot analysis, Cdx1 protein expression was also augmented in a dose-dependent manner by the bile acids mixture (fig 6C).
Effects of over-expression of Cdx1 on oesophageal cells
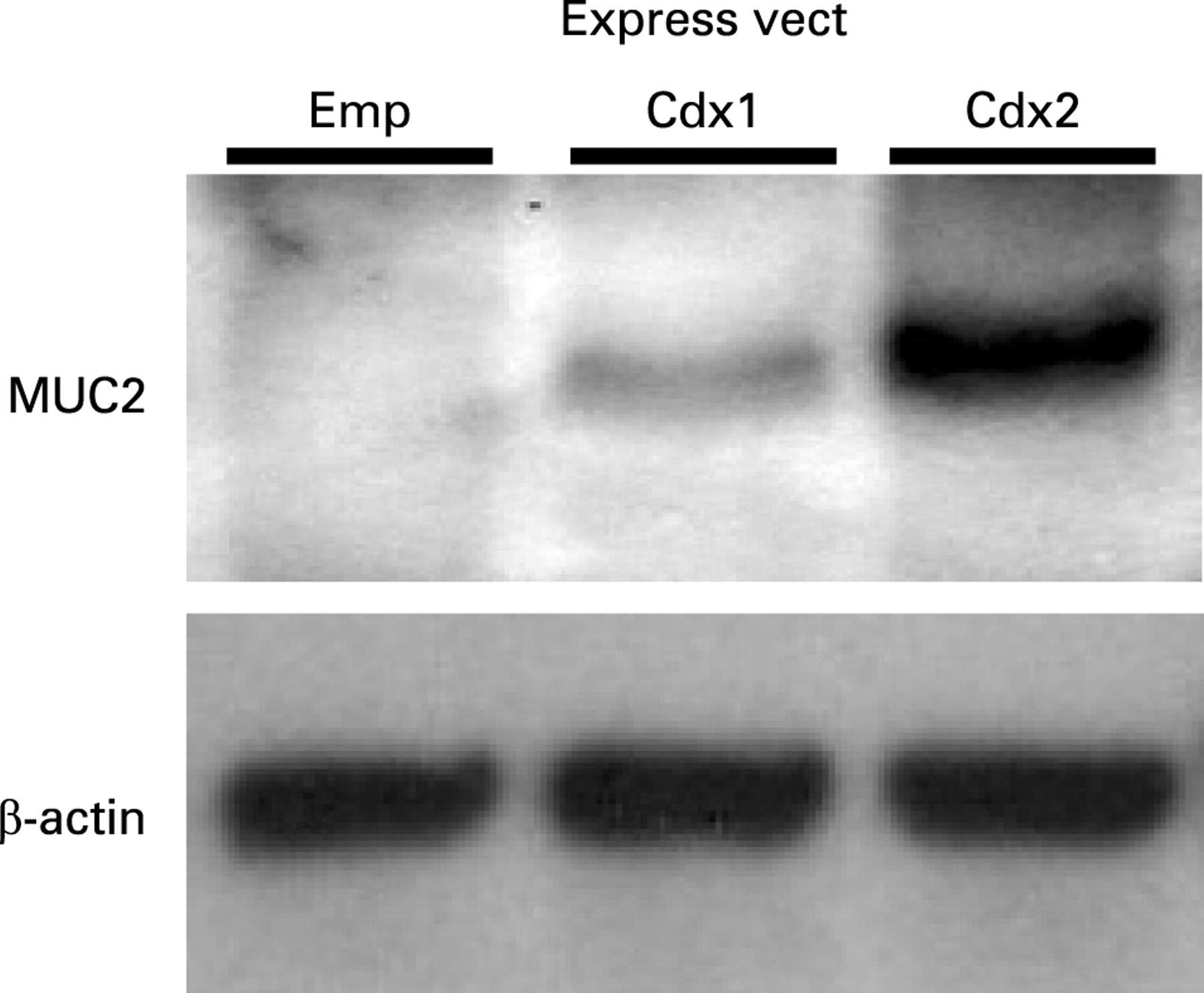
To investigate whether Cdx1 is the main regulator of intestinal metaplasia in oesophagi, we transfected a Cdx1 expression vector into the normal oesophageal cell line Het-1A and observed the expression of intestine specific mucin-2 (MUC2). The cells were transiently transfected with a Cdx1 expression construct and MUC2 expression was investigated. MUC2 was induced by transfection with a Cdx1 expression vector. When compared to the level of MUC2 expression induced by Cdx2, that induced by Cdx1 was significantly lower (fig 7). Therefore, we examined other indirect pathways that Cdx1 may use to regulate MUC2.

