Article Text

Abstract
Despite advances in understanding the roles of adiposity, food intake, GI and adipocyte-related hormones, inflammatory mediators, the gut–brain axis and the hypothalamic nervous system in the pathophysiology of obesity, the effects of different therapeutic interventions on those pathophysiological mechanisms are controversial. There are still no low-cost, safe, effective treatments for obesity and its complications. Currently, bariatric surgical approaches targeting the GI tract are more effective than non-surgical approaches in inducing weight reduction and resolving obesity-related comorbidities. However, current guidelines emphasise non-surgical approaches through lifestyle modification and medications to achieve slow weight loss, which is not usually sustained and may be associated with medication-related side effects. This review analyses current central, peripheral or hormonal targets to treat obesity and addresses challenges and opportunities to develop novel approaches for obesity.
- OBESITY
- BRAIN/GUT INTERACTION
- GASTROINTESTINAL HORMONES
Statistics from Altmetric.com
Introduction
The search for effective treatments for obesity has led to a greater understanding of adiposity, GI and adipocyte-related hormones, inflammatory mediators, the gut–brain axis and the hypothalamic nervous system involved in appetite regulation (figure 1). The effects of different interventions on the pathophysiological mechanisms of obesity are the subject of ongoing research. Understanding the pathophysiological mechanisms of obesity provides an opportunity to develop novel approaches to treat, at relatively low cost and enhanced safety, the ever-expanding population of obese people.
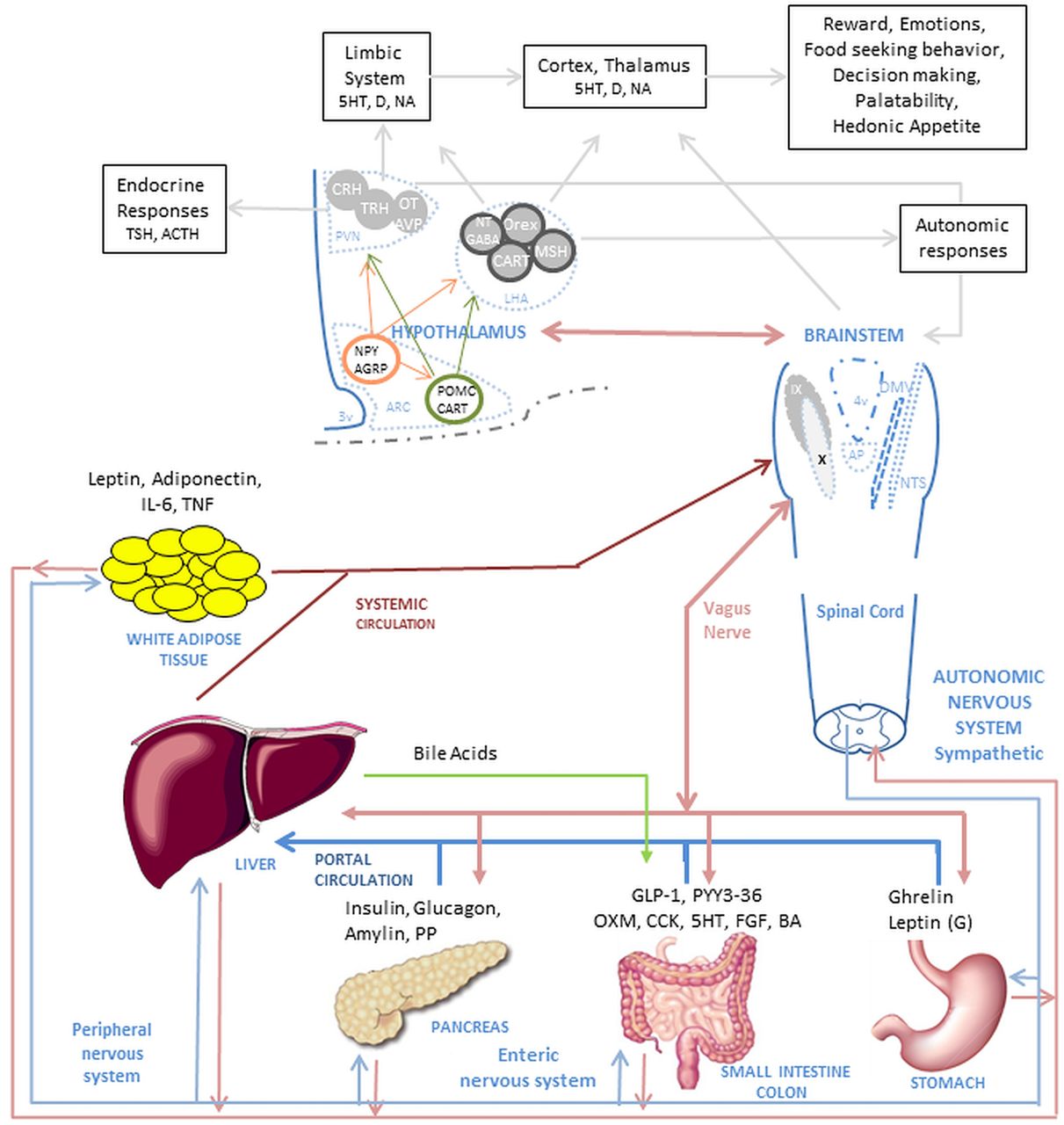
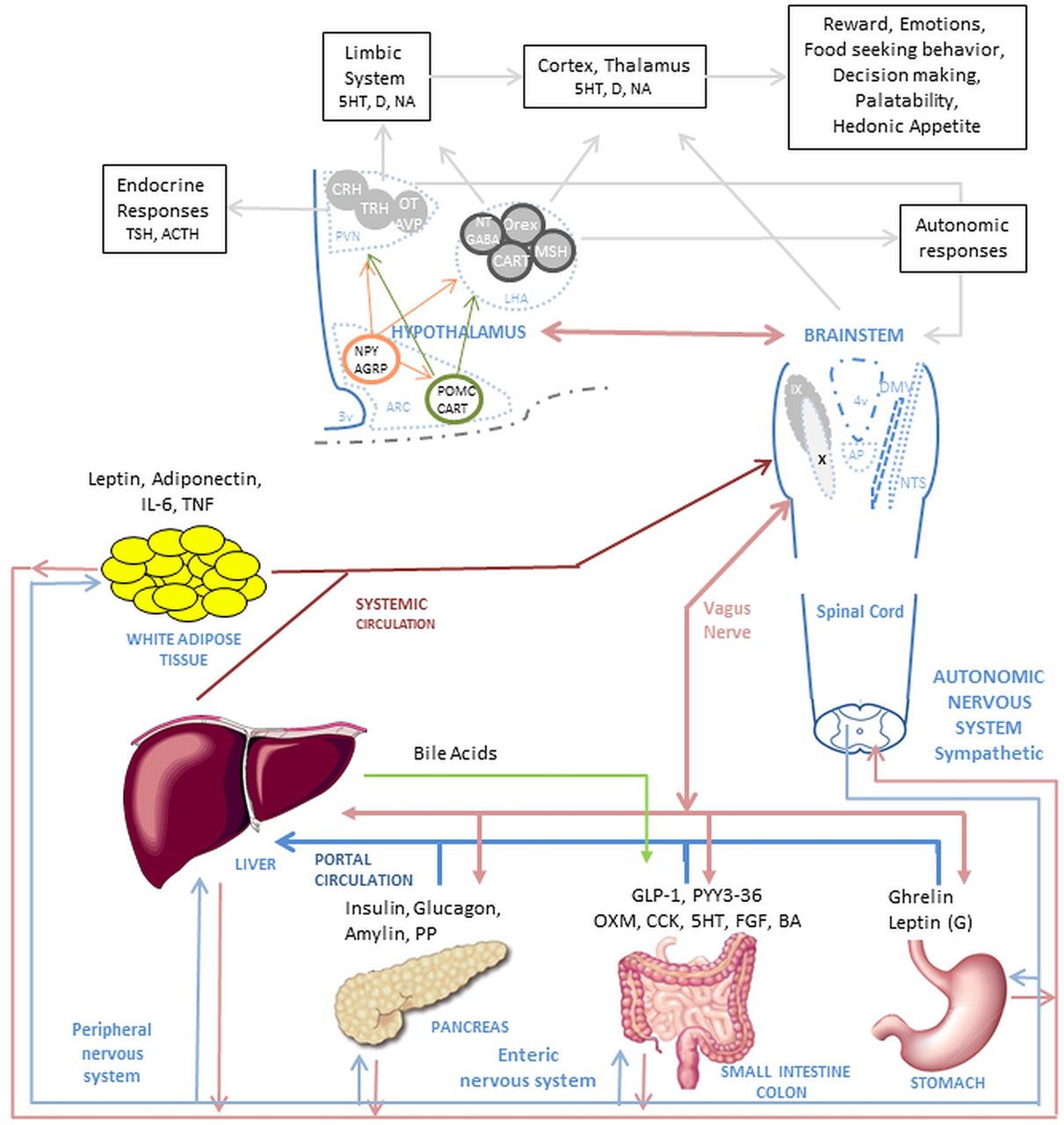
Complex mechanism of food intake regulation. The food intake process initiates when nutrients enter the GI tract. Digestion starts when the nutrients enter the stomach and produce mechanic-dilation decreasing acyl-ghrelin and increasing desacyl-ghrelin and gastric leptin. Stomach dilation sends signals through the vagus nerve and peripheral nervous system to the