Article Text

Abstract
Background and aim Chronic cholangiopathies have limited therapeutic options and represent an important indication for liver transplantation. Curcumin, the yellow pigment of the spice turmeric, has pleiotropic actions and attenuates hepatic damage in animal models of chemically-induced liver injury. Whether curcumin has beneficial effects in cholangiopathies is unknown.
Methods Potential anticholestatic, anti-inflammatory and antifibrotic mechanisms of curcumin were explored in vivo in Mdr2−/− mice as a murine model of chronic cholangiopathy; as well as in vitro in a cholangiocyte cell line (HuCCT1) and portal myofibroblasts (MFBs) isolated from Mdr2−/− mice.
Results Liver damage, cholestasis and fibrosis were reduced in Mdr2−/− mice after curcumin feeding. Moreover, curcumin inhibited cholangiocyte proliferation and expression of activation marker vascular cell adhesion molecule-1 in Mdr2−/− mice. Curcumin—similar to PPARγ synthetic agonist troglitazone—directly inhibited TNF-α-induced inflammatory activation of cholangiocytes in vitro, whereas these beneficial effects of curcumin were largely blocked by a PPARγ synthetic antagonist. In addition, curcumin blocked proliferation and activation of portal MFBs by inhibiting ERK1/2 phosphorylation, thus contributing to reduced fibrogenesis.
Conclusions These results show that curcumin may have multiple targets in liver including activation of PPARγ in cholangiocytes and inhibition of ERK1/2 signalling in MFBs, thereby modulating several central cellular events in a mouse model of cholangiopathy. Targeting these pathways may be a promising therapeutic approach to cholangiopathies.
- Cholangiopathy
- curcumin
- Mdr2
- portal fibrosis
- sclerosing cholangitis
- liver
Statistics from Altmetric.com
Chronic cholangiopathies such as primary sclerosing cholangitis (PSC) and primary biliary cirrhosis are characterised by progressive inflammation and subsequent development of biliary fibrosis and cirrhosis.1 2 Since the effectiveness of currently available medical therapies to slow the progression of cholangiopathies is limited, there is an urgent need for novel and effective medical treatment strategies. Importantly, cholangiocytes may undergo phenotypic and functional modifications, characterised by production of pro-inflammatory and profibrogenic mediators, thus contributing to propagation and progression of liver diseases.3–5 Therefore, it is attractive to hypothesise that targeting the bile duct inflammatory phenotype may be beneficial in treatment of cholangiopathies.
Curcumin, a non-steroidal yellow pigment found in rhizomes of the perennial herb Curcuma longa, has been used for centuries in Ayurvedic medicine to cure a wide range of gastrointestinal disorders, and is a component of the spice turmeric.6–11 In rodent models of chemically induced liver damage and fibrosis such as carbon tetrachloride and thioacetamide-induced liver cirrhosis, curcumin was shown to have anti-inflammatory, antioxidative and antifibrotic properties.12–15 Moreover, blocking c-Jun N-terminal kinase (JNK) signalling by curcumin was shown to inhibit inflammation-mediated alterations of hepatobiliary transporter gene expression in hepatocytes.16 However, to date no study has addressed the effects of curcumin on bile duct inflammation and liver damage in cholangiopathies. We therefore designed a longitudinal study to explore potential molecular mechanisms of curcumin in multidrug-resistant protein 2 knockout (Mdr2−/−) mice serving as a genetic model of progressive cholangiopathy with biliary fibrosis.17–21
We herein demonstrate that curcumin reduces liver damage, cholangitis and biliary fibrosis in Mdr2−/− mice most likely by acting on multiple targets: inhibiting the inflammatory phenotype of bile duct epithelial cells through PPARγ activation and blocking proliferation/activation of portal myofibroblasts (MFBs) through inhibition of extracellular signal-regulated kinase 1/2 (ERK1/2) signalling.
Materials and methods
Animal experiments
Mdr2−/− mice (FVB/N background) were obtained from Jackson Laboratory (Bar Harbor, ME, USA) and housed with a 12:12 hour light/dark cycle with water and a mouse diet (Harlan Teklad, Madison, WI, USA) ad libitum. Male mice were used exclusively. Experimental protocols were approved by the local Animal Care and Use Committee according to criteria outlined in the Guide for the Care and Use of Laboratory Animals prepared by US National Academy of Sciences (National Institutes of Health publication 86–23, revised 1985).
Feeding protocols
Feeding of control and curcumin-enriched (4% wt/wt) diets in Mdr2−/− mice was performed for 4 and 8 weeks starting at the age of 4 weeks, a time point when the principal features of bile duct disease and biliary fibrosis are already present and show high activity.17 22 Curcumin was kindly gifted by Sabinsa Corporation in form of Curcumin C3 complex (Sabinsa Utah, Payson, UT, USA), which contained 96.24% curcuminoids, of which 72.1% was curcumin, 21.98% demethoxy curcumin and 5.91% bisdemethoxy curcumin. Wild type (WT) mice of the same age received a control diet (Harlan Teklad, Madison, WI, USA).

Curcumin reduces liver damage and cholestasis in Mdr2−/− mice. A. Chow- as well as curcumin-fed wild type mice (WT and WT+Cu) demonstrate normal liver structure (H&E). Representative liver histology of 8-week-old Mdr2−/− mouse shows sclerosing cholangitis. Curcumin feeding for 4 weeks reduces bile duct damage as well as periductal fibrosis in Mdr2−/− mouse (Mdr2−/−+Cu) compared with age-matched littermate. B. Curcumin feeding in WT mice is non-toxic as shown by unchanged serum parameters of liver injury (ALT) and cholestasis (AP and BAs). Compared with WT mice, untreated Mdr2−/− mice show increased serum parameters of liver injury (ALT) and cholestasis (AP and BAs), whereas Mdr2−/−+Cu mice have significantly reduced serum biochemical parameters of liver damage and cholestasis. Values are means±SD from five animals per group. *p<0.05 Mdr2−/− vs WT, #p<0.05 Mdr2−/−+Cu vs Mdr2−/−. ALT, alanine aminotransferase; AP, alkaline phosphatase; BAs, bile acids; bd, bile duct; pv, portal vein.
Food intake, body weight/liver weight measurement
Food intake was controlled at days 1, 3 and 7 during the first week, and further monitored once a week, whereas gain in body weight (BW) was monitored daily. Liver weight (LW) was compared to the BW and the percentage ratio was calculated (LW/BW).
Routine serum biochemistry
Blood was collected at harvesting and centrifuged for 15 min at 4500 rpm. Serum was stored at −80°C until analysis. Assays for alanine aminotransferase and alkaline phosphatase were routinely measured, whereas serum bile acid (BA) concentration was determined by using Bile Acid Kit (Ecoline S+ from DiaSys Diagnostic Systems GmbH, Holzheim, Germany) in a Hitachi 917 analyser (Boehringer Mannheim, Mannheim, Germany).
Liver histology
For conventional light microscopy, livers were fixed in 4% neutral buffered formaldehyde solution for 24 h and embedded in paraffin. Sections were cut 2 μm thick and stained with haematoxylin and eosin (H&E) or Sirius red.
Determination of hepatic hydroxyproline content
To quantify liver fibrosis, hepatic hydroxyproline (HP) was measured from standardised liver lobe as described previously.23
Immunohistochemistry
Detection of proliferation marker Ki-67, cholangiocyte cytoskeleton marker keratin 19 (K19), vascular cell adhesion molecule-1 (VCAM-1), neutrophil granulocyte marker CD-11b and lymphocyte markers CD4 and CD8 was performed as described.17 18
Western blotting for hepatic K19 and hepatocellular transporters Ntcp, Bsep, Mrp2 and Mrp3
Total proteins from liver homogenates were isolated as described24 and protein concentrations were determined using a Bradford kit (BioRad, Richmond, CA, USA). Western blotting for K19 was performed as previously described.18 For hepatocellular transporters, preparation of liver membranes, detection of immunocomplexes as well as densitometric quantification of band intensities, was performed as described.25 26
Measurement of bile flow and composition
Bile flow and output of biliary phospholipids (PLs), BAs, cholesterol and glutathione were determined as previously described.18 Experimental feeding was initiated in 4-week-old Mdr2−/− and WT mice. After 7 days of feeding either a control or a curcumin-enriched diet, mice were anaesthetised (10 mg of avertin intraperitoneally), abdominal cavity was opened, common bile duct was ligated and gallbladder was cannulated. After a 10 min equilibration period, bile was collected in pre-weighed tubes for 30 min. Bile flow was determined gravimetrically and normalised to LW. Samples were kept frozen at −20°C until analysis.
