Article Text

Abstract
Objective: Liver regeneration is mainly based on cellular self-renewal including progenitor cells. Efforts have been made to harness this potential for cell transplantation, but shortage of hepatocytes and premature differentiated progenitor cells from extra-hepatic organs are limiting factors. Histological studies implied that resident cells in adult liver can proliferate, have bipotential character and may be a suitable source for cell transplantation.
Methods: Particular cell populations were isolated after adequate tissue dissociation. Single cell suspensions were purified by Thy-1 positivity selection, characterised in vitro and transplanted in immunodeficient Pfp/Rag2 mice.
Results: Thy-1+ cells that are mainly found in the portal tract and the surrounding parenchyma, were isolated from surgical liver tissue with high yields from specimens with histological signs of regeneration. Thy-1+ cell populations were positive for progenitor (CD34, c-kit, CK14, M2PK, OV6), biliary (CK19) and hepatic (HepPar1) markers revealing their progenitor as well as hepatic and biliary nature. The potential of Thy-1+ cells for differentiation in vitro was demonstrated by increased mRNA and protein expression for hepatic (CK18, HepPar1) and biliary (CK7) markers during culture while progenitor markers CK14, chromogranin A and nestin were reduced. After transplantation of Thy-1+ cells into livers of immunodeficient mice, engraftment was predominantly seen in the periportal portion of the liver lobule. Analysis of in situ material revealed that transplanted cells express human hepatic markers HepPar1 and albumin, indicating functional engraftment.
Conclusion: Bipotential progenitor cells from human adult livers can be isolated using Thy-1 and might be a potential candidate for cell treatment in liver diseases.
Statistics from Altmetric.com
In response to liver injury, hepatic cells, especially hepatocytes, can rapidly proliferate to repair liver damage.1 After hepatectomy hepatocytes are able to undergo numerous cell divisions maintaining their fully differentiated state to compensate for hepatocyte loss, whereas undifferentiated liver progenitor cells play only a minor role in this process. Interestingly, under conditions such as some chronic liver diseases and after acute necrosis these hepatocyte progenitors play an important role, in particular when growth response of mature hepatocytes is impaired.1 2
Mechanisms leading to progenitor cell activation have been studied in detail using rodent models of hepatocarcinogenesis and liver regeneration after toxic injury.3 4 Under these conditions increasing numbers of small cells (∼10 μm) were observed having a large nuclear-to-cytoplasm ratio with an oval-shaped nucleus (oval cells). They express markers of both hepatocyte lineages and the biliary epithelium that includes bile ducts as well as the canal of Hering.5 6 Although histochemical studies suggest that epithelial cells residing within the canal of Hering might be resident liver stem cells, the precise origin remains ambiguous.6 In humans these cells are termed progenitor cells/ductular reaction7 and are activated in different diseases such as chronic viral hepatitis, alcoholic liver disease, non-alcoholic fatty liver disease and cirrhosis.4 8 They are observed in the portal region of the hepatic lobule9–11 and, furthermore, their numbers increase in direct proportion to disease severity.12
Hepatic progenitor cells have also been shown to express haematopoietic stem cell (HSC) markers CD34, c-kit (CD117)13 and Thy-1 (CD90).14 Accordingly, it was hypothesised that these cells are haematopoietic stem cells transdifferentiated into the hepatic lineage.15 However, recent reports have demonstrated that cell fusion rather than transdifferentiation is involved in this process. Also, it was shown that rodent oval cells do not originate from bone marrow,16 but bone-marrow-derived stem cells may contribute to liver regeneration in an as yet unknown mechanism.17
Isolation procedures for hepatic progenitor cells were established for rodents and in vitro studies have shown that these cells are capable of differentiating into hepatocytes and bile duct cells under appropriate conditions.18 Isolation of liver progenitor cells from human material has proven more difficult. Indeed, only a few reports describe an isolation of progenitor cells from fetal livers. These cells expressed various lineage specific antigens including biliary and hepatocellular markers.19 In adult human liver, progenitor cells have been identified by CD34 and c-kit immunoreactivity,13 but the so-called side-population was also found.20 Recently, it was documented that Thy-1 (CD90) is expressed in human hepatoblastoma.21 This surface antigen has also been useful for successfully isolating progenitor cells from human fetal liver.14 However, to our knowledge, there have been no attempts so far to use this marker for the separation of progenitor cell populations from adult human liver tissues. Since such cell populations may become central determinants for future treatments of liver diseases requiring clinically relevant liver regeneration, we aimed to identify and isolate Thy-1+ cells from adult liver tissue for cell transplantation purposes. We analysed the phenotypic characteristics of Thy-1+ human progenitor cells after isolation, studied their differentiation potential in vitro and monitored their repopulation capacity in vivo in a mouse model of liver regeneration.
