Article Text

Abstract
Objective Fibroblast growth factor 15/19 (FGF15/19), an enterokine that regulates synthesis of hepatic bile acids (BA), has been proposed to influence fat metabolism. Without FGF15/19, mouse liver regeneration after partial hepatectomy (PH) is severely impaired. We studied the role of FGF15/19 in response to a high fat diet (HFD) and its regulation by saturated fatty acids. We developed a fusion molecule encompassing FGF19 and apolipoprotein A-I, termed Fibapo, and evaluated its pharmacological properties in fatty liver regeneration.
Design Fgf15−/− mice were fed a HFD. Liver fat and the expression of fat metabolism and endoplasmic reticulum (ER) stress-related genes were measured. Influence of palmitic acid (PA) on FGF15/19 expression was determined in mice and in human liver cell lines. In vivo half-life and biological activity of Fibapo and FGF19 were compared. Hepatoprotective and proregenerative activities of Fibapo were evaluated in obese db/db mice undergoing PH.
Results Hepatosteatosis and ER stress were exacerbated in HFD-fed Fgf15−/− mice. Hepatic expression of Pparγ2 was elevated in Fgf15−/− mice, being reversed by FGF19 treatment. PA induced FGF15/19 expression in mouse ileum and human liver cells, and FGF19 protected from PA-mediated ER stress and cytotoxicity. Fibapo reduced liver BA and lipid accumulation, inhibited ER stress and showed enhanced half-life. Fibapo provided increased db/db mice survival and improved regeneration upon PH.
Conclusions FGF15/19 is essential for hepatic metabolic adaptation to dietary fat being a physiological regulator of Pparγ2 expression. Perioperative administration of Fibapo improves fatty liver regeneration.
- CHOLESTASIS
- LIVER REGENERATION
- GROWTH FACTORS
- LIVER FAILURE
- FATTY LIVER
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
FGF15/19 is a bile acid (BA)-induced ileum-derived enterokine that governs BA homeostasis. Lack of FGF15 results in impaired mouse liver regeneration after partial hepatectomy mainly due to toxic intrahepatic BA concentrations.
FGF15/19 regulates hepatic glucose metabolism and stimulates protein synthesis. Pharmacological administration or transgenic expression of FGF19 in mice increases hepatic lipid oxidation, reduces lipogenesis and protects from hepatosteatosis.
Hepatic steatosis, and the cholestasis associated with this condition, negatively affects experimental and human liver regeneration after hepatectomy or after living donor liver transplantation.
What are the new findings?
Endogenous FGF15/19 plays a central role in hepatic lipid metabolism. Lack of FGF15 results in increased hepatic steatosis and in the development of endoplasmic reticulum (ER) stress in the liver of mice fed a high fat diet.
FGF15/19 is a physiological negative regulator of the hepatic expression of the transcription factor Pparγ2, which is a major contributor to high-fat diet-induced hepatic steatosis.
FGF19 gene expression can be activated in parenchymal and biliary human liver cells by ER stress-inducing saturated fatty acids, and FGF19 protects these cells from lipoapoptosis.
We have developed a new chimaeric molecule encompassing FGF19 and apolipoprotein A-I termed Fibapo with improved pharmacokinetic properties. Fibapo retains FGF19 biological activities on liver BA and fat metabolism and displays potent hepatoprotective and proregenerative effects.
How might it impact on clinical practice in the foreseeable future?
The FGF15/19-PPARγ2 axis identified in this study may constitute a novel target for the prevention or treatment of hepatic steatosis.
Engineered FGF19-based molecules such as Fibapo may be applied perioperatively for the improvement of liver regeneration after resection or transplantation, particularly in the presence of hepatosteatosis and cholestasis.
Introduction
Non-alcoholic fatty liver disease (NAFLD), a spectrum of liver conditions ranging from simple steatosis to steatohepatitis, and eventually progressing to fibrosis and cirrhosis, affects up to 30% of the general population and 70%–90% of obese individuals.1 The accumulation of hepatic triglycerides (TG) in the cytoplasm of hepatocytes is the histological hallmark of NAFLD. However, TG accumulation per se is not thought to mediate hepatocellular damage, being considered as an adaptive protective response to cope with the increased flux of free fatty acids (FFA) within the liver.1 In patients with NAFLD, accumulating hepatic FFAs come from different sources, including the adipose tissue where insulin resistance promotes lipolysis, from de novo lipogenesis and from the diet.1 ,2 Peripheral insulin resistance leads to a compensatory hyperinsulinemia, which further drives hepatic de novo lipogenesis. FFAs’ excess, and particularly saturated FFAs’ accumulation, leads to hepatocellular insulin resistance and lipotoxicity through various mechanisms among which an excessive or prolonged endoplasmic reticulum (ER) stress response is believed to play a key role.1–3 Further to ER stress, FFAs may also promote increased synthesis and accumulation of bile acids (BA) leading to hepatocyte injury.4 Consistently, elevated hepatic BA levels have been found in patients with NAFLD.5 Moreover, FFAs can be also toxic for cholangiocytes, contributing to the development of cholestasis also observed in a subset of patients with NAFLD.6
Insulin is indeed essential for the coordination of lipid and carbohydrate metabolism, and hepatic steatosis appears when insulin signalling is impaired in adipose tissue and the liver.1 However, recent studies have identified fibroblast growth factor 19 (FGF19; FGF15 in rodents) as an additional nutritionally regulated hormone capable of inhibiting gluconeogenesis and stimulating hepatic glycogen and protein synthesis like insulin, but devoid of the lipogenic effects of this hormone.7 ,8 FGF15/19 was initially identified as a postprandial endocrine factor released into the portal circulation from ileal enterocytes involved in the inhibition of hepatic BA synthesis.9 In the context of diet-induced NAFLD, transgenic expression of FGF19 or its pharmacological administration to obese mice resulted in increased insulin sensitivity and reduced hepatosteatosis.10 ,11 Interestingly, a number of reports have found decreased fasting FGF19 serum levels in patients with NAFLD, type 2 diabetes or the metabolic syndrome, as well as an apparent resistance to FGF19 hepatic effects in insulin-resistant patients with NAFLD.12 Together, these findings suggest a potential role for FGF15/19 in hepatic lipid homeostasis and highlight its dysregulation in metabolic disorders. However, to our knowledge, the physiological function of this hormone in the response to an obesogenic diet has not been examined. In the first part of this study, we demonstrate the critical role of endogenous FGF15 in the response to diet-induced hepatic steatosis.