Messenger RNA analysis and Polymerase Chain Reaction (PCR)
RNA isolation, complementary DNA synthesis, and real-time PCR were performed as described previously.27
Isolation, culture and experiments with portal myofibroblasts
Portal MFBs were isolated as described.28 Briefly, the biliary tree was isolated from 12-week-old Mdr2−/− mice by collagenase and pronase digestion. Portal tract residues were allowed to adhere in Petri dishes and cultured in DMEM medium (PAA Laboratories GmbH, Pasching, Austria) supplemented with 10% foetal calf serum (FCS) and 1% penicillin/streptomycin. After 2–3 days an extensive outgrowth and adherence of portal MFBs was observed around biliary structures. Purification of portal MFBs from other cells was obtained through serial passages and cell type analysis was made by immunofluorescence staining for hepatocellular cytoskeleton marker K8/18, activated MFB marker α-SMA and nuclear marker DAPI. The majority of isolated cell population was α-SMA-positive and K8/18-negative, demonstrating sufficient purity (data not shown). Cells from passage 8–20 were used for experiments. Cell viability was tested by trypan blue dye exclusion after 24 h of curcumin incubation. At concentrations of 10 and 20 μm curcumin did not induce MFB death, while 30 μm was toxic; therefore, 10 and 20 μm concentrations were subsequently used for in vitro experiments. In proliferation studies, portal MFBs were stained for proliferation marker Ki-67. Nuclear and cytoplasmic proteins were isolated by using a Pierce kit (NE-PER Nuclear and Cytoplasmic Extraction Reagents, Rockford, IL, USA). Western blotting for NF-κB and pERK was performed by using polyclonal rabbit antibodies against NF-κB (dilution 1:2000) (NeoMarkers, Fremont, CA, USA) and pERK1/2 (dilution 1:1000) (Cell Signalling Technology, Inc, Danvers, MA, USA). Specific binding was detected by secondary anti-rabbit antibody (dilution 1:3000) (Cell Signalling Technology, Inc, Danvers, MA, USA). Equal protein loading was confirmed by Coomassie blue staining of gels and Ponceau S staining of membranes.

Hepatic fibrosis is reduced in Mdr2−/− mice by curcumin. A. Sirius red staining did not differ between chow-fed and curcumin-fed wild type mice (WT and WT+Cu respectively). Representative Sirius red staining shows pronounced fibrosis with collagen deposition (red) around bile ducts in Mdr2−/− mouse (arrowheads) compared with WT mouse. Four-week curcumin-treated Mdr2−/− mouse (Mdr2−/−+Cu) demonstrates reduced collagen deposition compared with age matched untreated littermate (original magnification ×20). B. Hepatic hydroxyproline (HP) content is significantly reduced by curcumin in Mdr2−/− mice. C. Curcumin feeding significantly inhibits hepatic Col1α2 mRNA level in Mdr2−/− mice. Expression levels are normalised to 18s gene expression and levels of WT are expressed as 1. Values are presented as means±SD from 4–5 mice per group. *p<0.05 Mdr2−/− vs WT, #p<0.05 Mdr2−/−+Cu vs Mdr2−/−, bd, bile duct; pv, portal vein.
Cholangiocyte culture experiments
Cholangiocarcinoma cell line (HuCCT1) was obtained from Japan Health Sciences Foundation Resources Bank, Osaka, Japan. Cells were incubated in RPMI culture medium (PAA Laboratories GmbH, Pasching, Austria) with 10% FCS and used for experiments at 80% confluence. TNF-α and troglitazone were from Sigma–Aldrich Chemie, GmbH, Steinheim, Germany. Bisphenol A diglycidyl ether (BADGE) was obtained from Sigma–Aldrich, Vienna, Austria. Cytotoxicity of curcumin was tested by trypan blue dye exclusion method.
Statistical analysis
Results were evaluated using SPSS V.14.0. Statistical analysis was performed using one-way analysis of variance test followed by Mann–Whitney. Data are reported as means of five animals per group (unless otherwise noted) ±SD. A p value ≤0.05 was considered significant.
Results
Curcumin significantly reduces liver damage and cholestasis in Mdr2−/− mice
We first addressed whether curcumin-enriched diet was well tolerated by Mdr2+/+ (WT) and Mdr2−/− mice. Survival rate (100%) and food consumption (average: 4–5 g/day) did not differ between study groups. LW/BW ratio was increased in 8-week-old Mdr2−/− mice compared with age-matched WT mice and remained unchanged after curcumin feeding (data not shown). Curcumin feeding reduced sclerosing cholangitis, ductular proliferation (figure 1A), serum liver enzymes (alanine aminotransferase and alkaline phosphatase) and BA levels in Mdr2−/− mice (figure 1B) after 4 weeks of feeding. Compared with 4-week-feeding, prolonged curcumin feeding for 8 weeks did not result in a further reduction of serum liver enzymes (figure 3B,C). Taken together, these data demonstrated beneficial effects of curcumin on cholestatic phenotype in Mdr2−/− mice.

Curcumin feeding for 8 weeks reduces liver damage and fibrosis in Mdr2−/− mice. A. Schematic illustration of feeding protocols in Mdr2−/− mice. Experimental feeding in Mdr2−/− mice was initiated at the age of 4 weeks, when the ductular proliferation and fibrosis showed high activity and lasted for 4 or 8 weeks. Feeding of age matched wild type mice followed the same experimental protocol. B. Serum liver enzymes and hepatic hydroxyproline were measured in Mdr2−/− mice to quantify liver damage and fibrosis after 8 weeks of curcumin feeding. Curcumin reduces serum liver enzymes ALT and AP as well as fibrosis in Mdr2−/− mice after 8 weeks of feeding. Values are presented as means±SD from four animals per group. #p<0.05 Mdr2−/−+Cu vs Mdr2−/−. ALT, alanine aminotransferase; AP, alkaline phosphatase; HP, hydroxyproline.
Hepatic fibrosis in Mdr2−/− mice is significantly reduced by curcumin
Biliary fibrosis, determined by Sirius red staining, measurement of hepatic HP levels and Col1α2 mRNA expression, was reduced in Mdr2−/− mice after 4 weeks of curcumin feeding (figure 2A–C). With age untreated Mdr2−/− mice showed progression of fibrosis as determined by increased hepatic HP levels (243±45.8 at 12 weeks vs 166±3.1 μg/g LW at 8 weeks). Prolonged curcumin feeding (figure 3D) resulted in a more pronounced relative reduction of HP levels (42.7% after 8-week-feeding vs 21.9% reduction after 4-week-feeding), but did not further decrease HP levels compared with 4-week-feeding (139±12 vs 129.6±15 μg/g LW, not significant). Taken together, these findings suggest that curcumin feeding had no effect on resolution, but rather reduced progression of hepatic fibrosis in Mdr2−/− mice via inhibition of collagen production.

Transport of biliary constituents and bile acid synthesis are not affected by curcumin in Mdr2−/− mice. A. Liver membranes for Western blotting were isolated from wild type (WT) and Mdr2−/− mice fed either a control or a curcumin-enriched diet for 4 weeks. Curcumin feeding does not influence levels of key hepatocellular transporters in Mdr2−/− mice (Mdr2−/−+Cu). Densitometry data are expressed as fold change relative to WT group. Values are means from three to four animals per group. β-actin protein levels were used as a protein loading control. *p<0.05 Mdr2−/− and Mdr2−/−+Cu vs WT. B. Cyp7α1 gene expression does not differ between WT, WT+Cu, Mdr2−/− and Mdr2−/−+Cu groups. Expression levels are normalised to 18s gene expression and levels of WT are expressed as 1. Values are presented as means±SD from five animals per group. Ntcp, Na+/taurocholate cotransporting polypeptide; Mrp2, multidrug resistance associated protein 2; Mrp3, multidrug resistance associated protein 3; Bsep, bile salt export pump; Cyp7α1, cholesterol 7 α-hydroxylase.