MATERIALS AND METHODS
Reagents and material
Antibodies for this study are summarised in table 1. Mouse anti-rabbit IgG, rabbit anti-mouse IgG, and mouse APAAP were obtained from Dako (Hamburg, Germany). Texas red-conjugated donkey anti-rabbit antibody and fluorescein isothiocyanate (FITC)-conjugated donkey anti-mouse antibody were obtained from Jackson Immunoresearch (Newmarket, UK). RNase A was purchased from Qiagen (Hilden, Germany), DNase I and fast red substrate from Roche Diagnostics (Mannheim, Germany). All further chemicals were purchased from Sigma (Taufkirchen, Germany) or Merck (Darmstadt, Germany).
Tissue samples from human liver resections were obtained from patients undergoing partial hepatectomy. Experimental procedures were performed according to the guidelines of the charitable state-controlled foundation HTCR (Human Tissue and Cell Research), with the informed patient’s consent approved by the local ethics committee of the University of Regensburg.22 A total of 39 liver specimens were categorised into regenerating (n = 20; age 53.7 (SD 15.6) years; male 13, female 7) and non-regenerating (n = 19; age 61.7 (SD 12.2) years; male 15, female 4) tissue according to a histological and clinical examination by specialist. Non-regenerating liver was assigned to patients with metastatic liver tumours of colorectal cancer and regenerating liver to patients with disease as following: fibrosis/cirrhosis (n = 4; age 53.0 (SD 9.3) years; male 4), fibrosis/non-alcoholic steatohepatitis (NASH) (n = 4; age 58.0 (SD 11.2) years; male 2, female 2), fibrosis/alcoholic steatohepatitis (ASH) (n = 1; age 23; female), cirrhosis/ASH (n = 2; age 64.5 (SD 4.5) years; male 2), steatosis (n = 7; age 53.7 (SD 19.7) years; male 3, female 4), ASH (n = 1; age 49 years; male), NASH (n = 1; age 52 years; male).
Isolation of Thy-1 positive cells from human liver tissue
Isolation of non-parenchymal cells
Liver tissue was digested using a modified two-step egtazic acid (EGTA)/collagenase perfusion procedure23 to isolate human hepatocytes. Non-parenchymal cells (NPCs) were obtained from the remaining liver tissue based on a protocol described recently.24 Briefly, up to 50 g of tissue was minced and incubated for up to 150 min under vigorous shaking at 37°C in 50 ml isolation buffer (154 mmol/l NaCl, 5.6 mmol/l KCl, 5 mmol/l glucose, 25 mmol/l NaHCO3, 20 mmol/l HEPES, pH 7.4) supplemented with 0.5 mg/ml collagenase type IV, 0.5 mg/ml pronase, 40 μg/ml DNase I, 5 mmol/l CaCl2 and 1 mmol/l MgCl2. Every 30 min undigested tissue was separated, minced and incubated again. The resulting suspension was filtered (meshes with 210 μm and 170 μm pore sizes) and washed three times in phosphate-buffered saline (PBS) containing 2 mmol/l ethylenediaminetetraacetic acid (EDTA). Furthermore, the cell suspension was purified using a 70% Percoll solution (800 g, 30 min, 18°C). After washing with PBS/EDTA, the NPCs were suspended in MACS buffer (PBS containing 2 mmol/l EDTA, 5 g/l bovine serum albumin (BSA)) and stored for up to 20 h at 4°C. Size profile of cell fractions was monitored for quality assessment of the dissociation procedure using a cell counter system (Casy Schärfe System, Reutlingen, Germany). NPC fractions were isolated with an average yield of 2.7 (SD 2.6)×106 viable cells per g wet tissue (n = 39).
Purification of Thy-1 positive cells
Thy-1+ cells were isolated from NPCs using the immuno-magnetic activated cell sorting (MACS) technique (Miltenyi Biotech, Bergisch Gladbach, Germany) according to the instruction manual. Briefly, cells were filtered (40 μm filter, BD Biosciences Europe, Erembodegem, Belgium) and 5×107 cells were incubated with 100 μl of a mouse anti-CD90 (Thy-1) monoclonal antibody (mAb) for 30 min at 4°C. After twofold washing with MACS buffer, cells were incubated with 100 μl of goat anti-mouse microbeads for 15 min at 4°C followed by two additional washing steps. Labelled cell suspensions were transferred onto MACS LS columns and washed three times while being located in the magnetic field. Thy-1+ cells were eluted with 5 ml MACS buffer following removal from the magnet. Cell number and viability were determined by the trypan blue exclusion technique.
Characterisation by flow cytometry
Cells were suspended in PBS, incubated with fluorescence-labelled mAb for 30 min at 4°C, washed with PBS and analysed following addition of propidium iodide (PI) (1 μg/ml PI per 1 μg/ml RNase A) on a FACSCalibur flow cytometer (BD, Heidelberg, Germany) using appropriate regions of interest and/or quadrant statistics in the compensated fluorescence dot plot diagrams. PI positive, membrane defect cells were excluded. Details concerning the antibodies are given in table 1.
Immunohistochemical staining
Cryosections (6 μm) from snap frozen tissues were fixed in ice-cold acetone (10 min). For cytospots freshly isolated cells were suspended in PBS, centrifuged (5×104 cells) onto glass slides (SuperFrost Plus; Merck) and fixed in acetone. Cultured cells were fixed using 4% paraformaldehyde for 20 min.