While the impact of NAFLD on human health is generally associated with chronic liver disease progression, there are specific circumstances in which the deleterious consequences of NAFLD can manifest acutely. NAFLD negatively affects the outcome of patients undergoing partial liver resection of primary or metastatic liver tumours, or in the setting of liver transplantation.13–15 Importantly, the presence of cholestasis, existing prior to resection or developing after liver transplantation, has been associated with steatosis and a worse patient’s outcome.13 ,16 Experimental studies in the models of NAFLD have also demonstrated the negative impact of this condition on liver regeneration after partial hepatectomy (PH).17–20 Interestingly, we and others have recently shown that FGF15/19 is important for mouse liver regeneration after PH, contributing to the preservation of BA homeostasis and to parenchymal cells’ survival and proliferation.21–23 Moreover, we demonstrated that FGF15/19 delivery increased mice survival in a model of extensive hepatectomy, suggesting the therapeutic potential of the perioperative administration of this enterokine.21 Therefore, the combined intrinsic biological activities of FGF15/19, namely its ability to lower liver fat, regulate BA levels and promote hepatocellular proliferation, would make this molecule an ideal tool to improve regeneration of steatotic and cholestatic livers. However, FGF15/19 protein has a very short half-life, with a high glomerular filtration rate.24 To overcome this limitation, we have developed a new chimaeric molecule based on the fusion of FGF19 with apolipoprotein A-I (ApoA-I) termed Fibapo (FA). As we have previously shown, the ApoA-I moiety confers biological stability and provides liver targeting through its interaction with scavenger receptor class B type I (SR-BI) highly expressed in hepatocytes.25 In the second part of this study, we demonstrate the remarkable metabolic and proregenerative activities of FA in a model of fatty liver.
Methods
Animals and treatments
Fgf15−/− mice and wild-type littermate controls (Fgf15+/+ ) have been described before.21 ,26 Obese db/db mice (C57BL/6 background, male, 8–10 weeks of age) were from Charles River (Barcelona, Spain). Fgf15−/− and Fgf15+/+ mice were fed a high-fat diet (HFD) (60% of kcal from fat, Research Diets, New Brunswick, New Jersey, USA) for 12 weeks, starting at 8 weeks of age. Body fat content was measured in live conscious animals using quantum molecular resonance (QMR) technology (EchoMRI-100-700, Echo Medical Systems, Houston, Texas, USA). Where indicated, after 14 hours of fasting, mice were subjected to a 500 μL bolus p.o of a nutrient-rich diet (Ensure, Abbot Laboratories, 4.6 mg/g body weight) or of palmitic acid (PA) resuspended in palm oil (2 mg/g body weight) (Sigma). Wild-type C57BL/6 mice (8–10 weeks of age) were fasted for 14 hours and then treated with tunicamycin (100 μg/mouse) in a 150 mM sucrose solution p.o. Hydrodynamic injections of plasmids were performed as described.25 PHs (66% and 85%) were performed in mice as described.21 Treatment of mice with recombinant FGF19 or FA for the indicated periods of time did not significantly affect weight or food intake. Mice received humane care, and all experiments were carried out in compliance with our institution’s ethical guidelines.
Additional Materials and Methods are included as online supplementary files and supplementary tables S1 and S2.
supplementary file
supplementary tables
Results
Lack of FGF15/19 leads to increased adiposity and hepatic steatosis accompanied by ER stress in response to a HFD
To directly assess the potential influence of FGF15/19 to an obesogenic diet, Fgf15+/+ and Fgf15−/− mice were fed a control or a HFD for 12 weeks. Whereas Fgf15−/− mice fed a chow diet gained less weight than Fgf15+/+ , this outcome was reversed on a HFD, without significant differences in food intake between genotypes (figure 1A). Total body fat and intrahepatic TG content were also markedly higher in Fgf15−/− mice (figure 1B, C). Hepatic TG levels were also elevated in Fgf15−/− mice compared with Fgf15+/+ , even under a normal diet (figure 1C). While de novo liver fatty acid synthesis from carbohydrate sources is mainly regulated by the transcription factors sterol-regulatory element-binding protein 1c (SREBP1c) and carbohydrate-responsive element-binding protein (ChREBP),1 lipid accumulation in diet-induced hepatic steatosis appears to be mainly mediated by peroxisome proliferator-activated receptor γ variant 2 (PPARγ2).27 Pparγ2 levels are low in the normal liver becoming induced upon HFD feeding.27 Accordingly, Pparγ2 expression was upregulated in Fgf15+/+ mice fed a HFD, and the expression of this gene reached highest levels in FGF15-null mice (figure 1D). Moreover, hepatic Pparγ2 expression was already elevated in Fgf15−/− mice fed a control diet (figure 1D). However, we did not observe any significant differences in the hepatic expression of the lipogenic genes Srebp1 and Fas (fatty acid synthase) between Fgf15−/− and Fgf15+/+ mice under any feeding conditions (data not shown).

Lack of FGF15/19 leads to increased adiposity and hepatic steatosis accompanied by endoplasmic reticulum (ER) stress in response to a high fat diet (HFD). Weight gain (A), body fat content (B) and liver triglycerides, TG (C) in Fgf15+/+ (wild-type, WT) and Fgf15−/− mice knockout (KO) after 12 weeks on a chow diet or a HFD. The expression levels of Pparγ2 mRNA (AU, arbitrary units) and PPARγ2 protein (D), and the mRNA levels of the PPARγ2 target genes Cd36 and Mogat1 (E) in the liver of Fgf15+/+ (WT) and Fgf15−/− mice (KO), fed a chow or a HFD for 12 weeks, were determined by qPCR or western blotting. Expression of Pparγ2, Cd36 and Mogat1 in the liver of Fgf15−/− mice (KO) at different time points after recombinant FGF19 administration (1 mg/kg, subcutaneously) (F). Circulating FGF19 levels and hepatic PPARγ2 mRNA expression in samples from healthy (n=18) and obese (n=69) patients as determined by ELISA and qPCR, respectively (G). Expression of genes involved in the ER stress response in the liver of Fgf15+/+ (WT) and Fgf15−/− mice (KO) fed a chow or a HFD for 12 weeks examined at the mRNA (H) and protein levels (I). Representative blots are shown, and quantitation of band intensities (normalised to β-ACTIN) is indicated. N=5 mice per group. *p<0.05; **p<0.01; ***p<0.001; ap<0.05 vs WT; bp<0.05 vs 0 hours; cp<0.001 vs 0 hours; dp<0.0001 vs 0 hours.