Curcumin increases bile flow and biliary glutathione output in Mdr2−/− mice
Since liver disease in Mdr2−/− mice primarily results from toxic bile composition due to increased concentrations of non-micellar bound BAs,29 we next addressed whether curcumin modified bile formation and output of main biliary constituents. To distinguish between direct effects on bile flow and liver healing properties of curcumin, 4-week-old WT and Mdr2−/− mice were fed short-term (7 days) either a control or a curcumin-enriched diet. In WT mice, curcumin increased bile flow and biliary glutathione output, but did not modify biliary BA, PL and cholesterol output (table 1). Compared with WT mice, biliary PL and cholesterol output was dramatically reduced in Mdr2−/− mice and remained unchanged after curcumin feeding. In addition, curcumin increased bile flow in Mdr2−/− mice and restored altered biliary glutathione output (table 1). However, total hepatic glutathione content did not differ between the groups (data not shown), suggesting that curcumin had no impact on hepatic glutathione synthesis. Collectively, these data show that curcumin increased bile flow and biliary glutathione output, but did not modify bile toxicity as reflected by unchanged biliary PL, cholesterol and BA output.
Curcumin effects on bile flow and composition in wild type and Mdr2−/− mice
Therapeutic effects of curcumin in Mdr2−/− mice are neither mediated by changes of bile acid transport nor synthesis
Reduction of serum BA levels in curcumin-treated Mdr2−/− mice may reflect alterations of BA transport and synthesis. Neither expression of the main sinusoidal BA importer (Ntcp) nor canalicular BA/anion exporters (Bsep, Mrp2, Mrp3) were altered by curcumin feeding in Mdr2−/− mice (figure 4A). Expression of Cholesterol 7 α-hydroxylase (Cyp7α1), the rate limiting enzyme for BA synthesis also did not differ significantly between WT, Mdr2−/− and curcumin-treated Mdr2−/− mice (figure 4B). Taken together, these data indicate that curcumin had no effect on hepatic BA homeostasis in Mdr2−/− mice.
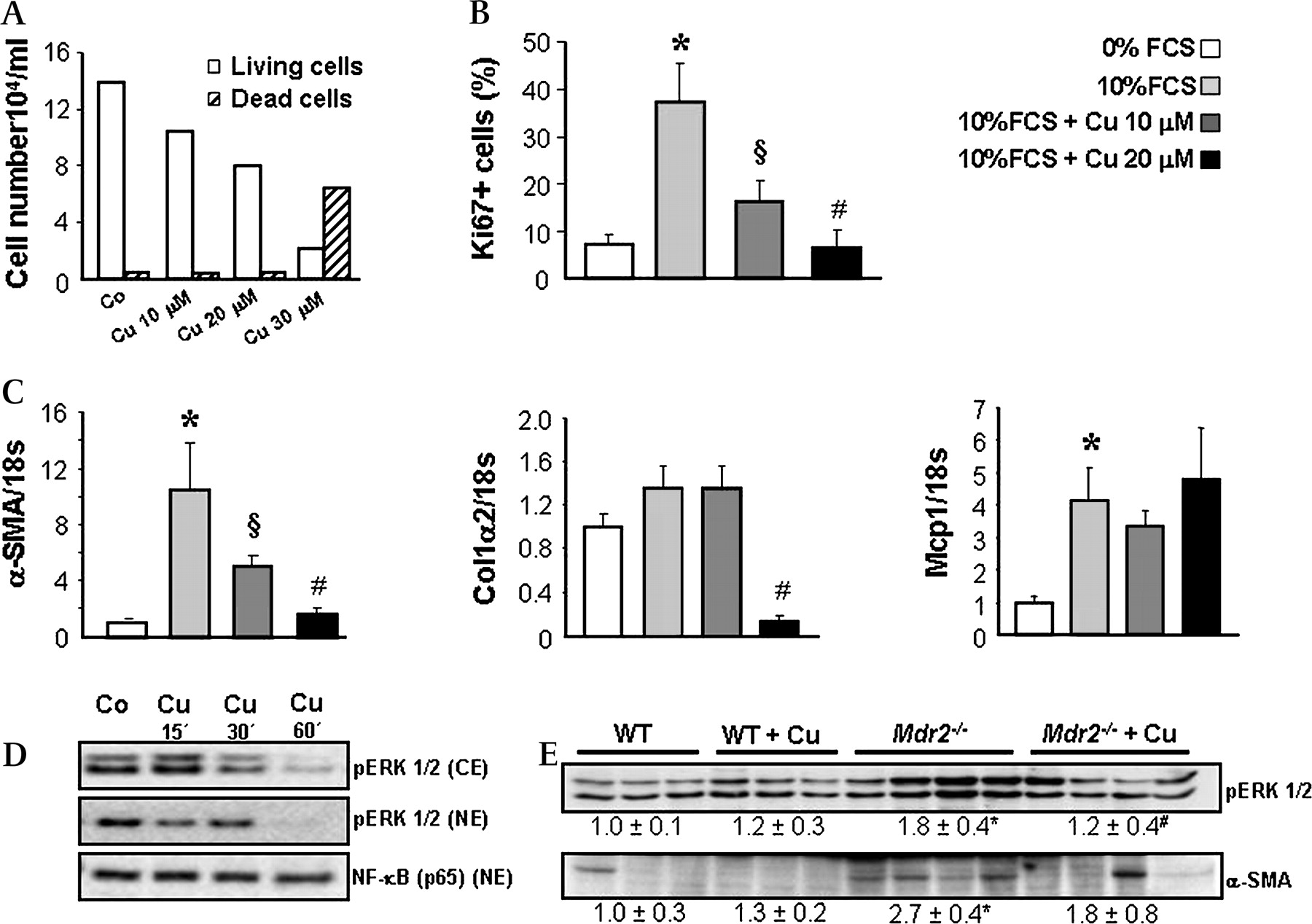
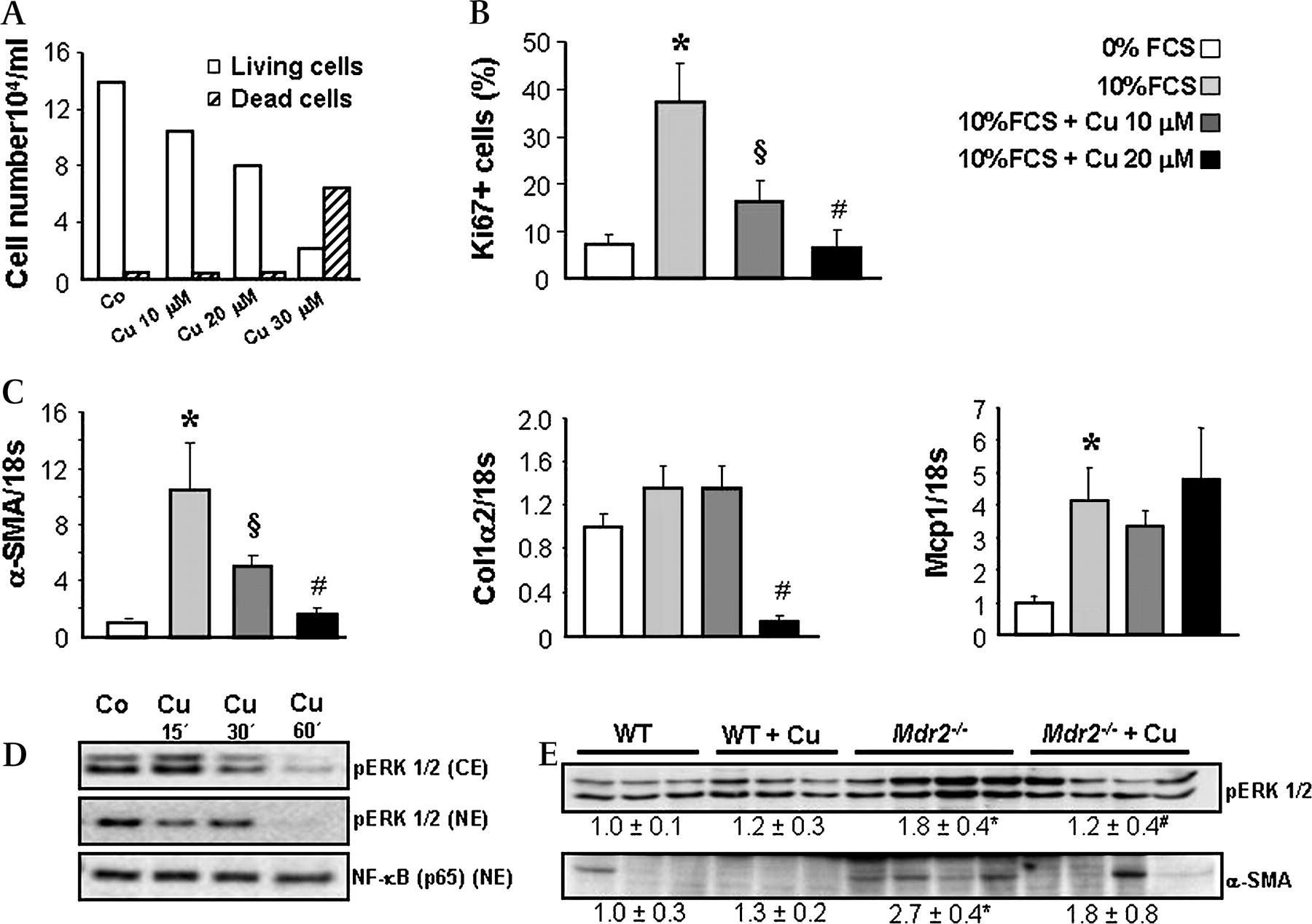
Curcumin inhibits proliferation and activation of portal myofibroblasts. A. Portal myofibroblasts (MFBs) were isolated from Mdr2−/− mice as described in Materials and methods and cytotoxic effect of curcumin was tested after 24 h incubation at concentrations of 10, 20 and 30 μm by using trypan blue dye exclusion method. Absolute number of living as well as dead cells was counted and calculated per 1 ml of cell suspension. At concentrations of 10 and 20 μm curcumin does not induce MFB cell death, whereas 30 μm is toxic. B. Influence of curcumin on MFB proliferation was assessed by immunohistochemical staining for Ki-67. Cells were first synchronised in 0% FCS containing medium and further grown on sterile cover slips using 0% FCS (to reduce proliferation), 10% FCS (to induce proliferation) and curcumin at concentrations of 10 and 20 μm in 10% FCS medium for 24 h. Number of Ki-67-positive nuclei was counted in 10 randomly chosen power fields in triplicate for each condition and the mean values of Ki-67-positive versus total cell number ratio (%) was calculated. Compared with 0% FCS condition 10% FCS stimulates MFBs proliferation, while incubation with curcumin (10 and 20 μm) reduces 10% FCS-induced proliferation of MFBs in a concentration-dependent manner. C. Curcumin effects on profibrogenic gene expression were studied in 10% FCS-activated MFBs after 24 h incubation (as