Alkaline phosphatase anti-alkaline phosphatase (APAAP) staining
Blocking of slides was performed with 3% fetal calf serum (FCS) in PBS followed by incubation with primary antibodies for 30 min and a washing step with PBS. For CK14 staining, additional incubation with mouse anti-rabbit immunoglobulins for 30 min was performed, followed by incubation with rabbit anti-mouse immunoglobulins for 30 min. After incubation with mouse APAAP (1:100 in PBS/1% FCS) for 30 min immunosignals were developed with fast red substrate (20 min) and counterstained with haematoxylin Quick Stain.
Immunofluorescence staining
After blocking (3% dried milk, 0.1% Triton-X100, PBS) cells were incubated with primary antibodies for 30 min, washed with PBS and incubated with the secondary fluorescence-conjugated antibodies. For co-localisation, steps were repeated with a second set of antibodies. Nuclei were counterstained with 5 μg/ml Hoechst 33258 in PBS.
Determination of mRNA expression by reverse transcription-polymerase chain reaction
Total RNA was extracted using the RNeasy Mini Kit (Qiagen). First strand cDNA was synthesised using 0.1 μg of total RNA and the avian myeloblastosis virus (AMV)-reverse transcription reaction (Promega, Mannheim, Germany) using 15 U AMV reverse transcriptase in a prolonged incubation time of 60 min. The polymerase chain reaction (PCR) was performed according to the manufacturer’s instructions (Invitrogen, Karlsruhe, Germany) using primers as listed in table 2. After an initial denaturation step of 3 min at 94°C, amplification was performed for 35 cycles at 94°C for 45 s, 30 s at the optimal annealing temperature and 72°C for 30 s. The PCR was terminated by a final elongation step at 72°C for 10 min. Amplified products were transferred on 1.5% agarose gels for electrophoresis followed by ethidium bromide staining.
Cell culture
Freshly isolated Thy-1+ cells were resuspended in adhering medium (William’s E medium, 5% FCS, 10 mmol/l nicotinamide, 20 mmol/l HEPES, 17 mmol/l NaHCO3, 550 mg/l sodium pyruvate, 0.2 mmol/l ascorbic acid 2-phosphate, 14 mmol/l glucose, 10−7 mol/l dexamethasone, 6.25 μg/ml insulin, 6.25 μg/ml transferrin, 6.25 ng/ml sodium selenite, 1.25 mg/ml BSA and 5.35 μg/ml linoleic acid, 100 U/ml penicillin, 100 μg/ml streptomycin) and plated onto collagen coated dishes (BD Biosciences Europe) at a density of 5×104/cm2. After 3–5 days the medium was replaced with culture medium (adhering medium without FCS, 20 ng/ml epidermal growth factor (EGF)). The medium was changed once a week. Viability and cell growth during the culture period were monitored by cell morphology.
Transplantation of hepatic progenitor cells
All animal housing and experimentation procedures were in accordance with German legislation on animal protection (latest version). Pfp/Rag2−/− mice (malfunction of NK cell, T and B cell depletion) were obtained at 8 months of age from Taconic (Lille Skensved, Denmark). The cellular transplantation model was performed as described recently.25 26 Briefly, recipient liver regeneration was inhibited by treating the mice with monocrotalin (30 mg/kg body weight; intraperitoneally) 6 and 4 weeks prior to transplantation. After partial hepatectomy, 1×106 human Thy-1 positive liver cells were administered by intrasplenic injection. Livers were removed after 7 weeks and fixed in 4% formaldehyde and paraffin embedded for further analysis.
RESULTS
Expression of progenitor cell marker in human liver sections
Expression analysis of markers described for liver progenitors were performed in cryo-sections of adult liver tissue from a patient with fat liver disease. Immunoreactivity for CD34 was found in endothelial cells lining the vascular structures of the portal tract and sparely on cells in the adjacent parenchyma (fig 1A), whereas c-kit expression was barely detected (fig 1C). Thy-1 expression was found in the portal tract area and in scattered cells in the surrounding parenchyma (fig 1B). Expression of the oval cell marker OV6 was detected on epithelial cells adjacent to the portal tract, but primarily on bile duct cells (fig 1D). Specific immunostaining of CK19 for biliary epithelial cells (fig 1E), CK18 for bile duct epithelium and parenchymal hepatocytes (fig 1F) is also demonstrated.