The biological significance of Pparγ2 overexpression was supported by the concomitant upregulation of two key PPARγ2 target genes, Cd36/FAT (fatty acid translocase) and monoacylglycerol O-acyltransferase 1 (Mogat1), involved in liver fat accumulation (figure 1E and online supplementary figure S1A).27 These findings suggested that FGF15/19 might directly regulate the expression of hepatic Pparγ2. To evaluate this possibility, we treated Fgf15−/− mice with recombinant FGF19 and measured the hepatic expression of Pparγ2, Cd36 and Mogat1 at different time points. As shown in figure 1F, FGF19 markedly reduced the expression of these genes. Consistently, Pparγ2 mRNA levels were also reduced in Fgf15−/− and wild-type mice upon infection with an FGF15-expressing adenovirus (see online supplementary figure S1B). This in vivo effect of FGF19 on Pparγ2 expression is likely to be mediated through a direct interaction with the hepatocyte, as we observed that treatment of cultured human hepatic cells (HepG2) with FGF19 significantly reduced Pparγ2 mRNA levels (see online supplementary figure S1C). Some of us recently reported that FGF19 circulating levels are low in obese patients.28 Data in figure 1G confirm this and interestingly show that hepatic PPARγ2 expression was markedly induced in these individuals.
supplementary figures
ER stress has been observed both in liver samples from patients with NAFLD and experimental models of fatty liver.1 ,2 ,29 Importantly, chronic ER stress is linked to lipid metabolism dysregulation and fatty liver disease progression.1 ,2 ,30 In view of this, we examined the expression of key ER stress response genes in the livers of Fgf15+/+ and Fgf15−/− mice fed control or HFD for 12 weeks. We found that while Fgf15+/+ mice still did not show signs of ER stress, FGF15-null mice on HFD displayed higher mRNA levels of Grp78, Atf6, Perk and C/EBP homologous proteins (CHOP), indicative of an ongoing ER stress response (figure 1H).19 ,30 ,31 Consistent with the previous findings in db/db obese mice,31 GRP78 protein levels were reduced in the liver of Fgf15−/− mice, particularly upon HFD feeding, while CHOP levels were induced (figure 1I).
FGF15/19 was recently identified as an ER stress response gene in human colonic cells and mouse ileum.32 In that study, FGF15/19 expression was triggered with chemical ER stress inducers in a farnesoid X receptor independent manner.32 However, the physiological significance of this response remains unknown. We reproduced the activation of ileal Fgf15 expression, along with that of ER stress response genes, in mice gavaged with the chemical ER stress inducer tunicamycin (figure 2A). In view of this, we speculated that a HFD, particularly a diet rich in saturated FAs which are strong ER stress inducers,2 could trigger ileal FGF15/19 expression. We found that mice gavaged with a bolus of palm oil enriched with PA, or with a standard nutrient-rich liquid diet (Ensure), showed the expected peak of ileal Fgf15 expression previously observed between 60 and 90 min post gavage (figure 2B).33 However, in PA-gavaged mice, ileal Fgf15 expression remained higher than in control mice at later time points, with a second peak between 6 and 8 hours, coinciding with the sustained activation of ER stress response genes (figure 2B). Concomitantly, increased hepatic expression of ER stress marker genes was observed in PA-gavaged mice compared with the mice fed the standard diet (figure 2C).