described in B). Curcumin inhibits Col1α2 and α-SMA gene expression, whereas Mcp-1 expression remains unchanged. Expression levels are normalised to 18s gene expression and levels of wild type (WT) are expressed as 1. B and C: Data are presented as means±SD. *p<0.005 10% FCS vs 0% FCS, §p<0.005 10% FCS+Cu 10 μm vs 10% FCS, #p<0.005 10% FCS+Cu 20 μm vs 10% FCS. D. Cytoplasmic and nuclear proteins were isolated from MFBs after incubation with 10% FCS and 20 μm curcumin and analysed for pERK1/2 and NF-kB (p65) by Western blotting. Curcumin does not modify nuclear translocation of NF-κB, but inhibits ERK phosphorylation in portal MFBs. Representative data from two independent experiments performed in duplicate for each condition are shown. E. Proteins were isolated from livers of wild type (WT) and Mdr2−/− mice fed either a control or a curcumin-enriched diet for 4 weeks and analysed for pERK and α-SMA by Western blotting. Curcumin significantly reduces cytoplasmic pERK1/2 in livers of Mdr2−/− mice. Densitometry data are expressed as fold change relative to WT mice. Values are presented as means±SD. *p<0.05 Mdr2−/− vs WT, #p=0.05 Mdr2−/−+Cu vs Mdr2−/−. Co, control; Cu, curcumin; FCS, Foetal Calf Serum; Col1α2, collagen, type I, α 2; α-SMA, α smooth muscle actin; Mcp-1, monocyte chemotactic protein 1; CE, cytoplasmic extracts; NE, nuclear extracts.
Curcumin reduces proliferation of activated portal myofibroblasts and inhibits ERK signalling
Since activated portal MFBs represent an important source for the extracellular matrix in Mdr2−/− mice, we next studied direct antifibrotic mechanisms of curcumin in portal MFBs. Since isolated MFBs as well as hepatic stellate cells (HSCs) demonstrate high proliferation rate and activity upon contact with the plastic, we used 0% FCS to inhibit MFB activation and synchronise the cells and 10% FCS to stimulate MFB activation. Compared with 0% FCS condition, 10% FCS stimulated proliferation as well as expression of fibrogenic markers in portal MFBs (figure 5B,C), while curcumin at 10 and 20 μm, two doses shown to be nontoxic (figure 5A), reduced 10% FCS-induced proliferation of portal MFBs (figure 5B). We next studied PPARγ (NR1C3), NF-κB and extracellular signal-regulated kinase (ERK) signalling as mechanisms shown to mediate proliferation and activation of fibrogenic cells.30–34 Activation of MFB-like cells is associated with diminished PPARγ expression and increased ability to modulate the recruitment and activation of inflammatory cells by expression of monocyte chemotactic protein 1 (Mcp-1).34 35 MFBs showed preserved basal PPARγ1 expression, which remained unchanged after curcumin incubation (data not shown). Moreover, expression of Mcp-1, another gene altered by PPARγ ligands in fibrogenic cells,34 remained unchanged in response to curcumin (figure 5C). However, curcumin profoundly inhibited 10% FCS-induced expression of activation markers Col1α2 and α-SMA in MFB cultures (figure 5C). These data led us to focus on PPARγ-independent pathways. Interestingly, cytoplasmic as well as nuclear content of pERK1/2 was dramatically reduced by curcumin (figure 5D) in portal MFBs, whereas nuclear NF-κB (p65) content remained unchanged. Under in vivo conditions, livers of curcumin-treated Mdr2−/− mice showed a reduction of MFB activation marker α-SMA and decreased cytosolic pERK1/2 content compared with untreated littermates (figure 5E). Collectively, these data suggest that the antiproliferative effects of curcumin on MFBs could be mediated via ERK1/2 inhibition.