Mucin-2 (MUC2) protein expression after transfection of Cdx1 or Cdx2 expression vector in Het-1A cells. Forty-eight hours after transfection of an empty, Cdx1, or Cdx2 expression vector in Het-1A cells, protein was extracted and subjected to western blot analysis for MUC2 and β-actin. Blots shown are representative of three separate experiments. Emp, empty.
Homologous auto-regulations of Cdx1 and Cdx2
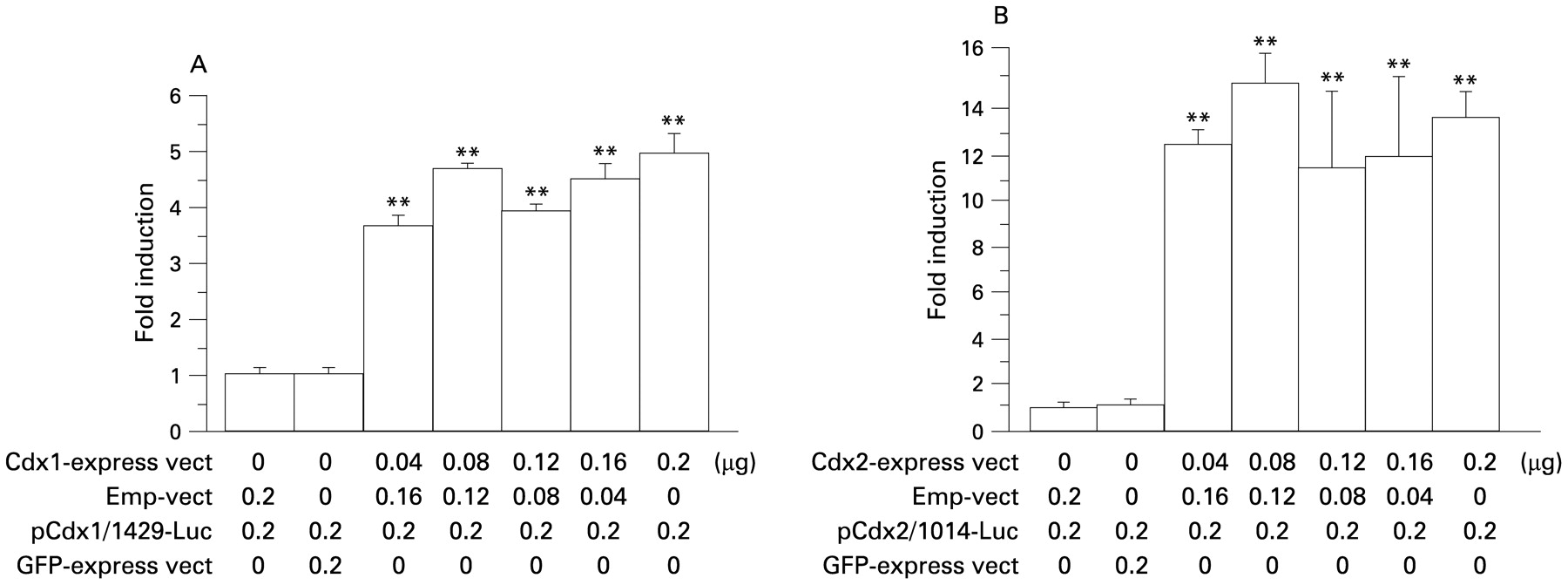
To investigate the homologous auto-regulation mechanism of Cdx1 on Cdx1 gene expression, Cdx1 promoter activity following stimulation with a Cdx1 expression vector was evaluated. Transfection of the Cdx1 expression vector into the cells increased Cdx1 promoter activity (fig 8A). This stimulatory effect by Cdx1 was specific, as transfection with a GFP expression vector did not have an effect on the promoter activity. To find the possible presence of a homologous auto-regulation mechanism in Cdx2 gene expression, Cdx2 promoter activity following stimulation with the Cdx2 expression vector was also evaluated. Transfection of the Cdx2 expression vector into the cells increased Cdx2 promoter activity (fig 8B).