FGF15 expression is induced by endoplasmic reticulum (ER) stress in mouse ileum. Wild-type mice received tunicamycin (Tm) (100 μg/mouse) in a 150 mM sucrose solution, or the same volume of this vehicle (Veh) by gavage, and were sacrificed 6 hours later. The expression of the indicated genes involved in ER stress, and that of Fgf15, was measured by qPCR in ileal tissues (A). Fasted wild-type mice were gavaged with a bolus of control liquid diet or with the same volume of a palmitic acid (PA)-enriched palm oil suspension. At the indicated time points, the expression of ER stress-related genes was determined by qPCR in the ileum (B) or liver (C) of these mice. N=5 mice per group. *p<0.05; **p<0.01; ***p<0.001. AU, arbitrary units
FGF19 protects liver cells from lipoapoptosis
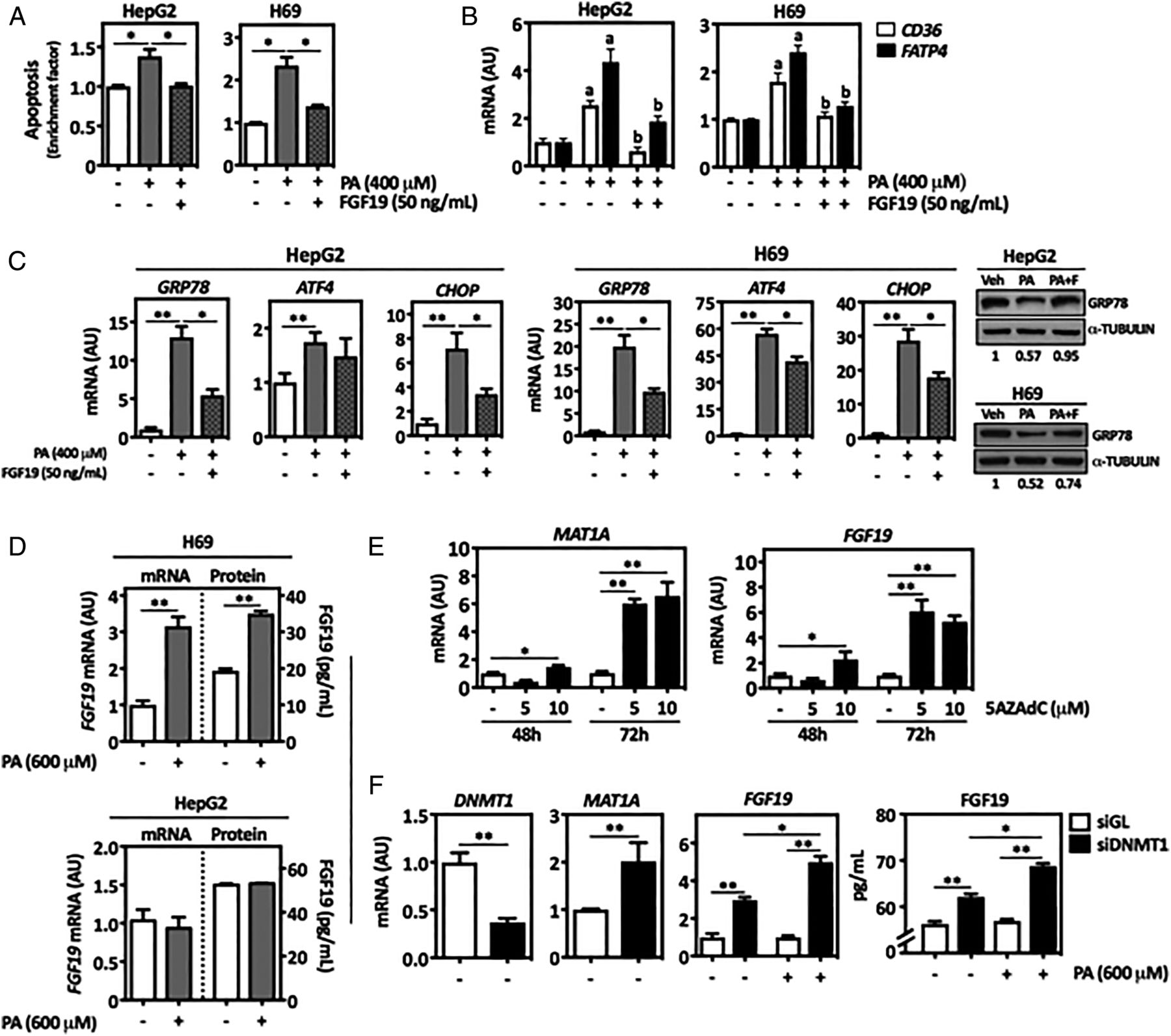
Excessive exposure of hepatocytes and cholangiocytes to saturated FAs such as PA triggers ER and oxidative stress, ultimately leading to lipoapoptosis.2 ,6 Accordingly, we found that PA-treated human cholangiocytes (H69 cells) and parenchymal cells (HepG2 cells) underwent apoptosis (figure 3A). This response, together with the activation of caspase 3, was reduced by FGF19 treatment (see figure 3A and online supplementary figure S2A). The expression of the FA transporter CD36 and the FA acyl-CoA activator fatty acid transport protein 4 FATP4 34 was induced upon PA treatment and was also downregulated by FGF19 (figure 3B). Consistent with the cytoprotective effect of FGF19, we observed that the expression of ER stress response genes triggered by PA was attenuated by FGF19 (see figure 3C and online supplementary figure S2B), while the levels of GRP78 protein were preserved (figure 3C). Interestingly, FGF19 expression was induced by PA treatment in H69 but not in HepG2 cells (figure 3D). FGF19 gene expression and inducibility are tissue-specific, and in the enterobiliary tract, it is found in the epithelial cells of the common bile duct, gall bladder and small intestine, but not in the normal liver parenchyma.35 ,36 Epigenetic mechanisms are to a great extent involved in cell type-specific gene expression. Accordingly, treatment of HepG2 cells with the demethylating agent 5-aza-deoxycytidine (5AZAdC) induced FGF19 mRNA levels (figure 3E). The expression of methionine-adenosyltransferase 1A (MAT1A), which is hypermethylated and downregulated in hepatocellular carcinoma (HCC), is shown as control of 5AZAdC treatments.37 Similar findings were obtained upon knockdown of DNA-methyltransferase 1 (DNMT1) (figure 3F). Interestingly, in the face of DNMT1 knockdown, PA treatment elicited FGF19 gene expression in HepG2 cells (figure 3F).