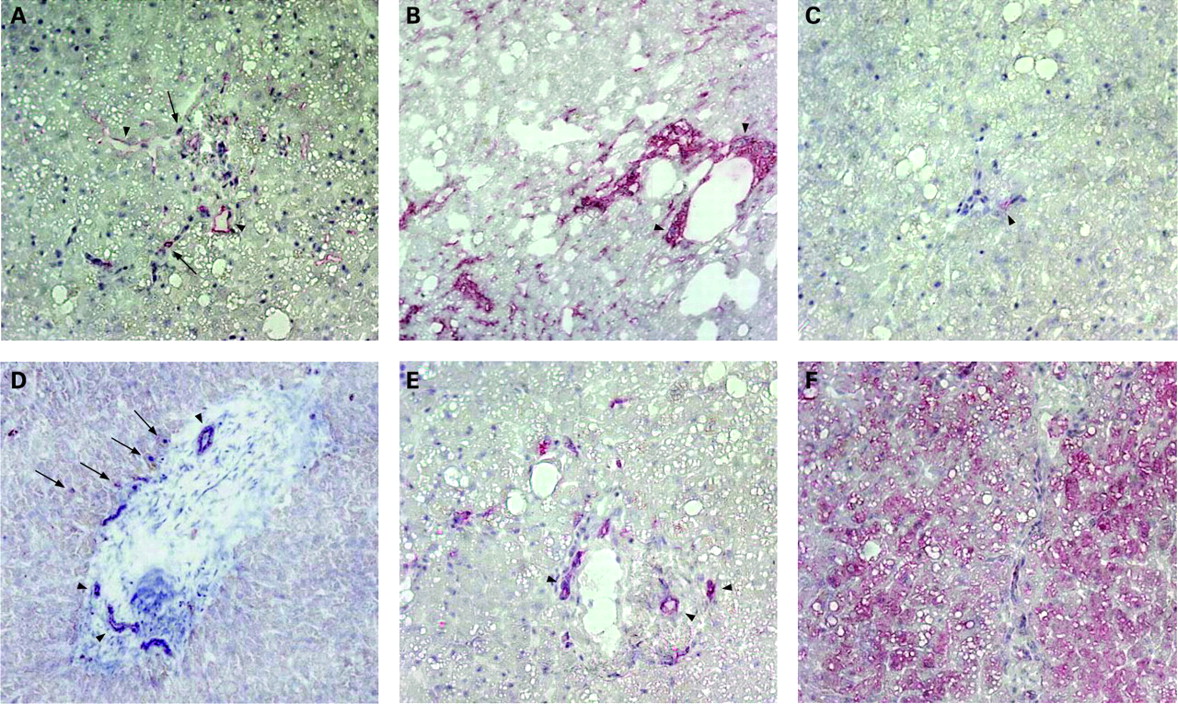
Cells of adult human non-parenchymal liver cell fractions express stem cell markers
Fractions of small liver cells separated from hepatocytes were isolated from discarded surgical adult human liver tissue after performing a multi-step isolation process (fig 2A–B). Immunocytochemical analysis of small liver cell fractions revealed a high percentage of non-parenchymal cells (Kupffer, stellate and endothelial cells; data not shown) and are therefore termed the non-parenchymal cell (NPC) fraction. NPC fractions were investigated by flow cytometry for expression of surface markers, which have been detected on liver progenitor cells by immunohistochemistry. Cells were found positive for CD34 (6% (SD 4%), n = 18), CD117 (8% (SD 7%), n = 18) and CD90 (9% (SD 6%), n = 22). Additionally, double immunostaining revealed, that 2% (SD 2%) (n = 11) of NPC fractions were CD34+/CD90+, 2% (SD 2%) (n = 9) were CD34+/CD117+ and 3% (SD 1%) (n = 5) were CD90+/CD117+.

Thy-1 positive cells express markers of liver progenitor cells
Thy-1 (CD90), the most abundant progenitor marker on NPC fractions, was selected as a marker for immuno-magnetic activated cell sorting (MACS). Subsequently, fluorescence-activated cell sorting (FACS) analysis was performed determining the quantity and characteristics of Thy-1 isolated cells. MACS sorted cells were highly positive for Thy-1 (83% (SD 7%), n = 6) and co-expressed CD34 (24% (SD 9%), n = 5) and hardly CD117 (1.8% (SD 0.7%), n = 5) (fig 2C–E). Additionally, we found expression of CD45 (35% (SD 9%), n = 5), CD14 (20% (SD 7%), n = 5) and CD3 (29% (SD 9%), n = 5) on some Thy-1+ cells. The overall outcome of viable Thy-1+ cells after MACS isolation was 1.4 (SD 0.9)×104 per 106 NPCs (n = 39). Furthermore, performing a correlation study we found a significant higher rate of Thy-1+ cells in tissues, in which regenerative events; for example, slight histological signs of hepatitis, fibrosis, or cirrhosis, were described (1.8% (SD 1.0%) of NPCs, n = 20) than in non-regenerating tissue (1.2% (SD 0.57%) of NPCs, n = 19) (fig 2G). The rates of Thy-1+ cells were not correlated with expression of CD45, CD14 and CD3 (data not shown). A more detailed analysis with type of disease, tissue damage and patient related data did not show any correlation to the rate of Thy-1+ cells (data not shown).