brainstem and hypothalamus. The digested nutrient passes to the small intestine and colon producing further mechanic-dilation, GI hormones release, bile acid and pancreatic juices secretion. These GI hormones have a local effect (paracrine) and peripheral effect, when secreted into circulation, passed through the liver and affect the muscle, adipose tissue, GI motility and function, and nucleus of the hypothalamus and brainstem. The paracrine and endocrine effect induces satiation and satiety. The muscle and adipose tissue release hormones which affect similar nuclei in the brain. The effect on the hypothalamus and brainstem trigger higher brain area responses, modulating behaviour and enhancing nutrient-related reward. In the hypothalamus, first order neurons in the arcuate nucleus (ARC) modulate appetite by NPY/AGRP pathway and satiation by the POMC/CART pathway. The neurons interact with second order neurons in the Paraventricular nucleus (PVN) and Lateral hypothalamic (LHA) area to send signals to higher brain areas and to the brainstem. In the brainstem, the Nucleus of the tractus solitarius (NST) and dorsal vagal complex (DMNV) interact with the periphery and GI system and brings signals to the higher brain areas and the hypothalamus. Abbreviations in alphabetical order: 5-HT, serotonin; ACTH, adrenocorticotropic hormone; AGRP, agouti-related peptide; ARC, arcuate nucleus; AVP, arginine vasopressin; BA, bile acids; CART, cocaine- and amphetamine-regulated transcript; CCK, cholecystokinin; CRH, corticotropin-releasing hormone; D, dopamine; DMNV, dorsal vagal complex; FGF, fibroblast growth factor -19; GABA, gamma-aminobutyric acid; GLP-1, glucagon-like peptide-1; IL-6, interleukin-6; LHA, lateral hypothalamic; MSH, melanocortin stimulating hormone; NA, noradrenaline; NPY, neuropeptide Y; NST, nucleus of the tractus solitaries; NT, neurotensin; OT, oxytocin; Orex, orexin; OXM, oxyntomodulin; PP, pancreatic polypeptide; PVN, paraventricular nucleus; peptide tyrosine–tyrosine (PYY3)-36, peptide tyrosine-tyrosine 3-36; POMC, proopiomelanocortin; TNF, tumour necrosis factor; TRH, thyroid-releasing hormone; TSH, thyroid-stimulating hormone.