Homologous auto-regulation mechanisms of Cdx1 and Cdx2. (A) Homologous auto-regulation mechanism of Cdx1. Cells were co-transfected with a Cdx1 promoter vector, Cdx1 expression vector, or empty vector (Emp-vect), then the cell lysates were used to determine luciferase activity 24 h after transfection. (B) Homologous auto-regulation mechanism of Cdx2. Cells were co-transfected with a Cdx2 promoter vector, Cdx2 expression vector, or empty vector, then the cell lysates were used to determine luciferase activity 24 h after transfection. Data are expressed as the n-fold increase in luciferase activity. Results are expressed as the mean (with the SEM) of four experiments. **p<0.01 vs control. GFP, green fluorescent protein.
Heterologous inter-regulation mechanism between Cdx1 and Cdx2
To investigate the heterologous inter-regulation mechanism of Cdx1 on Cdx2 gene expression, Cdx2 promoter activity following stimulation with a Cdx1 expression vector was evaluated. Transfection of the Cdx1 expression vector increased Cdx2 promoter activity (fig 9A). To confirm whether Cdx1 binds to the Cdx2 promoter, a ChIP assay was performed with Het-1A cells. A fragment with the expected size of 151 bp was detected in the immunoprecipitated DNA from cells transfected with the Cdx1 expression vector (fig 9B). On the contrary, no signal was detected in the immunoprecipitated DNA from cells transfected with an empty vector. Next, Cdx2 protein expression following stimulation with a Cdx1 expression vector was evaluated in Het-1A cells using immunofluorescence cytochemistry and cells transfected with the Cdx1 construct expressed the Cdx2 transcript (fig 9C).

Heterologous inter-regulation mechanism of Cdx2 stimulated by Cdx1. (A) OE33 cells were co-transfected with a Cdx2 promoter vector, Cdx1 expression vector, or empty vector (Emp-vect), then the cell lysates were used to determine luciferase activity 24 h after transfection. Data are expressed as the n-fold increase in luciferase activity. Results are expressed as the mean (with the SEM) of four experiments. **p<0.01 vs control. (B) Het-1A cells were transfected with an empty or Cdx1 expression vector, then lysed and subjected to a chromatin immunoprecipitation assay. Blots shown are representative of two separate experiments. (C) Effects of induced expression of Cdx1 in Het-1A cells. Cells were transiently transfected with a Cdx1 expression vector, then an immunofluorescence cytochemistry examination for Cdx1 and Cdx2 was conducted. Cells transfected with the Cdx1 expression vector expressed both Cdx1 and Cdx2 proteins. GFP, green fluorescent protein.