Curcumin attenuates reactive phenotype of bile duct epithelial cells by PPARγ activation
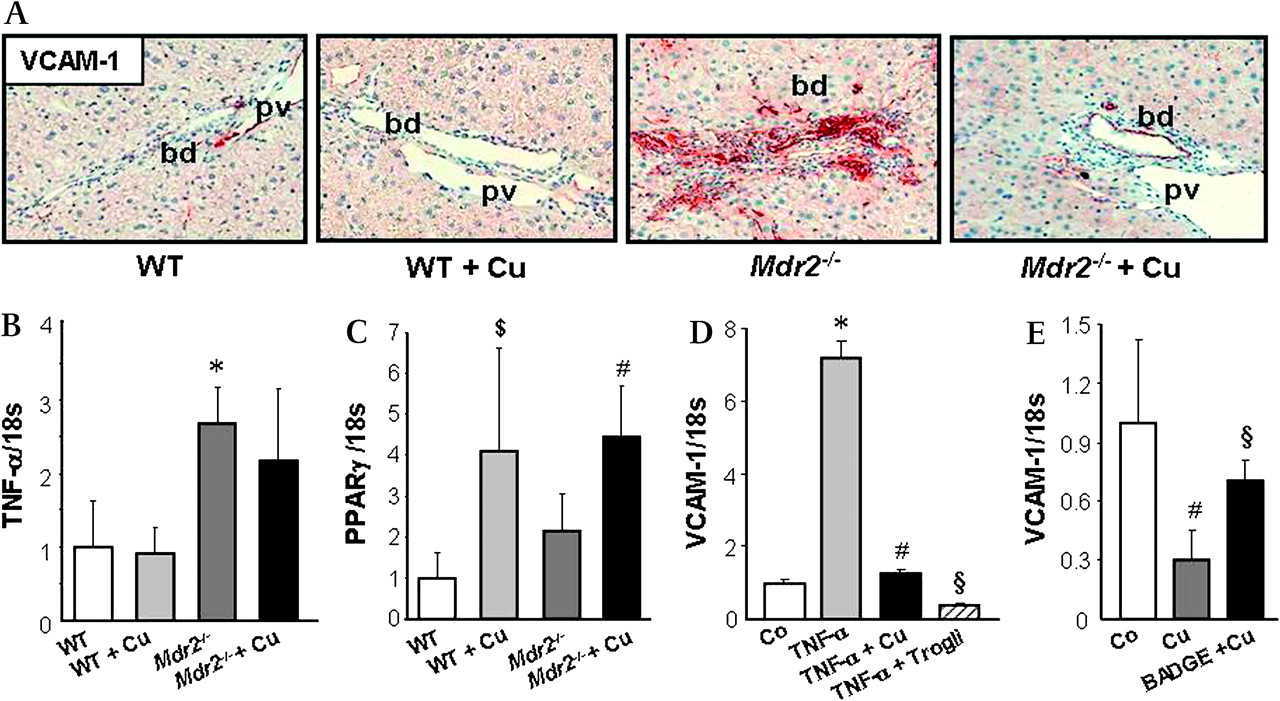
Cholangiopathy in Mdr2−/− mice is characterised by increased proliferation of cholangiocytes and overexpression of pro-inflammatory cytokines and adhesion molecules (eg, VCAM-1) described as reactive phenotype.5 17 Curcumin-treated Mdr2−/− mice showed decreased staining for K19 as well as significantly decreased hepatic K19 content (figure 6A,B) compared with untreated Mdr2−/− mice. In addition, bile ducts of untreated Mdr2−/− mice were highly positive for VCAM-1 expression compared with WT mice, whereas curcumin feeding reduced VCAM-1 staining in bile ducts of Mdr2−/− mice (figure 7A). VCAM-1 gene expression can be stimulated by pro-inflammatory cytokines such as tumour necrosis factor α (TNF-α).36 37 TNF-α expression was significantly increased in Mdr2−/− mice but remained unchanged after curcumin treatment (figure 7B). In contrast, PPARγ expression increased in both curcumin-treated WT and Mdr2−/− mice (figure 7C), suggesting, that inhibition of production in bile duct lining cells by curcumin could be PPARγ-mediated. In line with our in vivo findings, incubation of the cholangiocyte cell line HuCCT1 (expressing functionally active PPARγ) with TNF-α induced VCAM-1 expression, which was blocked by curcumin as well as specific PPARγ agonist troglitazone (figure 7D). Moreover, curcumin mediated inhibition of VCAM-1 expression in cholangiocytes was largely restored by pre-incubation and co-incubation with the PPARγ synthetic antagonist BADGE (figure 7E). Together, these findings suggest, that curcumin reduced bile duct damage by inhibiting cholangiocyte activation via PPARγ signalling.

Curcumin feeding significantly reduces bile duct proliferation in Mdr2−/− mice. A. Representative immunohistochemical staining for cholangiocyte cytoskeleton marker K19 (red) shows increased ductular proliferation in Mdr2−/− mouse compared with wild type (WT). Immunochemical staining for K19 does not differ between chow-fed (WT) and curcumin-fed wild type (WT+Cu) mouse, whereas 4-week curcumin-treated Mdr2−/− mouse (Mdr2−/−+Cu) demonstrates reduced K19 staining. B. Total protein was isolated from livers and analysed by Western blotting for K19 content. Mdr2−/−+Cu mice show significantly decreased K19 protein levels compared with control diet-fed age-matched littermates. Densitometry data are expressed as fold change relative to WT mice. Values are presented as means±SD. *p<0.05 Mdr2−/− vs WT, #p=0.05 Mdr2−/−+Cu vs Mdr2−/−.
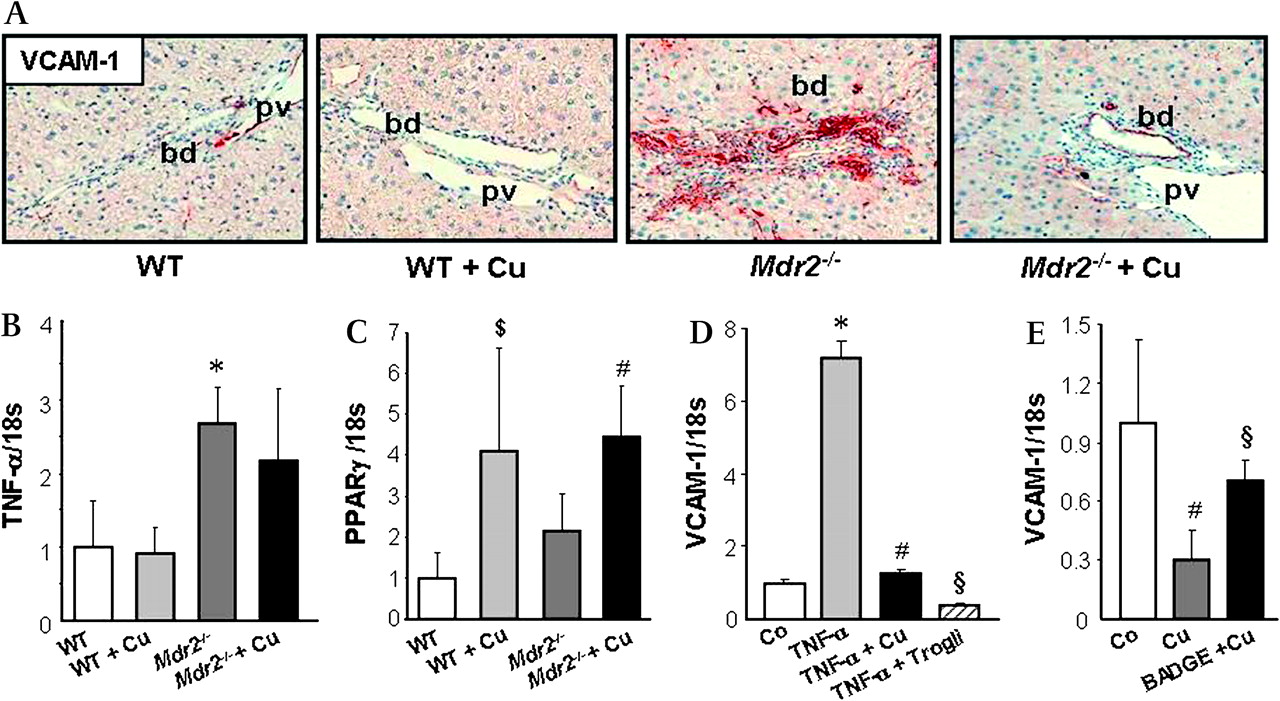
Curcumin decreases Vascular Cell Adhesion Molecule 1 (VCAM-1) production in bile duct epithelial cells. A. Representative immunohistochemical staining for VCAM-1 (red) in bile ducts of a chow-fed and curcumin-fed wild type (WT and WT+Cu respectively), untreated Mdr2−/− and 4-week curcumin-treated Mdr2−/− mouse (Mdr2−/−+Cu) is shown. Bile ducts from chow-fed and curcumin-fed WT liver stain negative for VCAM-1, while bile ducts in untreated Mdr2−/− liver demonstrate pronounced VCAM-1 staining. Curcumin feeding reduces VCAM-1 staining in bile ducts. bd, bile duct; pv, portal vein. B. Compared with WT mice both untreated and curcumin-treated Mdr2−/− mice demonstrate increased expression of pro-inflammatory cytokine TNF-α. C. Baseline levels of PPARγ gene expression do not differ between untreated Mdr2−/− mice and wild type mice (WT); both curcumin-treated WT and Mdr2−/− mice (Mdr2−/−+Cu) show induced PPARγ expression. B and C: Expression levels are normalised to 18s gene expression and levels of WT are expressed as 1. Values are means±SD of five mice per group. *p<0.05 Mdr2−/− vs WT, #p<0.05 Mdr2−/−+Cu vs WT and Mdr2−/−, $p<0.05 WT+Cu vs WT. D. TNF-α-stimulated VCAM-1 expression in cholangiocytes is blocked by curcumin. HuCCT1 cells were incubated either with TNF-α alone (to induce VCAM-1 expression) or with simultaneous TNF-α (10 ng/ml) and curcumin (10 μm) or a PPARγ specific agonist troglitazone (100 μm) for 24 h. Curcumin as well as troglitazone completely inhibit TNF-α induced VCAM-1 expression in the cholangiocyte cell line HuCCT1. E. Inhibition of VCAM-1 expression in HuCCT1 cholangiocytes by curcumin is partially restored by PPARγ synthetic antagonist BADGE. HuCCT1 cells were pre-incubated with BADGE (50 μm) for 12 h and further incubated either with curcumin alone (10 μm) or simultaneous curcumin and BADGE for 24 h. D and E: Representative experiments performed in triplicate for each condition are presented. Values are means±SD. D: *p<0.005 TNF-α vs Co, #p<0.005 TNF-α+Cu vs TNF-α, §p<0.005 TNF-α+Trogli vs TNF-α, E: #p<0.05 Cu vs Co, §p<0.05 BADGE+Cu vs Cu. Co, control; Cu: curcumin; Trogli, troglitazone.