Search strategy and selection criteria
References for this review were identified through searches in PubMed for articles published from June 1974 to August 2013 by use of the terms “brain-gut axis”, “food intake”, “vagus nerve”, “GI hormones”, “treatment”, “guidelines”, “bariatric surgery”, “gastric sleeve”, “sleeve gastrectomy”, “biliopancreatic diversion” and “endoscopic weight loss procedures” in combination with the term “obesity”. Articles resulting from these searches and relevant references cited in those articles were reviewed. Articles published in English were included.
Current state of obesity treatment
Despite information on complex neural, hormonal, metabolic and inflammatory mechanisms in obesity, bariatric–metabolic surgery is the only current treatment for obesity that is effective in the long term. Lifestyle modification and current pharmacological approaches are generally associated with modest (average 5 kg) weight loss that is poorly sustained in a majority of patients.1 The paradox between the relatively low efficacy of treatments targeting the complex neurohormonal mechanisms in obesity and the greater efficacy of restrictive or malabsorptive surgery is illustrated by their effects on cardiovascular mortality. The Swedish Obese Subjects Trial is the largest prospective bariatric surgery study with long-term follow-up, and it showed a significant reduction in cardiovascular mortality2; in contrast, the Look Ahead Trial, a randomised, controlled, National Institutes of Health (NIH)-sponsored trial in 5145 overweight/obese adults with type 2 diabetes designed to determine the long-term health benefits of weight loss achieved by lifestyle modification, showed no difference in cardiovascular mortality after 9 years of follow-up.3
Non-surgical interventions
The two main non-surgical approaches for the treatment of obesity and related complications are intense lifestyle modification and medications. Guidelines, based on Category A evidence and published by the National Heart, Lung and Blood Institute in 1998, suggest “that a 10 percent reduction in body weight reduces disease risk factors. Weight should be lost at a rate of 1 to 2 pounds per week based on a calorie deficit of 500–1000 kcal/day”.4 The guidelines also recommend increased physical activity and pharmacological approach to augment weight loss. This approach has been efficacious in multiple large-scale clinical trials: Diabetes Prevention Program showed that intense lifestyle modification prevented by 58% the incidence of diabetes when compared with placebo controls.5 Similarly, the Look Ahead Trial showed that intense lifestyle intervention resulted in 7% weight loss and improved diabetes control.6
In recent years, somewhat efficacious pharmacological approaches (such as sibutramine and rimonabant) received initial regulatory approval but were subsequently withdrawn from the market because of adverse effects such as depression or hypertension resulting from their central actions on adrenergic, serotonergic or cannabinoid mechanisms. Until recently, the only approved medication was the pancreatic lipase inhibitor, orlistat, which is associated with relatively small changes in weight and GI adverse effects, such as bloating and diarrhoea, which reduce compliance with orlistat intake over the long term. New pharmacological approaches, recently approved by the FDA, decrease appetite and result in weight loss: Lorcaserin (Belviq) is a serotonin 2c (5-HT2C) receptor agonist that activates pro-opiomelanocortin (POMC) neurons of the hypothalamic arcuate nucleus, decreasing appetite7 and resulting in an average 5.8% weight loss when compared with 2.1% in the placebo group.8 A second, approved drug is the combination phentermine-topiramate extended release (ER) (Qsymia) that produces mean 8–10% weight loss in different trials when compared with 1.6% weight loss in the placebo group.9 This degree of weight loss is lower than that observed with bariatric surgery. In view of the potential for central nervous system (CNS)-mediated and cardiovascular adverse effects with these centrally-acting drugs, they require close postmarketing surveillance. Lorcaserin has high affinity and selectivity and is a full agonist for 5-HT2C receptors. By virtue of its high selectivity for 5-HT2C receptors relative to 5-HT2A (18-fold) and 5-HT2B (104-fold) receptors,10 lorcaserin avoids induction of hallucinations11 and cardiopulmonary toxicity,12–14 respectively. Nevertheless, the FDA approved phentermine-topiramate ER (Qsymia) with 10 postmarketing requirements.15
Bariatric (Metabolic) surgery
Bariatric surgery remains the most effective treatment option for obese patients. Available procedures include laparoscopic and open Roux-en-Y gastric bypass (RYGB), sleeve gastrectomy, adjustable gastric band, vertical banded gastroplasty, duodenal switch and biliopancreatic diversion. RYGB is currently the bariatric surgical procedure of choice. In a meta-analysis of 136 studies including 22 094 patients, RYGB resulted in an average excess body weight loss of 62%, with resolution of diabetes in 84%, of hypertension in 68%, and of obstructive sleep apnoea in 81%, and improved hyperlipidaemia in 97%.16 ,17 Unlike medications and lifestyle modifications, the effects of bariatric surgery seem to be sustained in the long term. Thus, the recently updated Swedish Obese Subjects Study demonstrated mean changes in body weight after bariatric surgery (specifically with 13% RYGB, 19% gastric banding and 68% vertical banded gastroplasty) −23% (at 2 years), −17% (at 10 years), −16% (at 15 years) and −18% (at 20 years).2
Despite proven efficacy and the fact that mortality from bariatric surgery is comparable to that of cholecystectomy or appendectomy in bariatric centres with high surgical volumes, it is estimated that less than 1% of obese subjects who qualify for bariatric surgery will undergo such intervention.18 High costs and early and late complications of bariatric surgery are the main hurdles for widespread use. Early complications include anastomotic leaks, internal hernias, thromboembolic events, bowel obstruction, GI haemorrhage and wound complications. Late complications include gallstones formation, marginal ulceration, anastomotic stricture, incisional hernia, gastro-gastric fistula, dumping syndrome, micronutrient deficiencies and weight regain.