FGF19 protects from lipoapotosis, and its expression is induced by palmitic acid (PA) in liver cells. Human liver parenchymal cell line HepG2 and cholangiocyte cell line H69 were treated as indicated with PA (400 μM) and FGF19 (50 ng/mL) for 24 hours, and then apoptosis was measured (A). Analysis by qPCR of CD36 and FATP4 mRNA levels in HepG2 and H69 cells treated with PA and FGF19 for 24 hours as indicated above (B). Analysis by qPCR of endoplasmic reticulum (ER) stress-related genes mRNA levels in HepG2 and H69 cells treated with PA and FGF19 for 24 hours as indicated. Right panel show representative western blot analyses of GRP78 protein levels in HepG2 and H69 cells treated as indicated above. Representative blots are shown, and quantitation of band intensities (normalised to α-TUBULIN) is indicated (C). H69 and HepG2 cells were treated with PA (600 μM) or its vehicle (Veh, 1% bovine serum albumin (BSA) with <1% isopropanol) for 24 hours. Cellular FGF19 mRNA levels were measured by qPCR, and FGF19 protein levels were determined in conditioned media by ELISA (D). HepG2 cells were treated with the DNA demethylating agent 5-aza-deoxycytidine (5AZAdC) as indicated, and the mRNA levels of methionine-adenosyltransferase 1A (MAT1A) and FGF19 were measured by qPCR (E). HepG2 cells were transfected with DNA-methyltransferase 1 (DNMT1)-specific siRNAs, or with control siRNAs (siGL) for 48 hours, and where indicated cells were also treated with PA (600 μM) for another 24 hours. The expression of DNMT1, MAT1A and FGF19 mRNAs was measured by qPCR, and the levels of FGF19 protein were determined in conditioned media by ELISA (F). *p<0.05; **p<0.01; ap<0.05 vs Veh; bp<0.05 vs PA alone. AU, arbitrary units
Development of the chimaeric FGF19 variant Fibapo and evaluation of its biological activity
An initial biological characterisation of FA, a fusion protein encompassing FGF19 coupled to human ApoA-I in its C-terminus (see online supplementary figure S3A), was performed by the hydrodynamic injection to mice of plasmids encoding FA or FGF19. Serum half-life of FGF19 was markedly extended upon ApoA-I conjugation, and as expected FA was incorporated into the high density lipoproteins (HDL) fraction of serum lipoproteins (see online supplementary figure S3B, C). To demonstrate the biological activity of FA, we first generated hepatotropic adeno-associated viral vectors (AAV) harbouring FGF19, FA or ApoA-I cDNAs regulated by a hepatocyte-specific promoter (AAV-FGF19, AAV-FA and AAV-ApoA-I).38 As shown in figure 4A, serum levels of FA were significantly higher than those of FGF19 in Fgf15−/− mice. We examined the tissue distribution of FGF19 and FA and found high levels of both proteins in the liver, the main site of AAV infection, while FA was also detected in white and brown adipose tissue, muscle and brain (figure 4B). FA reproduced the biological effects of FGF19 downregulating hepatic Pparγ2 and Cyp7a1 gene expression, and reducing intrahepatic TG content and serum BA levels in Fgf15−/− mice. Infection with AAV-ApoA-I had no major effects, with the exception of Pparγ2 expression (figure 4C, D). Similar effects were observed in obese db/db mice, in which infection with AAV-FGF19 or AAV-FA also reduced Pparγ2, Mogat1, Cd36 and Fatp4 mRNA levels and intrahepatic TG content, as well as hepatic Cyp7a1 expression and BA levels (see figure 4E, F and online supplementary figure S4). Noteworthy, AAV-ApoA-I also reduced the expression of the genes involved in lipid metabolism tested here, and partially decreased hepatic TG levels (figure 4E). Interestingly, the ER stress response that develops in the liver of db/db mice, demonstrated by the low levels of GRP78 protein as described,31 was also counteracted by AAV-FGF19 and AAV-FA (figure 4G). Next, we subjected these mice to a model of acute liver failure due to extensive parenchymal resection (85% PH).21 Mice infected with AAV-FA showed significantly enhanced survival (figure 4H). We found that the circulating levels of FA measured 6 and 12 hours after 85% PH were markedly higher than those of FGF19 (see online supplementary figure S5).

Development of the FGF19-based chimaeric protein Fibapo (FA) and evaluation of its biological activity. Fgf15−/− mice were infected with adeno-associated viral vectors (AAV) expression vectors encoding FGF19 and FA cDNAs. FGF19 and FA levels were determined in serum by ELISA at the indicated time points (A) or were detected by western blotting in the indicated tissues 28 days after infection (B). Fgf15−/− mice were infected with AAV vectors encoding the control cDNA luciferase (Luc), ApoA-I (A), FGF19 (F) or FA. The hepatic levels of Pparγ2 mRNA and triglycerides (TG) were determined 28 days after infection (C). Expression of Cyp7a1 mRNA in the liver and serum bile acid (BA) levels were also measured in these mice (D). db/db mice were infected with the AAV vectors described above. Ten days after infection, the expression of the indicated genes along with TG levels were measured in the liver (E). The expression of Cyp7a1 and hepatic BA contents were also examined in these liver tissues (F). The expression of GRP78 was determined by western blotting in liver samples from control mice (db/+, heterozygous mice, Hz) and db/db mice infected with the indicated AAVs. Representative blots are shown, and quantitation of band intensities (normalised to β-ACTIN) is indicated (G). Survival of db/db mice that were infected with the indicated AAVs and 10 days later were subjected to 85% partial hepatectomy (PH) (at least 10 mice were used per condition) (H). ap<0.05 vs AAV-Luc; bp<0.01 vs AAV-Luc; cp<0.05 vs AAV-ApoA-I; dp<0.01 vs AAV-ApoA-I; ep<0.05 vs AAV-FGF19 and AAV-ApoA-I; *p<0.05 vs AAV-Luc and AAV-ApoA-I. AU, arbitrary units.