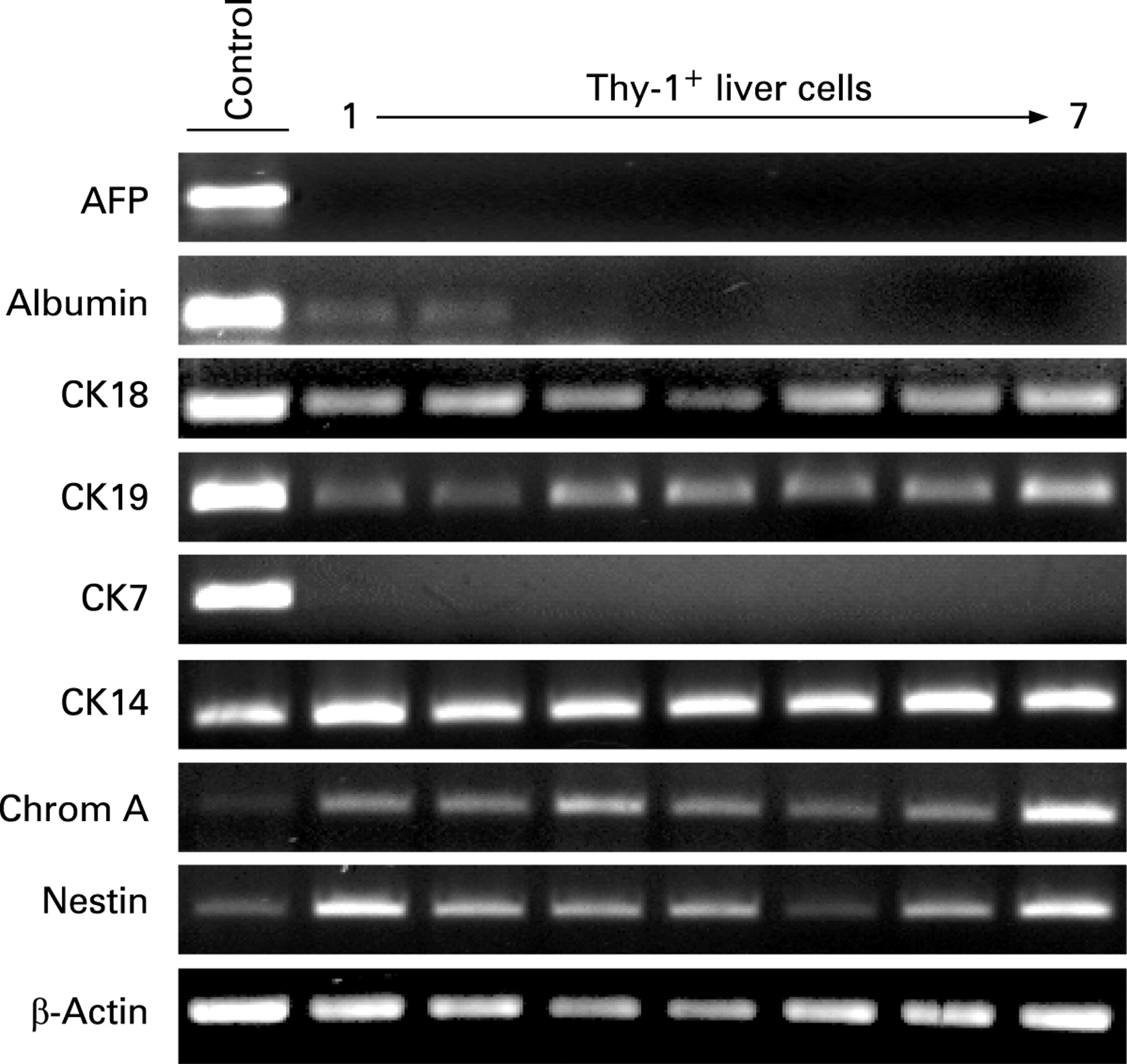
Expression of progenitor and hepatic cell markers were determined on freshly isolated Thy-1+ cells by immunostaining (fig 3). About 60% of the Thy-1+-sorted cells demonstrated positivity for liver progenitor markers CK14, M2PK, OV6 and biliary/progenitor marker CK19 (fig 3A). Double immunofluorescence staining of Thy-1+ cells revealed co-localisation of CK14 with CD117, CD34 (liver progenitor and HSC), CK19 (bile duct/progenitor cell), HepPar1 (hepatocyte) and OV6 (fig 3B). mRNA analyses of the Thy-1+ cell population from 27 donors confirmed the expression of CK14. Also, we detected two additional progenitor genes expressed in Thy-1+ cells of all donors, chromogranin A (pancreatic) and nestin (neural) (fig 4). Positive gene expression was also found for hepatocyte markers CK18 and albumin (10 of 27), as well as biliary marker CK19, but not for CK7. alpha-fetoprotein (AFP) was not detected in any of the cell preparations (fig 4).