Given the current state of therapies for obesity, it is timely to question the 1998 guidelines that recommend slow weight loss with non-surgical approaches that produce modest and poorly sustained efficacy and potential centrally mediated side effects. The next sections review advances in our understanding of central and peripheral pathways controlling energy intake and expenditure in that order and discuss the potential to develop effective, minimally invasive treatments for obesity.
Targeting the central nervous system in obesity treatment
Appetite and satiety are controlled by hypothalamic centres in connection with the limbic (emotions and reward) and the autonomic nervous systems. These systems may provide novel targets for development of drug treatment for obesity. There are fenistrations in the blood–brain barrier (BBB) that are limited to specific brain regions, specifically the area postrema in the floor of the fourth ventricle and the subfornical organ and organum vasculosum of the lamina terminalis in the anterior wall of the third ventricle that are highly interconnected with the supraoptic, median preoptic and paraventricular nuclei of the hypothalamus. The latter organs play a role in water homeostasis and associated behaviours (thirst). In contrast, area postrema and the associated nucleus of the solitary tract (NST) and dorsal motor nucleus of the vagus nerve (DMNV) are the more likely locations for GI neuropeptide ‘sensing’ by the CNS.19 Regardless, either the fenistrations or specific transport processes are able to deliver gut or adipose-related hormones to these pivotal brain centres.19 ,20
The hypothalamic nuclei
The CNS coordinates a complex and dynamic process to regulate energy balance. Food intake, appetite and satiation are mainly regulated by the gut–brain axis,21 ,22 that is, gut hormones, the vagal complex, brainstem, medial basal hypothalamus and higher cortical brain centres.21 The current overarching hypothesis is that appetite is stimulated by the secretion of acyl-ghrelin (AG) from the stomach during the fasting period. Ghrelin acts on specialised neurons in the arcuate nucleus of the hypothalamus to activate the agouti-related peptide/neuropeptide Y (AgRP/NPY) pathway.23–25 This pathway is responsible for stimulating appetite centres in the cerebral cortex that are involved in food-seeking behaviour and prepare the GI tract for food intake by a vagal response.26 Satiation (or the feeling of fullness after meal ingestion) and satiety (absence of appetite) are induced by several hormones including leptin, insulin, glucagon, cholecystokinin, peptide YY, oxyntomodulin (OXM) and glucagon-like peptide-1 (GLP-1), which are secreted from the adipose tissue, pancreas and GI tract after food intake. These gut hormones inhibit the AgRP/NPY pathway in the arcuate nucleus and stimulate the POMC/α-melanocyte stimulation hormone (αMSH) pathway.22 ,27 The neurons in the arcuate nucleus are called ‘first-order’ neurons because of their ‘direct’ contact with peripheral hormones that mediate satiation or satiety. ‘Second-order’ neurons arise from the paraventricular nucleus, lateral hypothalamic area and ventromedial nucleus of the hypothalamus.28 Through projections caudally to the NST and DMNV in the brainstem, these second order neurons stimulate the gastric vagal efferent fibres to delay gastric emptying by stimulating intra-gastric nitric oxide system (NOS) neurons, thus decreasing motility.29 ,30 Second-order neurons also increase energy expenditure by inducing hyperthermia,31 ,32 and cranially directed neurons stimulate higher brain centres and produce food reward or feeling of well-being after a meal.30 ,32–36 These neuronal circuits ultimately control energy intake and expenditure by interacting with the GI tract, peripheral nervous system and adipose tissue.
Current understanding of the role of the hypothalamus and its circuits in energy balance is based on in vitro and in vivo observations of the effects of hypothalamic injuries or knock down, knock out and/or overexpression of target genes or pathways. However, interpretation of these experimental models is complicated by the presence of redundant pathways or compensatory mechanisms that can adapt the response to modification of specific targets, receptors or mechanisms. Novel techniques applied in animal models, such as optogenetics (that can photo-stimulate specific neurons) or short hairpin RNA studies (that can ‘silence’ one or more specific gene targets), may provide more comprehensive insights of the control of appetite and satiation without evoking compensatory mechanisms. The application of such techniques on AgRP neurons showed this pathway can regulate feeding behaviour independent of the POMC pathway,37 challenging the current paradigm of interaction of the AgRP/NPY and the POMC/αMSH pathways and questioning accuracy of current knowledge based on genetically modified animal models.38
In humans, less invasive methods have been developed to study these hypothalamic nuclei. Ideally, one would like to measure the electrical activity (action potentials) in these nuclei before, during and after eating to determine their signalling mechanisms. Unfortunately, this is not possible with current techniques. A surrogate measure of neural activity is blood flow, specifically, as a brain area becomes electrically active, blood flow increases to supply oxygen and glucose and remove carbon dioxide.
Brain blood flow can be measured using MRI. Three general imaging techniques are available for measuring blood flow to the hypothalamus, each with their own strengths and limitations. The primary challenge for each of these techniques is the small size of the hypothalamic nuclei; for example, each half of the hypothalamus is only 3–4 mm thick, and the arcuate nucleus is a cell cluster within the hypothalamus measuring only a few hundred microns in diameter. MRI systems with higher magnetic field strengths (eg, 3 tesla (T), 7 T) are more beneficial for imaging the hypothalamus as they have the higher resolution necessary to accurately measure such small structures.