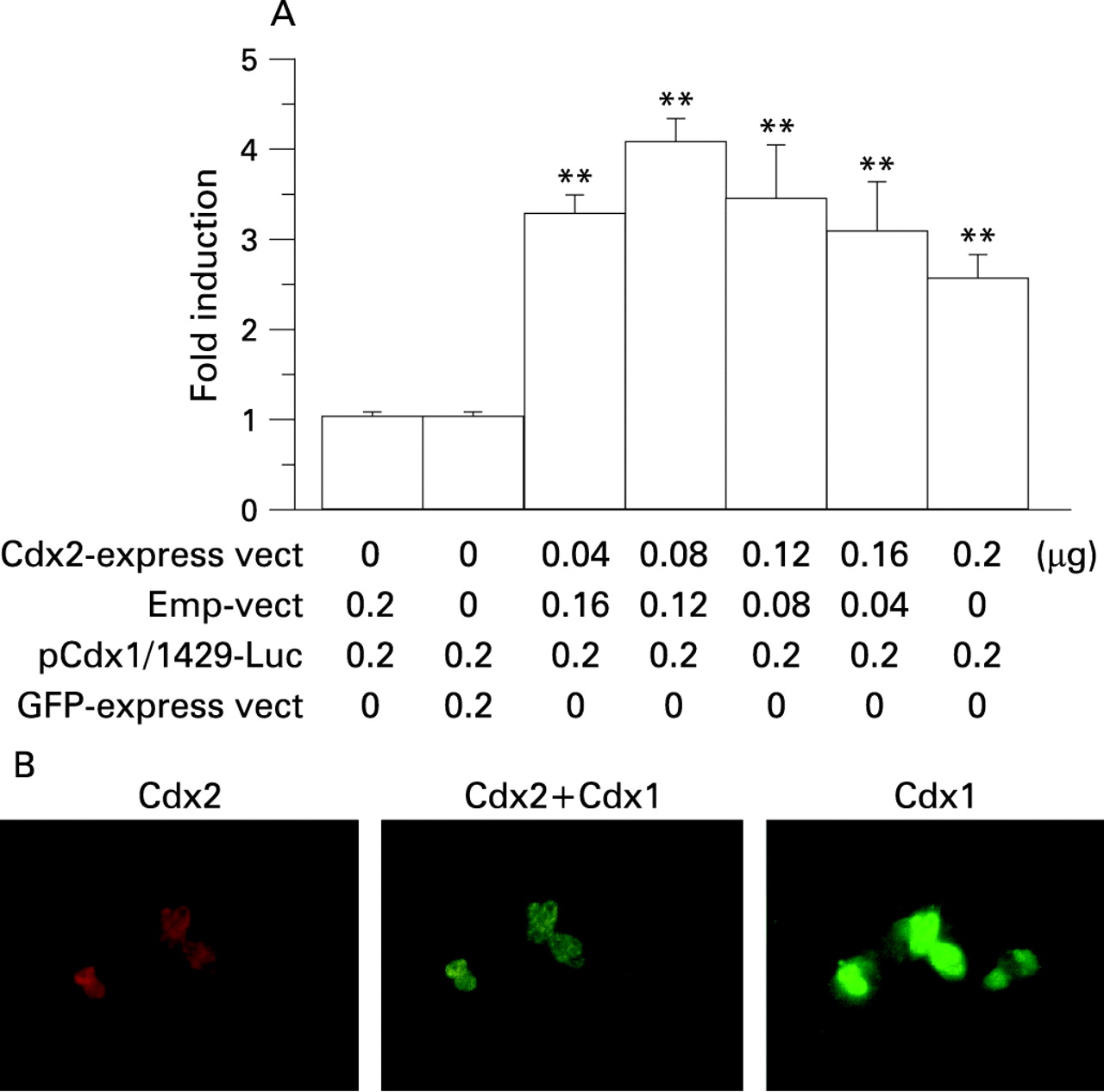
To investigate the heterologous inter-regulation mechanism of Cdx2 on Cdx1 gene expression, Cdx1 promoter activity following stimulation with a Cdx2 expression vector was evaluated and the results showed that transfection of the Cdx2 expression vector increased Cdx1 promoter activity (fig 10A). Finally, Cdx1 protein expression following stimulation with a Cdx2 expression vector was evaluated in Het-1A cells using immunofluorescence cytochemistry. Cells transfected with the Cdx2 construct expressed the Cdx1 transcript (fig 10B).

Heterologous inter-regulation mechanism of Cdx1 stimulated by Cdx2. (A) OE33 cells were co-transfected with a Cdx1 promoter vector, Cdx2 expression vector, or empty vector (Emp-vect), then the cell lysates were used to determine luciferase activity 24 h after transfection. Data are expressed as the n-fold increase in luciferase activity. Results are expressed as the mean (with the SEM) of four experiments. **p<0.01 vs control. (B) Effects of induced expression of Cdx2 in Het-1A cells. Cells were transiently transfected with a Cdx2 expression vector, then an immunofluorescence cytochemistry examination for Cdx2 and Cdx1 was conducted. Cells transfected with the Cdx2 expression vector expressed both Cdx2 and Cdx1 proteins. GFP, green fluorescent protein.