Recombinant Fibapo improves the regeneration of steatotic livers
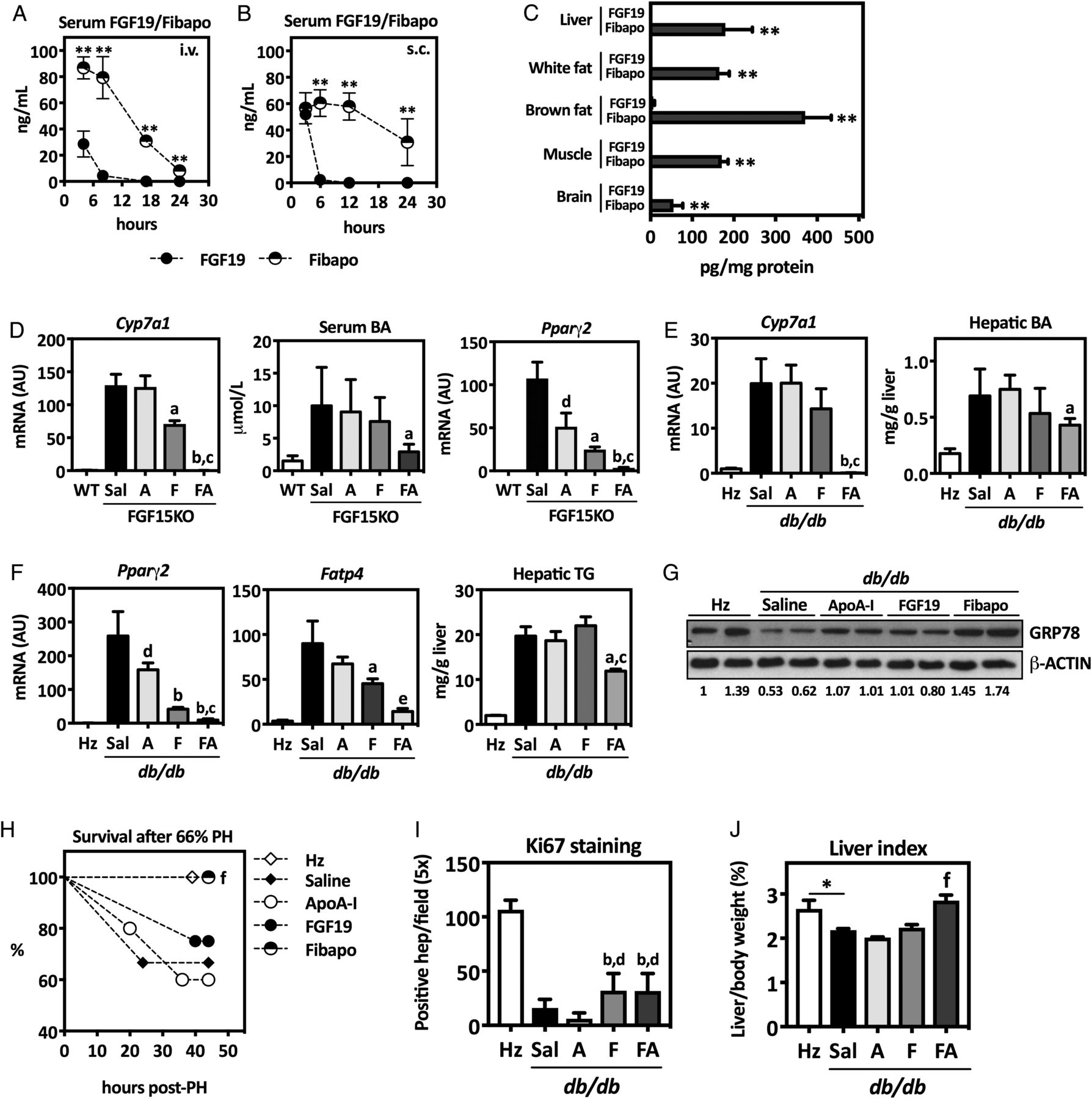
Next, we examined the pharmacological properties and potential application of recombinant FA in mouse models of fatty liver regeneration. First, we verified that after equimolar administration, the levels of FA in circulation were more stable in time than those of FGF19, both after intravenous or subcutaneous injections (figure 5A, B). Indeed, serum half-life of FA was 8.5 hours, significantly higher than that of FGF19 (1.8 hours) (see online supplementary figure S6A)24 We also analysed the tissue distribution of FA and FGF19 upon intravenous administration. Twenty-four hours after injection, FA was detected in FGF19 target tissues, including liver, brain and adipose tissues,35 while FGF19 was only detectable in brown adipose tissue (figure 5C). Similar data were obtained after subcutaneous administration of both proteins (not shown). To examine the biological activity of FA in parallel to FGF19, and ApoA-I as control, we administered a single subcutaneous injection of equimolar amounts of these proteins to Fgf15−/− mice. FA potently reduced Cyp7a1 expression and circulating BA levels, as well as Pparγ2 expression (figure 5D). We also tested FGF19 and FA in db/db mice that received three equimolar subcutaneous injections of FA, FGF19 or ApoA-I on alternate days. Forty-eight hours after the last administration, we measured the hepatic expression of Cyp7a1, Pparγ2 and Fatp4 along with the intrahepatic levels of BAs and TGs. For all these parameters, FA displayed significantly better activity than FGF19 (figure 5E, F). Interestingly, FA also improved ER stress in db/db mice livers, as indicated by GRP78 protein levels (figure 5G). To evaluate whether the beneficial effects of FA on the liver of obese and steatotic mice could improve their outcome after hepatic resection, we performed a 66% PH in db/db mice that had been treated with FGF19, FA or ApoA-I, as described above. The db/db mice show high early mortality and impaired regeneration after PH.18 We reproduced this response and found that all db/db mice treated with FA survived the intervention, while a marginal effect was observed in animals that received FGF19 (figure 5H). Survival was monitored in all groups for up to 7 days post PH without finding any additional changes (not shown). Staining of liver tissues harvested 44 hours after PH with Ki-67 revealed a marked impairment in cell cycle progression in control db/db mice compared with lean (heterozygous) animals, and this was partially recovered by FGF19 and FA treatment, but not by Apo-AI (figure 5I). Nevertheless, besides the effect of FA on mice survival, its overall proregenerative activity appeared superior to the other treatments, as indicated by liver index values 44 hours after PH that were similar to those found in lean heterozygous mice (figure 5J). FA administration also resulted in reduced intrahepatic BA concentrations and improved liver biosynthetic activity, as indicated by increased levels of serum albumin (see online supplementary figure S6B). Mechanistically, the proregenerative and trophic effects of FA may stem in part from the ability of FGF19 to stimulate protein synthesis in hepatic cells.7 Accordingly, we found increased levels of phosphorylated ribosomal protein S6 in the livers of FA-treated mice 44 hours after PH (see online supplementary figure S6C). Consistent with this observation, both FGF19 and FA, but not ApoA-I, induced the phosphorylation of p70S6kinase (p70S6K) in human parenchymal liver cells (Hep3B cells), and this response was abrogated by the FGFR4-specific inhibitor BLU9931 (see online supplementary figure S6D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Fibapo (FA) shows improved pharmacological properties than FGF19 and is effective in preventing postresection liver failure in mice with fatty livers. Serum levels of FGF19 and FA determined by ELISA at different time points after injection of equimolar amounts of both proteins (45 nmol/kg body weight) intravenously (A) or subcutaneously (B) in Fgf15−/− mice. Tissue distribution of recombinant FGF19 and FA in different mouse tissues 24 hours after intravenous administration of equimolar amounts of both proteins as determined by ELISA (C). Effect of a single administration to Fgf15−/− mice of equimolar amounts of ApoA-I, FGF19 or FA (45 nmol/kg body weight) on liver Cyp7a1 mRNA expression, serum bile acids (BA) levels and Pparγ2 mRNA levels examined 24 hours after protein injections. Values in Fgf15+/+ (wild-type, WT) mice are shown as controls (D). Effect of the administrations to db/db mice of equimolar amounts of ApoA-I, FGF19 or FA (45 nmol/kg body weight) on liver Cyp7a1 mRNA expression and BA levels examined 48 hours after protein injections. Values in db/+ (heterozygous, Hz) mice are shown as controls (E). Determination of Pparγ2 and Fatp4 mRNA and triglycerides (TG) levels in the livers of db/db mice treated as indicated above (F). Expression levels of GRP78 protein determined by western blotting in the livers of db/db mice treated with ApoA-I, FGF19 and FA as described above. Representative blots are shown, and quantitation of band intensities (normalised to β-ACTIN) is indicated (G). Heterozygous db/+ and db/db mice (N=5 mice per group) treated as indicated above underwent partial hepatectomy (PH) (66%) 48 hours after the last administration of the recombinant proteins (or saline, Sal) and survival was monitored up to 44 hours post PH (H). Quantification of Ki67 staining in liver tissue sections taken from mice 44 hours post PH (I). Liver to body weight ratio after 44 hours post PH (J). *p<0.05; **p<0.01; ap<0.05 vs ApoA-I; bp<0.01 vs ApoA-I; cp<0.01 vs FGF19; dp<0.05 vs saline; ep<0.05 vs FGF19; fp<0.05 vs all other db/db groups. AU, arbitrary units.