Contrast-enhanced perfusion imaging is the most common MR technique to measure blood flow and has the highest resolution. However, this technique requires an intravenous bolus injection of gadolinium contrast and, therefore, can only be performed once per day due to potential renal toxicity. ‘Functional MRI’ is a technique that measures relative changes in blood flow between two different alternating experimental conditions. However, this technique is better suited to experimental designs where the two conditions are varied relatively rapidly (on the timeframe of seconds); most studies of hunger and satiety occur in a timeframe of minutes and cannot easily alternate between different conditions. Furthermore, this technique is susceptible to artefacts from the sphenoid and ethmoid sinuses that limit evaluation of the hypothalamus.
Pulsed arterial spin labelling (PASL) is an MR technique that labels blood with a magnetic ‘tag’ as it enters the skull and images it in the brain a short time later. PASL is ideally suited for the study of the hypothalamus as it: (1) does not require intravenous contrast and therefore can be performed many times over a few hour period; (2) does not suffer from significant susceptibility artefact; (3) yields highly reliable and reproducible quantifiable measurements of blood flow; and (4) using a 3 T MRI scanner, has sufficient resolution to measure global hypothalamic blood flow while minimising the influence of neighbouring areas.
As it is a relatively new MRI technique, only a few PASL studies of obesity have been performed to date. For example, PASL was used to measure absolute blood flow changes in different regions of the brain in response to stimuli such as ingestion of different simple sugars; thus, there is differential stimulation of hypothalamic regions with glucose and fructose.39–41 This approach has the potential to understand the activation of anatomical regions of the hypothalamus in response to nutrients during postprandial satiation.
Further studies are required to understand the adaptive and complex processes and neuroplasticity in the hypothalamus of obese subjects and to explain the function of CNS centres in the weight gain in already obese subjects in a food-rich situation. Specifically, it is still unclear why the brain functions as though the individual is still ‘hungry’ despite the presence of obesity, or why it is so difficult to maintain weight loss after a period of food-deprivation.42 ,43 Some authors have claimed that the long-term effect of bariatric surgery is mediated by a change in hypothalamic plasticity and resetting of the homeostatic ‘set point’, that is, the hypothalamic adaptive response that sets the total caloric intake per day.44 Experimental data supporting such a set point are changes in hypothalamic activity after bariatric surgery in diet-induced obesity in rodents45; in contrast, other studies refute the presence of a set point since no changes in the hypothalamus were observed after bariatric surgery when compared with caloric restricted rodents.46–48
With the introduction of MRI with PASL, it may be possible to clarify whether there are changes in the hypothalamic activity in humans during different stages of the postprandial period (cephalic, gastric and enteric) as well as in response to pharmacological, behavioural or surgical interventions.
The continuing challenge is to develop CNS targeted therapies that avoid effects on pathways unrelated to energy intake in the higher brain centres that may serve other physiological and psychological functions. Using novel imaging technologies, there is potential to appraise the responsiveness of specific hypothalamic and other brainstem and brain centres to macronutrients, a nutrient-poor or nutrient-rich environment, and to therapy directed at these redundant and complex pathways.
The brainstem–hypothalamic connections
The brain stem, through the predominantly afferent nucleus of the NST and the integrative and DMNV complex, controls the enteric nervous system in response to signals from energy levels in the periphery.26 ,30 The energy levels also influence brainstem nuclei in the area postrema in the floor of the fourth ventricle where there is a thin BBB.49 The DMNV has ascending and descending connections with the arcuate, paraventricular nucleus and lateral area of the hypothalamus and other higher brain centres to regulate food intake.30
Like the hypothalamus, the brainstem plays a major role in mediating brain–gut coordination through the nuclei mentioned above. As with the hypothalamus, the DMNV has a permeable BBB and is susceptible to the influence of circulating hormones such as incretins and glucose-regulatory hormones that may be associated with taste aversion in animal models and nausea in humans through activation of the area postrema in the brainstem.50
The role of the vagus nerve
The vagus nerve innervates most of the regions of the GI tract involved in energy intake, satiation and digestion,30 and it serves as a crucial link between the brain, brainstem and gut.
The afferent neuronal fibres of the ventral and dorsal vagal trunks in the abdomen are involved in regulating appetite and satiation. The vagus nerve is directly stimulated by mechanical stretch or change in viscus tension when food, or later chyme, passes through the lumen of the GI tract. The vagus nerve is also stimulated indirectly by neurohormonal mechanisms activated by the same mechanical and chemical stimuli arising from the gut. In the stomach, ghrelin secretion inhibits afferent vagal fibres to induce appetite—an orexigenic effect. Leptin secreted in the stomach stimulates vagal fibres and induces satiety—an anorexigenic effect. In the rest of the GI transit of chyme, the vagal afferent fibres are stimulated by other anorexigenic hormones such as cholecystokinin (CCK), GLP-1 or peptide YY.
Efferent vagal neuronal fibres control much of the motor and secretory apparatus of the digestive tract involved in food digestion and absorption. Partial vagotomy, or total sub-diaphragmatic vagotomy, or intermittent vagal nerve electrical stimulation to inhibit vagal function in humans51 decreased food intake and induced early satiety and weight loss.
The vagus nerve plays a dual role, interacting with anorexigenic and orexigenic pathways. These interactions appear to be altered in obesity.52 Greater understanding of methods to modulate vagal activity to induce weight loss could conceivably arise through stimulation of the anorexigenic pathway or inhibition of the orexigenic pathway. Clearly, further studies are needed to understand the effect of the vagus nerve in weight loss and glycemic control.
Targeting the gut in the treatment of obesity
Bariatric surgery
Bariatric surgery, especially RYGB, has thus far proved to be the most effective intervention for obesity and type 2 diabetes. The exact mechanism by which RYGB exerts its effects on weight loss and glycemic regulations is not fully understood; the timing of resolution of diabetes suggests that RYGB's effects on diabetes are independent of the degree of weight loss. Animal and human studies suggest that the restriction of food reservoir and malabsorption is insufficient to account for RYGB's effects to enhance satiation and satiety, improve food seeking behaviours and food selection, increase energy expenditure and improve glucose homeostasis.53–55 Thus, further studies are required to clarify physiological alterations in gut neuroendocrine signalling, GI motility, autonomic nervous system signalling, bile acid production and absorption, gut microbiota and other potential interactions with the CNS, pancreas, liver, muscles and adipose tissue in response to RYGB.