Curcumin significantly reduces recruitment of inflammatory cells as well as hepatocellular damage and proliferation in Mdr2−/− mice
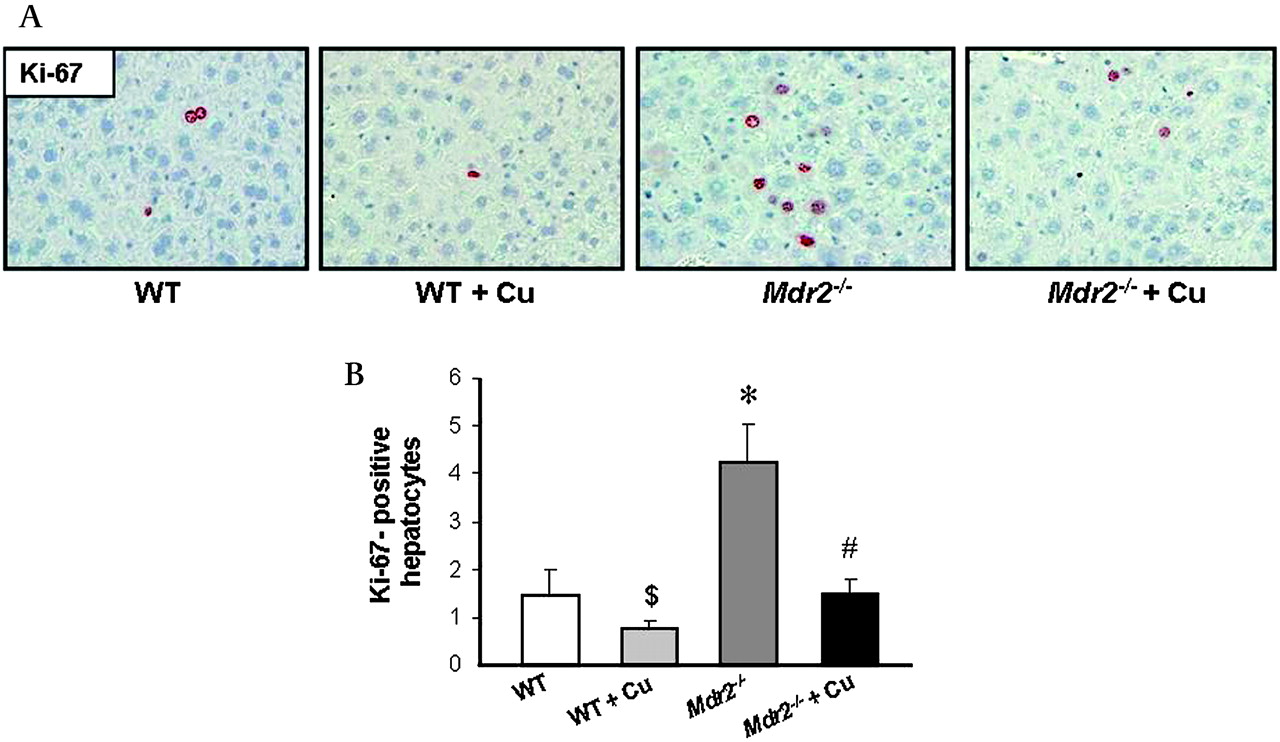
Expression of adhesion molecules and inflammatory cytokines by bile duct cells and activated MFBs stimulates an inflammatory response in the portal tract of Mdr2−/− mice as shown by increased CD11b levels (figure 8A). Curcumin feeding in Mdr2−/− mice resulted in pronounced reduction of CD11b staining (figure 8A,B). In addition, compared with WT mice Mdr2−/− mice demonstrated increased staining of lymphocyte marker CD4 in portal tracts, which was reduced by curcumin (figure 9A). However, CD8 positive cells were not detectable in Mdr2−/− mice at this time point independent of treatment condition (figure 9B). Interestingly, expression of TNF-α, IL-1β and TGF-β did not differ between curcumin-treated and untreated Mdr2−/− mice (figure 7B and data not shown), suggesting that inhibition of VCAM-1 production by cholangiocytes was sufficient to reduce infiltration by neutrophil granulocytes. Since hepatocellular proliferation is a consequence of liver damage, we next studied hepatocellular proliferation by immunohistochemical staining for the proliferation marker Ki-67. Mdr2−/− mice showed increased amount of Ki-67-positive hepatocytes compared with WT, while curcumin reduced Ki-67-staining (figure 10A, B).

Recruitment of neutrophil granulocytes is reduced in portal fields of Mdr2−/− mice after curcumin feeding. A. Representative immunohistochemical staining shows reduction of CD11b-positive cells in portal fields of 4-week curcumin-treated Mdr2−/− mice (Mdr2−/−+Cu) compared with untreated Mdr2−/− mice. CD11b staining did not differ significantly between WT and WT+Cu groups. B. Portal accumulation of neutrophil granulocytes was assessed by counting the number of cells in 20 portal fields per mouse in 4–5 mice per group. Curcumin-treated Mdr2−/− mice show significant reduction of CD11b-positive cells compared with age matched littermates fed a control diet. Values are means±SD from four animals per group. *p<0.05 Mdr2−/− vs WT, #p<0.005 Mdr2−/−+Cu vs Mdr2−/−.