Discussion
In addition to its inhibitory effect on hepatic BA synthesis, FGF15/19 has been implicated in the regulation of energy homeostasis.8 ,9 ,39 It was early observed that FGF19 transgenic mice showed reduced fat content on a normal diet and were protected from diet-induced obesity.10 This phenotype was explained by enhanced energy expenditure due to increased brown adipose tissue mass, as well as by reduced lipogenesis and enhanced hepatic lipid oxidation.10 Glucose homeostasis and insulin sensitivity were also improved in FGF19 transgenic mice or upon recombinant FGF19 administration to obese mice.10 ,11 These responses apparently involve the action of FGF19 on different tissues, including adipose tissue, liver and also the central nervous system.9 The important effects of FGF15/19 on glucose metabolism, including stimulation of glycogen synthesis and inhibition of gluconeogenesis, were recently demonstrated through more physiological approaches using FGF15-null mice.7 ,33 Now we provide novel evidence on the essential role of endogenous FGF15 in fat metabolism, particularly in the adaptation to high dietary fat contents. Interestingly, under a chow diet, we found that Fgf15−/− animals gained less weight than wild types. Although we do not have a specific explanation for this difference, we may especulate that lack of FGF15/19, a relevant hormone governing nutrient metabolism, may somehow impair the physiological weight gain of young adult mice. Conversely, Fgf15−/− mice showed increased weight gain, exacerbated systemic adiposity and hepatosteatosis when fed a HFD. Previous in vitro experiments demonstrated that FGF19 counteracted the upregulation of lipogenic genes such as Srebp1 and Fas elicited by insulin in cultured hepatocytes.40 As mentioned before, we did not observe significant differences in liver Srebp1 and Fas expression between Fgf15−/− and Fgf15+/+ mice. However, we found that lack of FGF15 resulted in a marked upregulation of hepatic Pparγ2 expression, a response that was exacerbated upon HFD feeding. Pparγ2 is a transcription factor expressed at very low levels in normal human and mouse hepatocytes, induced in models of obesity and steatosis, and involved in the upregulation of adipogenic genes such as Cd36 and Mogat1.27 Importantly, hepatic overexpression of Pparγ2 and Mogat1 can drive liver steatosis even under a normal diet.27 ,41 While the influence of FGF15 on metabolic regulation in extrahepatic tissues (eg, adipose tissue) is likely relevant for systemic lipid metabolism,39 ,42 we believe that the repressive effect of FGF15/19 on liver Pparγ2 expression may be relevant for the metabolic adaptation to fat intake. In support of this, we found that exogenous FGF15/19 administration inhibited hepatic Pparγ2 expression and function (ie, Pparγ2 target genes expression) and that the expression of PPARγ2 was markedly induced in the liver of obese patients, who also show reduced FGF19 circulating levels.28 Pparγ2 promoter regulation in the liver is not completely understood, but the involvement of transcriptional regulators from the Activator Protein 1 family, as well as Small Heterodimer Partner and Hepatocyte Nuclear Factor 4α, all of them downstream targets of FGF15/19 signalling,8 has been demonstrated.43 ,44 According to our findings, FGF15/19 would be the first identified physiological signal driven by nutritional cues that can modulate hepatic Pparγ2 expression.