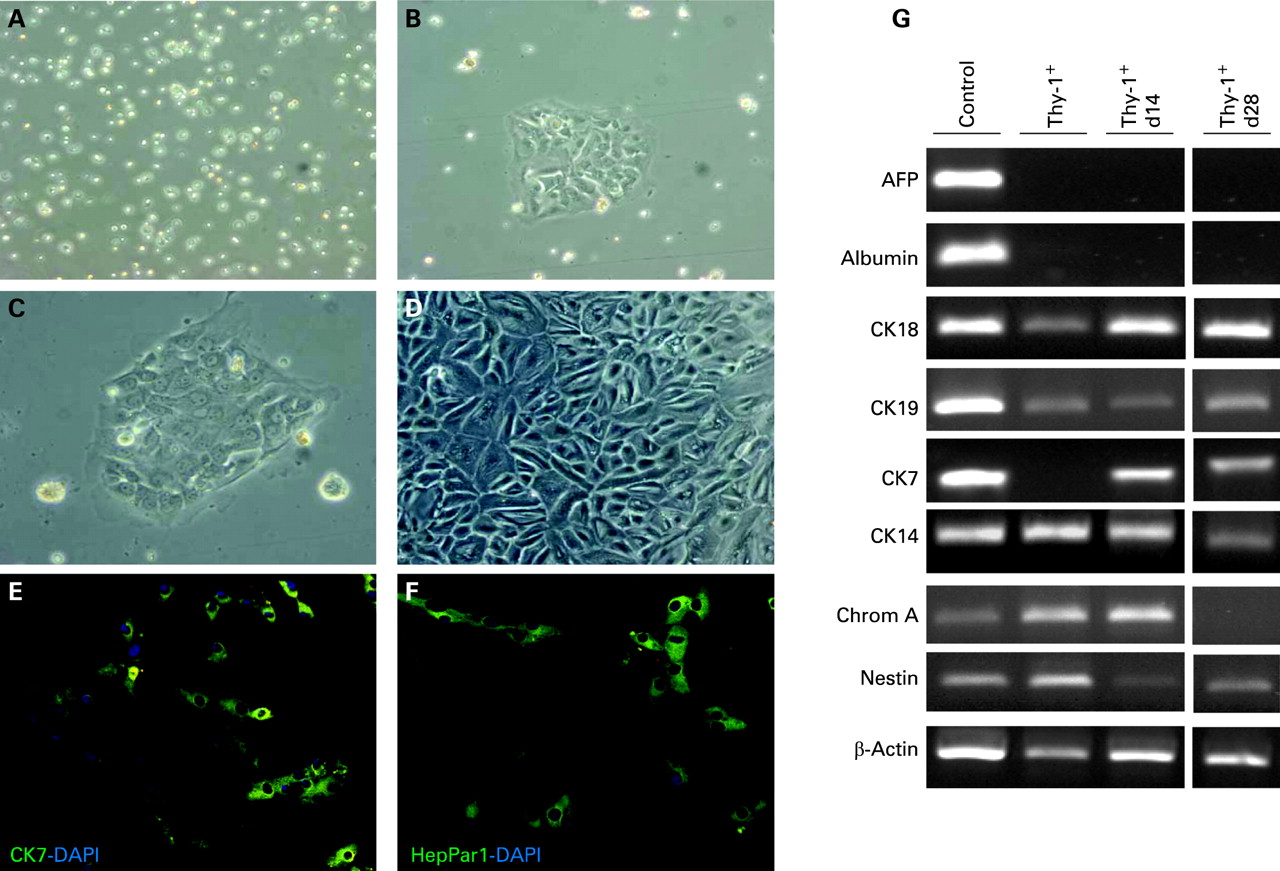
Thy-1 positive liver cells in culture show differentiation towards hepatocytes and bile duct cells
Thy-1+ cells were seeded on collagen-coated culture dishes and maintained in culture under conditions described for hepatocyte growth.27 Efficiency of plating was sufficient and after 10–14 days cells started to grow into colonies (fig 5A–C). At day 28 of culture an almost complete monolayer was formed (fig 5D). Furthermore, mRNA analyses by reverse transcription polymerase chain reaction (RT-PCR) were performed throughout culturing and differences in gene expression were observed depending on the culture period (fig 5G). The hepatic marker CK18 strongly increased from day 14 to 28, whereas albumin could not be detected. Furthermore, CK19 expression was almost unchanged up to day 28, while the gene expression of the bile duct marker CK7 was clearly enhanced. In contrast, a reduction of the progenitor markers CK14, chromogranin A and nestin was observed as a function of time in culture. AFP was not expressed either in fresh or in cultured cells. In addition, Thy-1+ cells cultured for 14 days were stained with antibodies against HepPar1 and CK7 and demonstrated expression of hepatocyte and bile duct specific proteins, respectively (fig 5E,F).