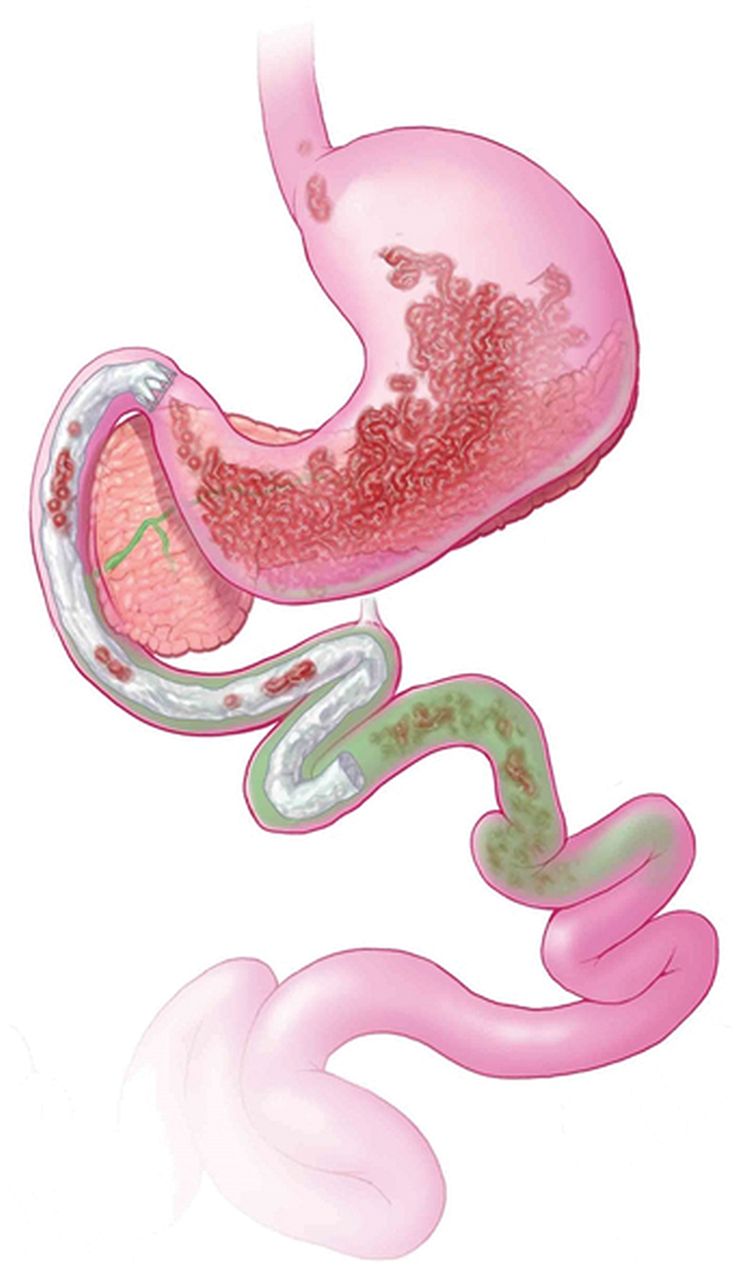
RYGB surgery is a complex procedure with at least five distinct components, all of which may have biological relevance in the induction of weight loss or ameliorating hyperglycaemia: (1) isolation of the gastric cardia by creation of a small gastric pouch, (2) exclusion of the distal stomach from contact with food, (3) exclusion of the proximal intestine from contact with food, (4) exposure of the jejunum to partially digested nutrients and (5) partial vagotomy (figure 2). Each of these distinct components of RYGB has the potential to alter appetite, energy absorption and expenditure, and glucose homeostasis; thus, replication of these individual anatomic alterations can be novel approaches to develop peripherally targeted treatments for obesity. The next section analyses each of these distinct components.

Biological relevance of Roux-en-Y gastric bypass (RYGB) surgery. RYGB is a complex procedure with at least five distinct components, all of which may have biological relevance in the induction of weight loss or ameliorating hyperglycaemia: (1) isolation of the gastric cardia by creation of a small gastric pouch, (2) exclusion of the distal stomach from contact with food, (3) exclusion of the proximal intestine from contact with food, (4) exposure of the jejunum to partially digested nutrients (5) and partial vagotomy.
The gastric cardia and ghrelin
Isolation of the gastric cardia and exclusion of the distal stomach may initially limit caloric intake by induction of nausea (and rarely vomiting), thereby discouraging overeating. In addition, the stimulation of the gastric mechanical and chemical receptors, rapid pouch emptying and release of ghrelin may also contribute to the induction of weight loss.56 ,57 Ghrelin is a peptide produced from the pre-pro ghrelin gene, mainly in the stomach. It is known to increase gastric emptying and to undergo cyclical changes in blood concentrations in the fasting and postprandial periods, being highest during fasting. AG is metabolised by the ghrelin activating enzyme, ghrelin-O-acyltransferase (GOAT), to deacyl-ghrelin (DAG). AG and DAG have different physiological effects: AG increases gastric emptying and appetite, whereas DAG decreases gastric emptying, induces postprandial fullness, and improves insulin sensitivity.58 The published literature suggests there is a decrease in plasma ghrelin post-RYGB, with attenuation of the cyclical changes in ghrelin levels.59 ,60 The literature is unclear about effects of AG or its metabolite, DAG, whose actions are different, as detailed above. Thus, the contribution of the changes in plasma ghrelin concentrations to the clinical efficacy of RYGB requires further investigation.