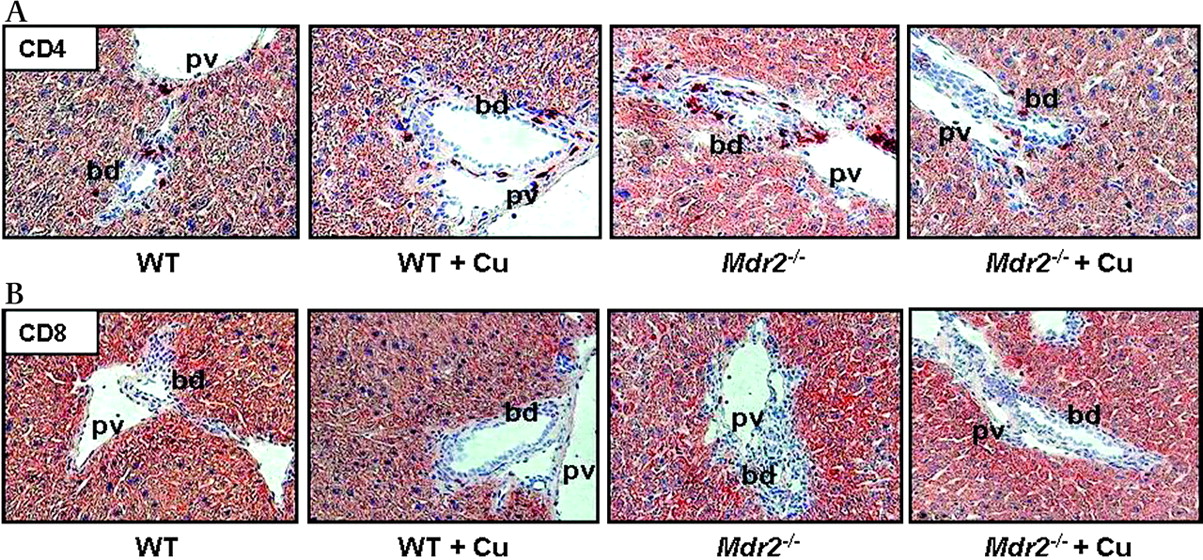
Curcumin reduces recruitment of lymphocytes to the portal fields of Mdr2−/− mice. A. Representative staining for lymphocyte marker CD4 demonstrates increased accumulation of CD4 positive cells in the portal field of Mdr2−/− compared with wild type (WT) mouse. 4-Week curcumin feeding reduces CD4 positive lymphocyte accumulation in portal fields of Mdr2−/− mouse. No pronounced differences are observed in curcumin-fed (WT+Cu) compared with chow-fed WT mice. B. No CD8 positive cells are detected in the portal fields of WT and Mdr2−/− mice independent of treatment. bd, bile duct; pv, portal vein.

Curcumin significantly reduces hepatocellular proliferation in Mdr2−/− mice. A. Immunohistochemical staining for proliferation marker Ki-67 (red) in a wild type (WT), 4-week curcumin-fed wild type mouse (WT+Cu), untreated Mdr2−/− and curcumin-treated Mdr2−/− mouse (Mdr2−/−+Cu) is presented. Compared with WT, WT+Cu mouse shows reduced Ki-67 staining. Mdr2−/− mouse demonstrates increased number of Ki-67-positive hepatocytes compared to WT. In contrast to untreated Mdr2−/−, curcumin-treated Mdr2−/− mouse shows reduced number of Ki-67-positive hepatocytes (original magnification ×20). B. Ki-67-positive hepatocytes were counted in 30 high-power fields per mouse in sections of five animals per group. Curcumin feeding significantly reduces number of proliferating hepatocytes in Mdr2−/− mice. Values are means±SD from five animals per group. *p<0.05 Mdr2−/− vs WT, #p<0.05 Mdr2−/−+Cu vs Mdr2−/−, $p<0.05 WT+Cu vs WT.
Discussion
This study demonstrates that the natural compound curcumin improves bile duct injury and biliary fibrosis in a mouse model of chronic cholangiopathy. We demonstrate that curcumin targets important pathogenic steps in cholangiopathies such as MFB proliferation (via pERK1/2 inhibition) and cholangiocellular VCAM-1 expression (in a PPARγ-dependent fashion). Taken together, curcumin reduces portal inflammation and fibrosis in the Mdr2−/− mouse liver.
Chronic cholangiopathies include a wide spectrum of diseases ranging from autoimmune disorders (PSC, primary biliary cirrhosis, acute/chronic allograft rejection, graft vs host disease) to congenital genetic defects (Alagille syndrome, cystic fibrosis, MDR3 mutations, etc). Long-term beneficial effects of the one established medical therapy, ursodeoxycholic acid, are unclear, and liver transplantation often remains the ultimate therapeutic solution.38–41 Curcumin, an extract from turmeric has been used as traditional medicine for treatment of various gastrointestinal disorders.6–11 42 Interestingly, the early data on curcumin use date back to 1934 when pronounced choleretic effects after intravenous curcumin injection were observed by using the choledochal fistula in rabbits and analysing duodenal juice in humans.43 Shortly thereafter, curcumin was successfully used to treat cholestatic jaundice in patients.10 Acute choleretic effects of curcumin were further confirmed in bile fistula and isolated perfused rat liver models.44 45 Interestingly, curcumin was also shown to reduce gallstone formation in mice after lithogenic diet feeding.46 However, the impact of curcumin on bile composition and chronic cholangiopathies has not yet been explored.
Mdr2−/− mice lacking biliary PL excretion have been characterised as a suitable mouse model for spontaneous development of sclerosing cholangitis.17 Bile duct damage and fibrosis in Mdr2−/− mice start early after birth and evolve over time to a full-blown phenotype at the age of 8 weeks.17 22 To study the potential action of curcumin on progression of liver injury in a mouse model of sclerosing cholangitis we designed 4-week and 8-week feeding protocols (figure 3A) in Mdr2−/− mice starting from the time of weaning (age of 4 weeks). Formation of ‘toxic bile’ due to increased concentrations of non-micellar bound BAs is considered to play a central role in initiation and propagation of bile duct injury in Mdr2−/− mice.29 Since curcumin feeding decreased liver damage and serum parameters of cholestasis in Mdr2−/− mice, we first explored whether curcumin modulates bile flow and composition. Interestingly, the protective action of curcumin in Mdr2−/− mice was not paralleled by changes of main biliary constituents, indicating that its beneficial effects may not be related to changes of bile micellisation. These data are consistent with unchanged expression of BA excretion (Bsep, Mrp2, Mrp3), uptake (Ntcp) or synthesis (Cyp7α1) systems suggesting that curcumin improved serum BA levels indirectly via reduced liver damage in Mdr2−/− mice. In line with early reports in humans10 43 and rats45 curcumin feeding increased bile flow both in WT and Mdr2−/− mice. Moreover, biliary glutathione output was increased in Mdr2−/− as well as WT mice without changes in total hepatic glutathione content showing that glutathione synthesis is not induced by curcumin. Together with unchanged expression of canalicular Mrp2, mediating glutathione export into bile, we speculate that differences in biliary glutathione output may result from reduced biliary damage and glutathione degradation by gamma-glutamyl transpeptidase in Mdr2−/− mice.