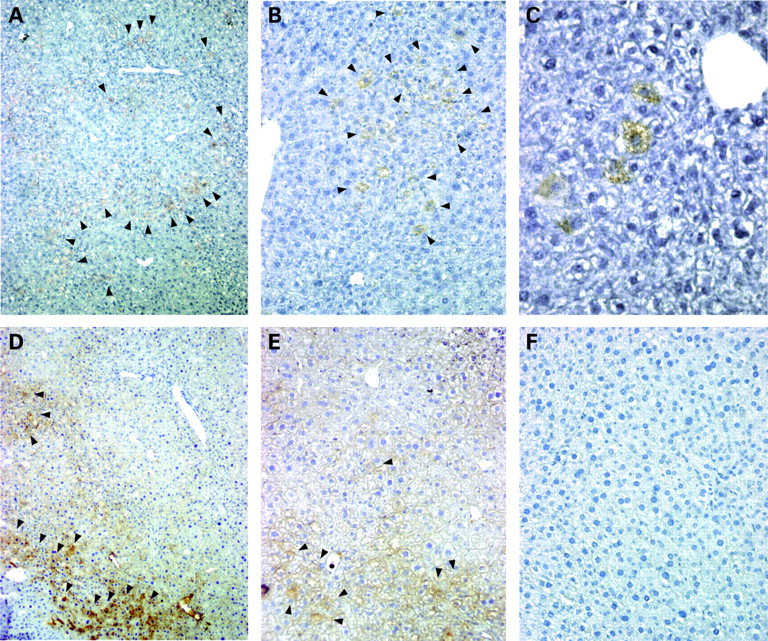
Transplanted Thy-1 positive cells demonstrate engraftment in mouse livers
Integration of transplanted Thy-1+ cells was studied using a mouse transplantation model, which impedes proliferation of residual hepatocytes after partial hepatectomy and therefore supply transplanted Thy-1+ cells with a growth advantage over host hepatocytes.25 Seven weeks after transplantation human hepatic cells were detected in the mouse liver background by staining with HepPar1 (human hepatocyte specific antigen) antibody (fig 6A–C), which showed no relevant cross-reactivity for murine antigens. In cryosections of the respective materials, human albumin, a marker of functional activity and hepatocyte differentiation was also detected (fig 6D,E). HepPar1 and albumin positive cells were localised in the periportal region, demonstrating a predominant periportal engraftment of transplanted Thy-1+ liver cells. After transplantation into mouse liver Thy-1+ sorted cells featured a typical human hepatocytic phenotype indicating integration of the transplanted cells within the mouse liver.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
The objective of this paper was to determine whether Thy-1 positive cells (1) were found in and could be isolated from human liver tissue; (2) demonstrate multiple lineage markers common with a progenitor phenotype; and (3) have the potential for cellular treatment in liver regeneration. The data presented show that adult human liver harbours Thy-1 positive cells which express hepatocellular and biliary markers representing their bipotential character. These cells have a repopulation capacity after transplantation and therefore resemble a liver progenitor cell type.
Because of their anatomical homology, both adult and fetal livers have been considered to harbour progenitor cells in the ductules/canals of Hering at the interface between the parenchymal and the portal tract mesenchyme.6 28 These progenitor cells were defined in humans based on immunohistochemical studies as larger than 6 μm and less than 40 μm in diameter,7 which agrees with the isolated progenitor cell fraction (10 (SD 3) μm) in this study and, furthermore, might correspond in size to progenitor cells isolated from rat (8–10 μm)29 and mouse (7–15 μm).30 The isolated small liver cells partly showed co-expression of CD34, c-kit and Thy-1, which are markers of the haematopoietic compartment and were described as being present in human13 14 and rodent liver.31 Furthermore, we found some Thy-1+ cells positive for CD45 which was also reported earlier by others using human fetal14 and murine liver.30 CD45 positivity, in general, indicates a haematopoietic lineage,32 but recent reports clearly show that hepatic side-population cells, a further characteristic of progenitor cells,33 both CD45 positive and negative, can differentiate into hepatic cells expressing HepPar1 and albumin20 and upon transplantation into a mouse model, can contribute to liver regeneration.34 Therefore, the CD45 surface antigen expression status is suggested as not important in terms of the hepatic differentiation potential. Additionally, by using immunohistochemistry we found Thy-1 and CD34 expressed on endothelial cells lining the vascular structures of the portal tract which might correspond to a stem cell niche of the mesenchymal compartment giving rise to mesodermal and epithelial tissue as suggested earlier.14
Thy1+ cells in human liver feature bipotential progenitor cells with hepatic characteristics
Progenitor cells are often recognised by expression of organ specific fetal proteins, which demonstrate their early status in development. Isolated human Thy-1+ cells showed specific expression of M2PK, the fetal M2 isoform of pyruvate kinase (M2PK), which is considered as a marker for liver progenitor cells, not expressed in adult hepatocytes and ductal cells.12 Furthermore, we did not find mRNA expression of AFP, a serum glycoprotein produced by hepatoblasts during the fetal development, a defining feature of progenitor cells in many rodent models of injury7 and in human fetal livers.14 On the other hand, it was reported that AFP in progenitor cells from adult human liver was only occasionally observed7 and it was recently shown that during progenitor cell activation in a rat model, the non-ductular progenitor cells expressing nestin and Thy-1 were negative for AFP.35
A characteristic of liver progenitor cells is the co-expression of different cell specific proteins enabling a differentiation into hepatic and/or biliary epithelial cells. We could demonstrate that the biliary marker CK19 as well as the oval cell marker OV636 were not only detected on bile duct cells by immunohistology, but also were present on Thy-1+ cells indicating these markers as liver progenitor markers as suggested before for CK197 and OV6.37 The bipotential nature of Thy-1+ cells is shown by expression of CK19 and HepPar1, antigens identified on liver progenitor cells during early stages of gestation.38 Enhanced or reduced expression of either markers commits to biliary or hepatocyte epithelial cell lineages. HepPar1 antibodies recognise the mitochondrial fraction of human hepatocytes and do not label haematopoietic or non-parenchymal cells.39 Further evidence suggesting Thy-1+ cells as bipotential liver progenitors comes from our RT-PCR analysis which showed