Bypass of the proximal small intestine
The proximal small intestine is extremely efficient in absorption of monosaccharides,61 and it plays a major role in glucose homeostasis and in the pathogenesis of diet-induced diabetes. Wang and colleagues have shown that direct administration of lipids into the upper intestine of non-diabetic rats suppressed glucose production through an intestine–brain–liver neural axis; this effect was not observed in rats with diet-induced diabetes.62 Furthermore, Goto-Kakizaki (GK) type 2 diabetic rats that undergo duodeno-jejunal bypass (DJB) develop markedly improved oral glucose tolerance compared with rats that undergo gastrojejunostomy (which does not bypass the duodenum); these effects are reversed with re-establishing flow through the duodenum.63
Patients who undergo DJB have greatly improved glycemic control with only minimal effect on body weight.64 ,65 However, diabetes remission rates are much lower after DJB compared with traditional RYGB, suggesting that other effects of RYGB are important in weight loss and that diabetes remission rates may be proportional to the degree of weight loss.66 On the other hand, vertical sleeve gastrectomy (VSG) does not bypass the duodenum and produces significant weight loss and glycemic improvement. In a randomised controlled trial, VSG was associated with weight loss comparable to that observed with RYGB.67 The incretins and insulin-glucose responses were also similar 3 months after the procedures, although RYGB had a greater response than VSG earlier than 3 months postsurgery.68 The potential additional role of the bypassed duodenum in ameliorating glycemic control, independent of the effects of incretins, is supported by a recent meta-analysis which showed that RYGB surgery produced a higher rate of remission of type 2 diabetes when compared with VSG (OR 2.46 CI 1.48 to 40.9).69–72 Further studies are needed to understand the differences in these bariatric surgeries and to ascertain if the glycemic improvement is due to weight loss alone or whether the bypass of the duodenum is really necessary.
Incretins
Within the mucosa of the small intestine, enteroendocrine cells express functional gustducin-coupled sweet taste receptors that ‘taste’ luminal nutrients and release gut peptides that mediate satiety and enhance insulin secretion (incretins).73 ,74 These peptides exert their effects through interaction with the afferent vagal and spinal nerve fibres innervating the gut, or by entering the blood stream to function as hormones, influencing the area postrema or hypothalamic areas through the locally permeable BBB.
GLP-1 is the major known incretin, synthesised from the enteroglucagon or pre-proglucagon gene in the enteroendocrine L-cells, which are located in the proximal small bowel, but are more abundant in the distal small bowel and colon.75 In addition to GLP-1, the enteroglucagon gene encodes multiple peptides involved in energy balance and glycemic control; these include glucagon and OXM.
GLP-1 is released in response to contact with luminal nutrients or bile acids, and by submucosal neural stimulation mediated by voltage-gated Ca++ channels.76 GLP-1 inhibits glucagon secretion, stimulates glucose-dependent insulin secretion, increases insulin sensitivity, induces nausea and satiety and is associated with weight loss.77–79 GLP-1 levels are blunted in response to luminal glucose in type 2 diabetes, resulting in an early defective insulin response to the luminal glucose load.80 ,81
GLP-1 and insulin levels significantly increase in the early postprandial period after RYGB, an effect that is not observed with laparoscopic adjustable gastric band or diet-induced weight loss because the latter do not accelerate gastric emptying of nutrients.82 ,83 The early postprandial increase in these peptide levels results from enhanced delivery of partially digested nutrients to the distal small intestine, directly stimulating the L cells. Indeed, isoperistaltic ileal transposition above the ligament of Treitz (which accelerates delivery of nutrients to the distal small bowel) in diabetic rats and humans resulted in significant improvement in GLP-1 production in response to glucose and improved glucose homeostasis.84–86 The endogenous GLP-1 effect on glucose homeostasis and GI motility is sustained when examined an average of 5 years after RYGB.87 In a randomised, controlled trial, the GLP-1 agonist, liraglutide, 3 mg, produced a significant mean weight loss of 7.2 kg compared with 2.8 kg in the placebo group.88 Conversely, studies in rodents have shown that the effect of weight loss and glycemic improvement after VSG is independent of GLP-1 activity.89
Peptide tyrosine–tyrosine (PYY) is another gut hormone with significant effect on the ileal brake and regulation of food intake. Pharmacological doses of PYY induced satiety;90 ,91 PYY is also increased after RYGB92A large phase II clinical trial of intranasal peptide YY3–36 had to be discontinued early due to severe nausea and vomiting,93 which is most likely explained by PYY stimulating the area postrema in the brainstem resulting in nausea50 or delay in gastric emptying induced by PYY3–36.94–96
Quantifying the contribution of the rise in GLP-1 and other incretins to the therapeutic effects of RYGB and weight-independent improvement in glycemic control requires further study. While further information may lead to novel applications of incretins in weight loss therapy, experience with GLP-1 and intranasal PYY3–36 suggests that adverse effects such as nausea may limit the potential clinical efficacy.
Autonomic denervation in RYGB
The vagus nerve enters the abdomen from the oesophageal plexus as anterior and posterior nerve trunks. Denervation of sub-diaphragmatic branches of these trunks may occur after RYGB. The contribution of this denervation to weight loss and metabolic changes after RYGB is unclear.
Through the actions of melanocortin-4 receptors (MC4R) in the DMNV and thoracic sympathetic preganglionic neurons, the autonomic nervous system plays an important role in the improved insulin sensitivity and enhanced energy expenditure after RYGB.97 Furthermore, a variant in the human MC4R (I215 L) locus that enhances basal activity of the receptor is associated with improved outcomes after RYGB. The peripheral signals mediating enhanced activation of central MC4R signalling are yet to be identified.98 Further studies are needed to understand the role of MC4R signalling in satiety.