Bile ducts in cholangiopathies show increased proliferation and acquisition of new properties such as expression of adhesion molecule VCAM-1, a gene induced in epithelial cells in response to the pro-inflammatory cytokine TNF-α. Curcumin reduced VCAM-1 staining in cholangiocytes of Mdr2−/− mice, while expression of stimulatory pro-inflammatory cytokines TNF-α, as well as IL-1β and TGF-β remained unaffected. In addition, curcumin completely inhibited TNF-α-induced VCAM-1 expression in cholangiocytes in vitro. These findings support the concept that primary inflammation and activation of cholangiocytes is an important event in the pathogenesis of cholangiopathies. Targeting cholangiocyte inflammation might represent an important strategy to cure bile duct diseases despite preserved exposure to the causative damaging factor and pro-inflammatory mediators. Previous reports in other cells have shown that curcumin may act as a PPARγ activator.47–52 Since curcumin-mediated inhibition of VCAM-1 expression in HuCCT1 cells was restored by PPARγ specific antagonist BADGE, it is attractive to speculate that inhibition of both basal and TNF-α-induced VCAM-1 expression in cholangiocytes by curcumin may be at least in part mediated by PPARγ. Previous studies have shown reduction of bile duct proliferation and biliary fibrosis by PPARγ agonists in bile duct-ligated rats.34 53 In line with these studies our data suggest that PPARγ activation in cholangiocytes may inhibit a central pathogenetic event in cholangiopathies, namely acquisition of the cholangiocyte reactive phenotype.3 5
Curcumin reduced liver fibrosis in Mdr2−/− mice after both 4 and 8 weeks of feeding. Although relative reduction of hepatic HP levels was more pronounced after prolonged feeding (42.7% reduction after 8 weeks vs 21.9% reduction after 4 weeks), the absolute HP levels did not differ significantly between 4 and 8 weeks of feeding, suggesting that curcumin inhibits progression of fibrosis rather than stimulating resolution of established fibrosis. A central event towards fibrosis is activation of HSCs and their transformation to MFB-like cells in response to pro-inflammatory cytokines and chemokines.54 55 In biliary fibrosis, the subpopulation of portal MFBs contributes greatly to fibrosis.56 Several studies reported that activation of HSCs and their transformation to MFB-like cells with fibrogenic properties was associated with diminished level and trans-activating ability of PPARγ, while NF-κB activation was increased.31 57–60 Additionally, incubation of activated HSCs with PPARγ agonists was shown to revert the fibrogenic phenotype of these cells by inhibiting collagen production, proliferation, migration and production of the pro-inflammatory chemokine Mcp-1.34 35 60–62 The importance of PPARγ in maintaining the quiescent state of HSCs was further demonstrated by its ectopic expression in activated stellate cells resulting in inhibition of their extracellular matrix producing ability.31 Curcumin reduced proliferation as well as fibrotic activity of portal MFBs isolated from Mdr2−/− mice. In contrast to previous reports, demonstrating that curcumin reduced the activated phenotype of primary HSCs by increasing PPARγ expression and activity, 49 50 52 63 exposure of portal MFBs to curcumin did not induce PPARγ1 gene expression. Moreover, curcumin did not modify Mcp-1 expression, a marker of activated MFB-like cells35 64 known to be inhibited by PPARγ ligands.34 Based on these findings, we suggest that curcumin had a very modest influence on PPARγ activation in MFBs perhaps due to the lack of specific PPARγ co-factors.
Interestingly, curcumin was shown to modulate NF-κB and ERK signalling pathways in HSCs in vitro resulting in reduction of their activation and proliferation.30 49 65 Unchanged nuclear NF-κB content after curcumin incubation is consistent with the lack of PPARγ activation in portal MFBs. However, the reduced proliferation and activation of MFBs was associated with inhibition of ERK1/2 phosphorylation both in vitro and in vivo, suggesting that direct inhibition of ERK signalling could contribute to the reduction of biliary fibrosis by curcumin.
In agreement with the key role of inflammatory cells in the pathogenesis of PSC,66 67 decreased fibrosis in curcumin-treated Mdr2−/− mice correlated with reduced CD11b and CD4 positivity. In addition, direct interaction of HSCs with CD8 positive lymphocytes was suggested to play an important role in liver fibrosis.68 69 However, CD8 positive cells were virtually undetectable in both control and curcumin-treated Mdr2−/− mice at the age of 8 weeks.
Reduced hepatocellular proliferation reflected by decreased Ki-67 staining is in line with improved liver injury in Mdr2−/− mice. Since liver regeneration occurs in response to liver damage, it is tempting to speculate that these antiproliferative effects may result indirectly from pleiotropic effects of curcumin on different liver cell types. Moreover, antiproliferative effects of curcumin have been reported in several cancer cells,63 suggesting that curcumin might also have direct antiproliferative effects in hepatocytes.
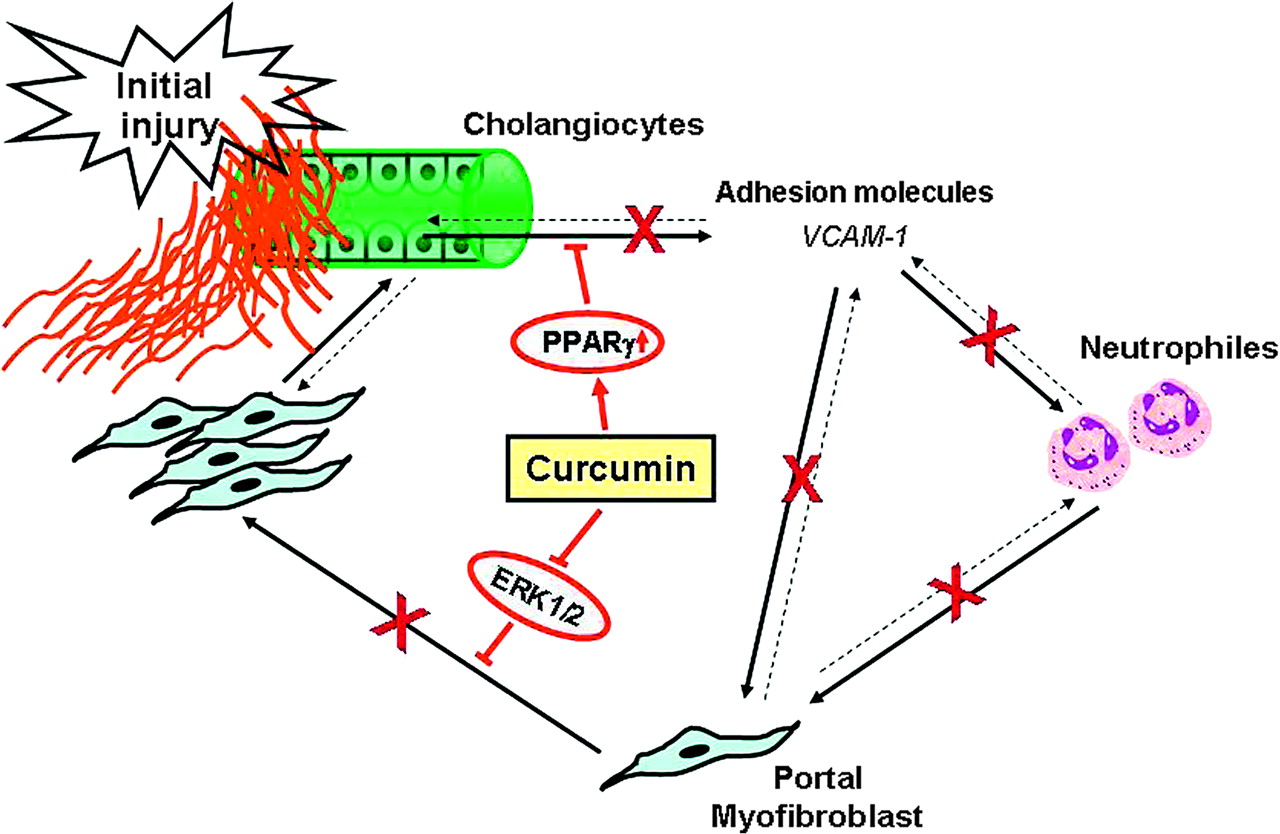
In conclusion, we demonstrate that curcumin is beneficial in a genetic mouse model of cholangiopathy and biliary fibrosis, and propose potential molecular and cellular targets of curcumin in this model (figure 11). Collectively, our findings have important implications for understanding the pathophysiology of cholangiopathies and suggest that combined targeting of cholangiocyte and MFB activation could represent a central strategy to treat or delay the progression of chronic cholangiopathies and liver fibrosis.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Hypothetical mechanisms of curcumin action in Mdr2−/− mice. Our data suggest that curcumin may have pleiotropic actions on several critical steps involved in the pathogenesis of liver damage and biliary fibrosis in Mdr2−/− mice. By activation of PPARγ in cholangiocytes, curcumin may block inflammatory activation and VCAM-1 expression in cholangiocytes, resulting in decreased attraction of neutrophil granulocytes as well as myofibroblasts in the portal fields. In addition, by inhibiting the ERK1/2 signalling curcumin may decrease proliferation of portal myofibroblasts the major fibrogenic cells in Mdr2−/− mice, thus contributing to reduction of liver fibrosis.
Acknowledgments
We gratefully acknowledge Dr W Erwa (Graz) and colleagues for performing biochemical analyses of serum liver tests.
References
Footnotes
A commentary on this article will appear in the May issue
Funding This work was supported by the Austrian Science Foundation, grant numbers P18613-B05, P19118, SFB 3008 (to MT), DK56239 from NIH/NIDDK (to SJK) and by the PhD Program of Medical University of Graz.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Digest