DISCUSSION
Our results suggest that Cdx1 is an important molecular mediator in the development of Barrett’s epithelium. Cdx2 is reported to be expressed in Barrett’s epithelium and a number of studies have found that it is also a key mediator in the development of Barrett’s oesophagus.12 13 22–24 Previously, we showed that the bile acid cholic acid activated the Cdx2 promoter via NF-κB and stimulated the production of Cdx2 protein in oesophageal keratinocytes,14 which is a significant mechanism in the development of Barrett’s epithelium. In addition to Cdx2, Cdx1 is also expressed in Barrett’s metaplasia.15 17 However, in contrast to Cdx2, the roles of Cdx1 in the development of Barrett’s oesophagus are not well understood. A previous study reported that over-expression of Cdx1 induced intestinal metaplasia in transgenic mouse stomachs and they suggested that Cdx1 has a major role in the development of intestinal metaplasia.16 Furthermore, Wong et al15 suggested that Cdx1 is an important molecular mediator of Barrett’s metaplasia and that bile acids stimulate the Cdx1 promoter via NF-κB. Therefore, it is considered that Cdx1 and Cdx2 have similar roles in the development of Barrett’s oesophagus.
First, we examined the effects of bile acids on the expression of Cdx1 in vivo using Barrett’s epithelium formed in rats with an oesophago-jejunal anastomosis. This rat model created with oesophageal-jejunostomy develops severe oesophagitis, Barrett’s epithelium, and finally oesophageal adenocarcinoma.19 20 25–28 Barrett’s epithelium in rats resembles the lesions described in human Barrett’s oesophagus in regard to morphology and the expression pattern of cancer-related gene products, including p53, c-myc, and cyclooxygenase 2.28–30 An area of intermingled columnar and squamous epithelia is called multilayered epithelium, and is a presumed precursor of Barrett’s epithelium. Results of other studies with the present rat model suggest that the lesions in Barrett’s epithelium develop via a gut regenerative cell lineage.29 31 32
In the present study, we investigated Cdx1 expression in a rat model and compared it to that of Cdx2. Immunohistochemistry findings revealed that the distribution of Cdx1 positive cells was different from that of Cdx2. In our rat models, no metaplastic epithelia were observed at 2 months after surgery, while Cdx1 positive cells with nuclear staining were observed in the squamous epithelia above the anastomois site (fig 2B) and no Cdx2 positive cells were found. At 6 months after the procedure, metaplastic epithelia were observed, with Cdx2 positive cells observed in the metaplastic epithelia and minimally in the adjoining squamous epithelia area.14 In addition, Cdx1 positive cells with nuclear staining were observed not only in the metaplastic epithelia, but also in the wider area of squamous epithelia (fig 3C–E). Furthermore, Cdx1 positive cells in the squamous epithelia had spread diffusely to distant areas from the site of anastomosis (fig 4B). These findings suggest that bile acids stimulate Cdx1 expression in oesophageal squamous epithelium and Cdx1 plays an important role in the development of Barrett’s epithelium. When double staining of Cdx1 and Cdx2 was performed, the pattern of expression of Cdx1 was similar to that of Cdx2 in columnar cells of Barrett’s epithelium. Therefore, our results indicate that Cdx1 expression occurs prior to Cdx2 expression during development of the condition.
Next, we investigated the roles of bile acids on Cdx1 expression in vitro. In patients with Barrett’s oesophagus, the most common bile acids found in the refluxant are cholic acid, glycocholic acid and taurocholic acid.6 Since a mixture of bile acids is considered to provide the physiological stimulation, we investigated the changes in Cdx1 promoter activity following stimulation with such a mixture and found that the bile acids stimulated Cdx1 promoter activity in a dose-dependent manner (fig 6A). We also investigated whether bile acids directly stimulate Cdx1 expression in undifferentiated immature oesophageal keratinocytes. In a previous study, we established a primary culture of rat oesophageal keratinocytes and found that the major population of cultured cells were undifferentiated immature oesophageal keratinocytes.14 When the cells were stimulated by the bile acids mixture in the present study, Cdx1 positive cells with nuclear staining were observed (fig 6B). Further, we investigated Cdx1 protein expression following stimulation with the bile acids mixture by western blot analysis and found that the bile acids stimulated Cdx1 protein expression in a dose-dependent manner (fig 6C).