At variance with wild-type animals, Fgf15−/− mice developed signs of hepatic ER stress in our HFD model. Initially, the ER stress response may be an adaptive reaction to lipid overload; however, protracted ER stress has been associated with NAFLD progression. The ER is highly sensitive to excess lipids, and saturated FAs in particular promote ER stress ultimately leading to apoptosis of liver cells.1 ,2 Intriguingly, Fgf15 expression can be triggered by non-physiological activators of the ER stress response in mouse ileum.32 We found that acute feeding of a PA-enriched lipid mixture induced a potent ileal ER stress response and sustained Fgf15 expression, which was followed by signs of hepatic ER stress. It was recently demonstrated that the hepatic expression of β-Klotho, which together with FGFR4 forms the FGF15/19 receptor complex in hepatocytes,35 is induced in response to ER stress.45 Collectively, these observations suggest that upon increased fat intake, the enhanced release of FGF15/19 from the gut into the portal circulation, together with the induction of β-Klotho expression in hepatocytes, could facilitate the adaptation of hepatic metabolism to dietary lipid excess in a coordinated manner. Additionally, FGF15/19 has direct cytoprotective effects, as demonstrated in HCC cells.46 We found that FGF19 protected HepG2 and H69 cells from lipoapoptosis induced by saturated FAs. The mechanisms involved in this antiapoptotic activity can be multifarious, since FGF15/19 besides a hormone is also a growth factor that activates intracellular survival pathways.8 Nevertheless, we found that the upregulation of the FAT CD36 upon PA challenge was inhibited by FGF19 in both cell types. Overexpression of CD36 is associated with increased steatosis and apoptosis in the liver of patients with NAFLD.47 Perhaps, a more compelling mechanistic finding was the repression of FATP4 by FGF19 treatment. FATP4 expression is induced in the fatty liver, localising to the ER and mitochondria. This acyl-CoA synthetase catalyses the incorporation of saturated FAs into ER stress-inducing phospholipids and their conversion into other lipotoxic metabolites, mediating to a great extent PA-induced hepatocyte apoptosis.34 Consistently, in addition to enhanced cell survival, we also found reduced expression of ER stress marker genes in PA-treated cells that received FGF19. Interestingly, we observed that PA induced the expression of FGF19 in H69 cholangiocytes, but not in the hepatic parenchymal cell line HepG2. This response could represent a protective autocrine/paracrine loop to avoid cholangiocyte lipoapoptosis,6 similar to the upregulation of FGF19 in biliary epithelial cells in response to excessive BA concentrations.36 ,48 ,49 FGF19 gene expression seems to be differentially controlled in human biliary cells and hepatocytes. FGF19 behaves as a readily inducible gene in cholangiocytes compared with differentiated parenchymal cells, in which it is normally a silenced gene.36 ,50 Epigenetic mechanisms such as DNA methylation greatly influence cell type-specific gene expression. Accordingly, we found that inhibition of DNA methylation promoted FGF19 expression in HepG2 cells and made this gene responsive to PA stimulation. Considering that HCCs are characterised by global DNA hypomethylation, our findings could have broader implications, as they may contribute to explain the overexpression of FGF19 in a significant proportion of HCCs in which the FGF19 gene is not amplified.46 Moreover, these observations would be consistent with the recent finding of increased hepatocellular FGF19 expression in patients with chronic liver disease and steatohepatitis.51
The complement of biological activities encompassed by FGF19 including the downregulation of hepatic BA synthesis and lipid accumulation, its mitogenic and cytoprotective effects, and its capacity to stimulate protein synthesis could make FGF19 an ideal therapeutic molecule in the context of liver regeneration. Certainly, cholestasis and steatosis are factors that negatively influence organ recovery and regeneration after PH or transplantation in humans.13–16 Moreover, increased ER stress and PA overload are observed in steatotic livers after experimental PH, and modulation of ER stress improves regeneration in fatty livers.19 ,20 Therefore, the protective effects of FGF15/19 described here on steatosis-associated ER stress, as well as on PA-induced cell death, may be of particular relevance in this context. However, as we and others have observed, FGF19 has a very short half-life in circulation.24 To overcome this limitation and to enhance FGF19 hepatotropism, we developed FA. In addition to fulfilling these expected kinetic and biodistribution properties, FA retained FGF19 inhibitory activities on BA synthesis and also reduced hepatic Pparγ2 expression and TG contents. Its administration to obese mice improved survival after extensive (lethal) PH, and enhanced regeneration and promoted survival upon standard (66%) PH. These effects were observed when FA was expressed from AAV vectors, suggesting its potential administration using gene therapy strategies, and also when it was delivered as a recombinant protein. Interestingly, FA showed better therapeutic activity than FGF19 in the different models of liver steatosis and regeneration tested in our study. While part of this improved performance may be attributed to the enhanced pharmacokinetic properties conferred by ApoA-I, this moiety could also contribute to the therapeutic effects of FA. In fact, ApoA-I administration has been shown to reduce hepatosteatosis and to improve liver histology in obese mice, which is consistent with our observations in db/db mice.52 Moreover, the interaction of ApoA-I with its receptors SR-BI and ABCA1 expressed in hepatocytes triggers a variety of prosurvival signalling mechanisms.52 On the other hand, interaction of HDL-residing FA with SR-BI receptors through its ApoA-I moiety might also result in its internalisation in the cells. This might compromise the binding and activation of the FGFR4/β-Klotho complex, leading to a reduced biological activity. Nevertheless, while this situation may indeed occur, our in vivo observations still demonstrate a better pharmacological performance of FA compared with FGF19.
In summary, we have provided novel evidences on the important physiological role played by FGF15/19 in liver fat metabolism, and in protection from lipid-mediated cellular stress and injury. Our observations demonstrated that the biological activities present in FGF15/19 can be harnessed in a new molecule termed Fibapo with improved kinetic and pharmacological properties. As recently proposed for growth hormone,53 the perioperative administration of FA, alone or in combination with other strategies such as weight loss, could improve the outcome of patients with steatotic organs that need resection.
Acknowledgments
The authors thank Dr Laura Guembe, from CIMA Morphology core facility, for histological and immunohistological analyses.
References
Footnotes
MAA and CB share senior authorship. GA-S and IU made equal contribution to the study.
Contributors GA-S, IU, MUL, RU, ME, MB-V, MJ, HCC and CR-O performed in vitro and in vivo experiments and collected data. VC, AR, JMG-E and AG-N analysed human samples and collected data. GA-S, IU, FJC, JP, PB, FV, GF, CB and MAA designed experiments and discussed the data. IU, CB and MAA wrote and submitted the manuscript.
Funding This work was funded by CIBERehd; Grants FIS PI13/00359, PI13/00385 and PI16/01126 from Instituto de Salud Carlos III (ISCIII), co-financed by ‘Fondo Europeo de Desarrollo Regional’ (FEDER) ‘Una manera de hacer Europa’. ‘Adipoplast’ Network (BFU2015-70454-REDT). ‘Ramón y Cajal-I3’ contract to MUL. Marie Curie EU contract to MGF-B. ADA-University of Navarra fellowship to MJ; FPI fellowship from Ministerio de Economía to MB-V; Fundación Eugenio Rodríguez Pascual; Fundación M Torres; Fundación Mario Losantos; Fundación Familia Puig-Infante. We also thank Mr. Eduardo Avila Zaragozá for his generous support.
Competing interests Not declared.
Ethics approval The University Clinic of Navarra and the Santa Creu i Sant Pau Hospital Ethics.
Provenance and peer review Not commissioned; externally peer reviewed.