expression of the biliary lineage marker CK19 as well as hepatocytic lineage marker CK18 and partly albumin. Additionally, Thy-1+ cells revealed expression of the neuroendocrine marker chromogranin A28 and neural progenitor cell marker nestin,40 both suggested as marker proteins of bipotential liver progenitors. Expression of these markers has also been seen on differentiated cells from the haematopoietic lineage and therefore may indicate a relationship between the haematopoietic system and liver cell lineage either originating from or both having a common ancestor during the fetal liver development when haematopoietic cells colonise the liver.16 Furthermore, CK14 has been described as a marker of bipotential liver progenitors and was detected in human regenerating livers after massive necrosis.38 41 Although it was reported that OV6 antibodies in rat liver recognise an epitope common to both CK19 and CK14 present on rodent progenitor cells,42 in human tissue controversial data about specificity are reported describing expression patterns of OV6 and CK19 as non-coincidental43 and of OV6 and CK14/CK19 as similar.41 We found that Thy-1+ cells demonstrate double immuno-positivity for CK14 and CK19 as well as CK14 and OV6 suggesting co-expression of these markers on Thy-1+ cells. Co-expression of CK14, CK19 and positivity for OV6 of Thy-1+ cells might indicate a similarity of human progenitor cells to the progenitor cell development in rodents as it was postulated in human fetal liver.14 Our findings of Thy-1+ cells isolated from adult human liver tissue and expressing a variety of progenitor, biliary and hepatocytic markers might correspond to a more accepted understanding that human liver progenitor cells do not represent a single, defined cell type but rather a range of cells of intermediate hepatobiliary phenotype.7
Thy-1+ progenitor cells may differentiate into hepatic cells and repopulate a host liver
Activation and expansion of liver progenitor cells is dependent on a specific microenvironment involving multiple growth modulators, which are likely to be mimicked in a variety of in vitro models.44 Using a protocol for hepatocyte expansion27 we could induce proliferation of Thy-1+ cells in vitro, which was paralleled by differentiation into hepatocytic but also biliary lineage without favouring one of each other. During culture expression of progenitor markers (chromogranin A, nestin, CK14) decreased and both hepatocytic and biliary markers increased, which may resemble a process of liver development. Furthermore, it is suggested that progenitor cells losing CK14 expression become committed to either hepatocyte or biliary epithelial cell lineages.38 Interestingly, CD34+ and c-kit+ cells isolated from adult human liver were shown to differentiate only into biliary epithelial cell lineage based on a biliary cell growth media,13 which did have an impact on the differentiation of Thy-1+ cells in this study (data not shown).
To further assess the participation of transplanted Thy-1+ cells in liver regeneration a mouse model for cell transplantation after partial hepatectomy was used.25 26 Interestingly, freshly isolated Thy-1+ cells showed a weak or negative albumin expression. After transplantation the cells showed clear albumin expression even though it was not detectable during culture. Studies for in vitro differentiation of progenitor cells may not14 19 45 or do lead to expression of functional genes such as albumin19 45 depending on specific culture conditions. After transplantation and the under influence of a variety of factors in vivo, differentiation of progenitors is seen from low or none to albumin expressing cells even in a xenogenic model.19 45 However the repopulation efficiency was quite low (about 3%) to replace cells and reverse a pathological condition,46 but was within the range of published data for liver cell engraftment in xenogenic models. One reason might be murine liver failing the proper microenvironment for the human cells and therefore no cell cluster formation was seen.
Human bipotential liver progenitor cells would be an invaluable therapeutic tool to induce liver regeneration or to support liver function by cell transplantation, because of the increasing disparity between the increase in end stage liver diseases and the shortage of suitable donor organs. This study might suggest a strategy of isolating progenitor cells from resected, slightly diseased, liver tissue followed by an autogenic re-transplantation of, for example, genetically altered cells to enrich the regenerative potential. Future research on human adult liver progenitor cells should address the question of separation of different subpopulations, their specific capabilities for expansion in vitro and their applicability for transplantation.
Acknowledgments
We gratefully acknowledge the technical assistance of Anja Gräbe, Center of Liver Cell Research, University of Regensburg, Germany.
REFERENCES
Footnotes
Funding: This work was supported by a fellowship of the medical department, University of Regensburg (ReForM-C program).
Competing interests: None.
Ethics approval: Use of human tissue samples in this study was approved by the ethics committee of the University of Regensburg. All housing and experimental procedures using animals were in accordance with the most recent German legislation concerning animal protection.