Cdx2 is reported to be a direct transcriptional activator of a number of intestine specific genes, including MUC2, sucrase–isomaltase, lactase–phlorizin hydrolase, glucagon, and guanylyl cyclase C.33–36 We reported that over-expression of Cdx2 induced MUC2 expression in oesophageal keratinocytes,14 while Cdx1 is also known to be a direct transcriptional activator of some intestine specific genes, including MUC2 and intestinal alkaline phosphatase (IAP).37–39 In the present study, we examined whether the induction of Cdx1 expression in oesophageal epithelial cells would trigger a similar transdifferentiation to intestinal type columnar epithelial cells. Forced expression of Cdx1 in the normal oesophageal cell line Het-1A slightly induced goblet-like cells expressing MUC2 (fig 7). Cdx1 and Cdx2 have also been reported to differentially regulate intestine specific genes, including MUC2 and IAP,37–39 and MUC2 regulation by Cdx1 has been suggested to be cell type specific. Also, Yamamoto et al37 reported that Cdx2 stimulated MUC2 expression in a kidney cell line, COS-7, whereas Cdx1 did not, and Mesquita et al38 noted that Cdx1 induced MUC2 expression in various colon cancer cell lines, whereas it did not induce that expression in gastric cancer cell lines. Therefore, the mechanism of development of Barrett’s epithelium due to over-expression of Cdx1 by bile acids is suggested to be indirect.
A number of homeobox genes, including Cdx1 and Cdx2, have been shown to positively regulate their own expression.40–42 Multiple complex mechanisms have been suggested to function in the regulation of Cdx1 and Cdx2, since the Cdx promoter has multiple Cdx responsive elements, with a sequence of TTTAT(A/G) or TTTA(T/C).10 43 44 Therefore, we investigated possible homologous auto-regulation mechanisms of Cdx1 and Cdx2. The effects of bile acids on Cdx1 promoter activity in OE33 and Het-1A cells were nearly identical, whereas the transfection efficiency of OE33 cells was shown to be much higher than that of Het-1A cells. In addition, in the experiments with OE33 cells in which co-transfection of exact doses of the expression, promoter and control vectors was required, the results were more reproducible. Transfection of the Cdx1 expression vector into OE33 cells increased Cdx1 promoter activity, while that of the Cdx2 expression vector increased Cdx2 promoter activity (fig 8A,B). Activation of their own promoters by the Cdx1 and Cdx2 expression vectors was beyond the level of physiological activation by bile acids, as shown in fig 6, and reached peak levels at the lowest concentrations (fig 8A,B). Therefore, even at lower concentrations, Cdx1 or Cdx2 may strongly induce homologous activation of their own promoters. As a next step, we investigated the heterologous inter-regulation mechanisms between Cdx1 and Cdx2. Transfection of the Cdx1 expression vector induced Cdx2 promoter activity in a dose-dependent manner (fig 9A), while ChIp results revealed that Cdx1 protein binds to Cdx responsive elements of the Cdx2 promoter (fig 9B). Furthermore, transfection of the Cdx1 expression vector into the normal oesophageal cell line Het-1A induced production of Cdx2 protein (fig 9C). On the contrary, transfection of the Cdx2 expression vector into OE33 cells induced Cdx1 promoter activity (fig 10A) and that into Het-1A cells induced production of Cdx1 protein (fig 10B). Activation of the Cdx1 promoter by transfection of the Cdx2 expression vector was dose dependent at lower dosages (0.04–0.08 μg), whereas it slightly decreased with higher dosages (0.12–0.20 μg). Thus, a higher level of activation of Cdx2 may induce a negative feedback effect on Cdx1 expression. These findings suggest that auto- and inter-regulation between Cdx1 and Cdx2 contribute to cellular proliferation and transdifferentiation in intestinal metaplasia following stimulation by bile acids. Based on our findings that Cdx1 expression preceded that of Cdx2 in squamous epithelium in our Barrett’s rat model, it is suggested that over-expression of Cdx1 by bile acids induces MUC2 expression via Cdx2.