Microbiome in RYGB
Emerging evidence suggests that microbes that reside in the human GI tract can contribute to development of obesity. As a dynamic ecosystem, the microbiota closely interacts with host metabolism, for example, by breaking down otherwise indigestible carbohydrates and increasing short chain fatty acid absorption in the colon. This provides the host with additional energy and increases fat storage in adipose tissue.100 ,101 The microbiota composition in obese individuals appears to be structurally and functionally different from that of lean individuals, and it appears to be adapted to more efficient energy extraction and storage, contributing to obesity and its metabolic consequences. This adaptation is termed ‘obesity associated dysbiosis’.102
Recently, RYGB has been shown to cause shifts in faecal microbiota profiles, potentially contributing to some of the favourable effects of RYGB.103–105 In mice, changes in gut microbiota after RYGB seem to be mediated by anatomical rearrangement of the GI tract, rather than being a response to food restriction mediated weightloss.105 However, these findings have not been replicated in humans. It is also unclear which anatomical or functional component of the RYGB is predominantly responsible for the change in gut microbiota; the potential candidate mechanisms include changes in gastric pH, bile acid production and circulation, GI tract motility, gut hormonal profiles, and gut–brain interaction, each of which can independently result in a change in the gut microbiota.106
In summary, the effects of bariatric surgery are fundamentally different from those of restrictive diets. The latter often lead to physiological responses to weight loss that actually increase hunger and decrease energy expenditure, resulting in regain of the lost weight.1 ,99 The long-term efficacy of RYGB provides a paradigm shift in our understanding of the physiological mechanisms of reduced appetite and weight loss and the physiological responses which differ from those operant in diet-induced weight loss. The success of RYGB encourages focus on therapeutic approaches in obesity directed at the GI tract.
Peripheral interventions as alternatives to bariatric surgery
The previous discussion suggests five anatomical alterations may mediate the effects of RYGB on energy intake and expenditure. It also suggests that a multitude of physiological adaptations in the homeostatic mechanisms involved in body weight regulation may need to be addressed to overcome the compensatory responses leading to weight regain seen with restrictive diets.
Emerging technologies have opened the door to the use of approaches and devices to reproduce endoscopically many of the anatomical alterations of bariatric surgery. Targeting peripheral mechanisms has the potential to provide effective treatment of obesity and its associated disorders in a cost-effective and minimally invasive alternative to traditional bariatric surgery. This approach may also be applied in those with moderate obesity, vulnerable populations, such as children and adolescents, and at-risk superobese individuals.107
Endoscopic implantation of duodenal-jejunal bypass sleeve made from a Teflon liner (EndoBarrier, GI Dynamics, Lexington, Massachusetts, USA) shows promise and efficacy in the management of obesity and associated diabetes.108 ,109 When deployed in the duodenal bulb, this impermeable fluoropolymer sleeve, extending 60 cm into the small bowel, creates a mechanical barrier that allows food to bypass the duodenum and proximal jejunum, thus potentially manipulating the enteroinsular system (figure 3). Several prior studies have documented the technique's feasibility and efficacy on weight loss and on comorbidity such as non-alcoholic fatty liver disease.108–112 Recent rodent studies showed that the diversion of nutrients away from the duodenum decreases food intake, improves glucose homeostasis, increases circulating bile acids and increases small intestine villus length113 A pivotal U.S. multicentre FDA registry trial for this device is currently underway. The impact of this device in the long term requires elucidation as insulin sensitivity improved by >50% as early as 1 week after implantation, but there was a trend towards deterioration in all the above-mentioned variables 26 weeks after explantation.114
{kind=link}
{kind=link}
{kind=link}
Duodenal-jejunal bypass sleeve. The duodenal-jejunal bypass sleeve deploys in the duodenal bulb, this impermeable fluoropolymer sleeve extending 60 cm into the small bowel creates a mechanical barrier that allows food to bypass the duodenum and proximal jejunum, thus potentially manipulating the enteroinsular system (EndoBarrier, GI Dynamics, Lexington, Massachusetts, USA).
Recent advances in flexible endoluminal endoscopy now provide the necessary endoscopic suturing tools for transoral endoscopic gastric volume reduction in a fashion similar to sleeve gastrectomy. The feasibility and short-term efficacy of an endoscopic sleeve gastroplasty for the treatment of obesity has been recently reported.115
Conclusions
The obesity epidemic requires concerted effort, including behavioural, societal, legislative and therapeutic interventions. We have focused on the therapeutic interventions and identified challenges and opportunities to impact the unmet needs with non-surgical interventions (table 1). Despite the impact of bariatric surgery, we believe that further insights on vagal, hypothalamic and GI hormonal control are key to the development of interventions that can be applied in the large number of eligible patients with obesity and metabolic syndrome. It is also conceivable that future interventions targeting the microbiome or intraluminal mechanisms such as interference with intraluminal digestion or bile acid-related augmentation of incretin responses (not discussed here) may be feasible, peripherally-directed therapies. Meanwhile, the search for the magic bullet in central control mechanisms will continue, though it remains paradoxical that, despite all the drug development programmes being centred on hypothalamus or central mechanisms controlling appetite, the peripheral targets of bariatric surgery constitute the most salient direction for practical and effective obesity treatment at present.
Paradox between bariatric (RYGB) surgery and pharmacological interventions for obesity
Acknowledgments
We thank Cindy Stanislav for secretarial assistance.
References
Footnotes
-
Contributions AA, BKA-D and MC were responsible for study conceptualisation and writing of manuscript. JDP was responsible for writing of manuscript.
-
Funding National Institutes of Health to MC for studies in obesity (NIH RO1- DK67071).
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.