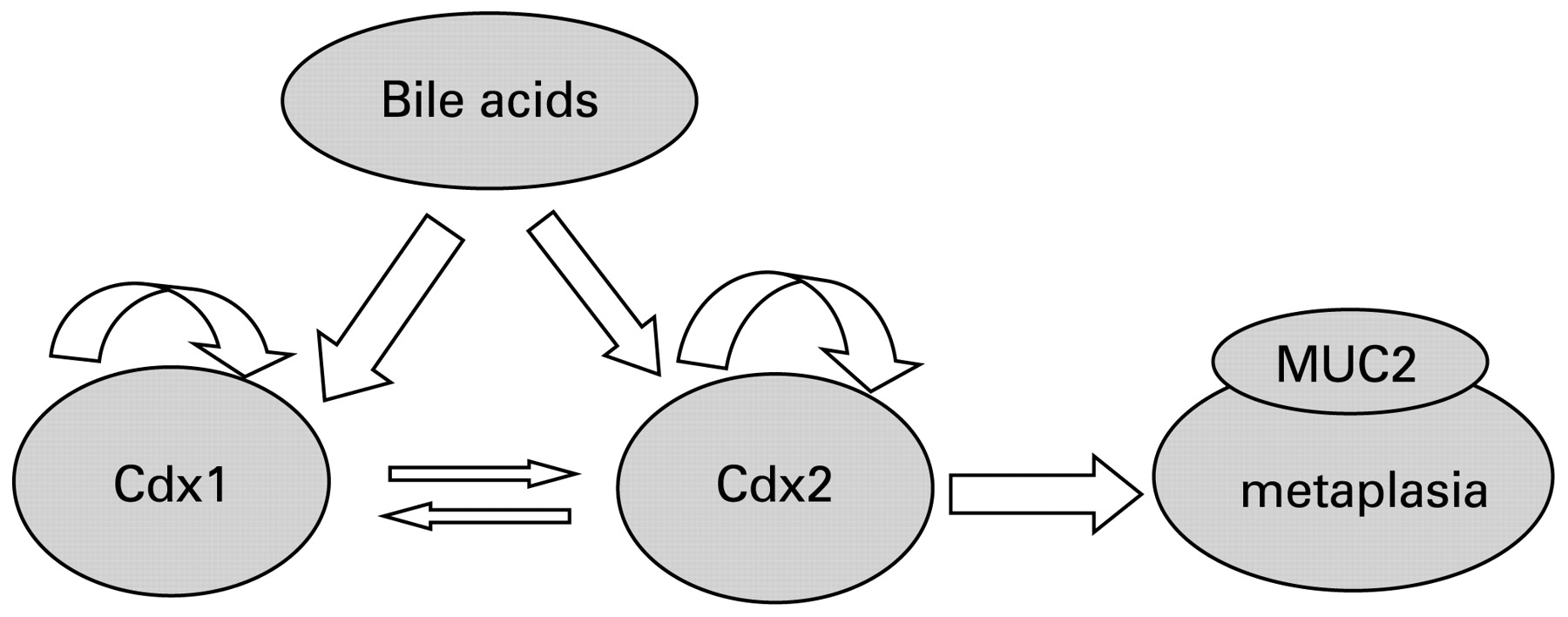
In conclusion, two mechanisms are suggested in the development of Barrett’s epithelium (fig 11). Bile acids induce the expression of both Cdx1 and Cdx2 in oesophageal keratinocytes. In addition, over-expression of Cdx2 induces MUC2 directly, whereas that of Cdx1 indirectly induces MUC2 via Cdx2 expression.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Schema of mechanism of Barrett’s epithelium development. Bile acids directly stimulate the expressions of Cdx1 and Cdx2. Next, over-expression of Cdx2 directly induces mucin-2 (MUC2), while that of Cdx1 indirectly induces MUC2 via Cdx2 expression.
REFERENCES
Footnotes
Competing interests: None.
Ethics approval: The experimental study protocol was approved by the institutional animal care and experimental committee of Shimane University on 1 February 2006.
Linked Articles
- Digest