Article Text

Abstract
Objective To assess a new adenovirus-based immunotherapy as a novel treatment approach to chronic hepatitis B (CHB).
Methods TG1050 is a non-replicative adenovirus serotype 5 encoding a unique large fusion protein composed of a truncated HBV Core, a modified HBV Polymerase and two HBV Envelope domains. We used a recently described HBV-persistent mouse model based on a recombinant adenovirus-associated virus encoding an over length genome of HBV that induces the chronic production of HBsAg, HBeAg and infectious HBV particles to assess the ability of TG1050 to induce functional T cells in face of a chronic status.
Results In in vitro studies, TG1050 was shown to express the expected large polyprotein together with a dominant, smaller by-product. Following a single administration in mice, TG1050 induced robust, multispecific and long-lasting HBV-specific T cells detectable up to 1 year post-injection. These cells target all three encoded immunogens and display bifunctionality (ie, capacity to produce both interferon γ and tumour necrosis factor α as well as cytolytic functions). In addition, control of circulating levels of HBV DNA and HBsAg was observed while alanine aminotransferase levels remain in the normal range.
Conclusions Injection of TG1050 induced both splenic and intrahepatic functional T cells producing cytokines and displaying cytolytic activity in HBV-naïve and HBV-persistent mouse models together with significant reduction of circulating viral parameters. These results warrant clinical evaluation of TG1050 in the treatment of CHB.
- CELLULAR IMMUNOLOGY
- CHRONIC VIRAL HEPATITIS
- HEPATITIS B
- IMMUNOTHERAPY
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Significance of this study
What is already known about the subject?
HBV chronic infection can be controlled but is rarely cured.
HBV immunotherapeutics have been described at preclinical and clinical levels with limited or no efficacy.
This novel class of HBV therapeutics has included HBsAg in the majority, the Core antigen in some and only one coding for the HBV polymerase.
HBV immunotherapeutics can induce functional T cells in HBV transgenic mice, typically mice transgenic for a single HBV antigen, but an effect on viral parameters has seldom been reported following their administration, particularly on levels of circulating HBsAg.
What are the new findings?
TG1050 is the only HBV immunotherapeutic covering in a single entity three HBV antigens/domains, including polymerase.
TG1050 induces persistence of multifunctional HBV-specific T cell responses up to 400 days after a single injection.
Following single as well as multiple injections, TG1050 can educate functional T cells in a HBV chronic environment and display significant and persistent antiviral activity, in particular an impact on the level of HBsAg.
TG1050 is the only adenovirus-based HBV immunotherapeutic currently planned for testing in the clinic.
How might it impact on clinical practice in the foreseeable future?
TG1050 will next be tested in the clinic in combination with nucleos(t)ide analogues in the treatment of chronic hepatitis B (CHB) and aims to increase the cure rate.
If successful, TG1050 will bring a novel treatment paradigm to patients with CHB.
Introduction
Infection by HBV is one of the major causes and risk factors for developing liver cancer.1 Over 2 billion people have been infected by HBV worldwide, and about 240 million of them are currently chronically infected and at high risk of developing cirrhosis and hepatocellular carcinoma.2 Current chronic hepatitis B (CHB) therapies include nucleos(t)ide analogues (NUC) aimed at inhibiting viral replication3 ,4 and pegylated interferon α (IFNα).5 ,6 Despite the ability of these treatments to control HBV replication in the great majority of patients and to improve liver histology, complete cure of HBV is achieved in only 3–5% of patients. Therefore, most patients require costly life-long treatments.
In patients resolving infection, development of broad and robust CD8+ and CD4+ T cell responses targeting multiple HBV antigens that produce cytokines and display cytolytic properties have been observed7 ,8 and correlated with virus control and/or elimination.9 In contrast, patients with CHB display weak, narrowed and dysfunctional HBV immune T cell responses.10–12 Among the new therapeutic arsenal being developed, immunotherapeutics aimed at inducing immune responses similar to those found in resolvers represent a growing field. Although first-generation HBV-specific immunotherapies have so far had limited success in the clinic, a number have been capable of inducing HBV-specific responses in patients with CHB.13 ,14 Most HBV-specific immunotherapeutics currently tested in clinical trials involve only 1–2 antigens and, except for a poxvirus-based candidate, none of them is based on a viral vector.15 ,16
TG1050 is a novel immunotherapeutic based on a non-replicative adenovirus 5 vector encoding a unique and large fusion protein composed of modified HBV Core and Polymerase and selected domains of the Env proteins. We show here that a single injection of TG1050 is broadly immunogenic, including in a HBV-persistent model,17 and displays antiviral properties.
Materials and methods
Animals
BALB/c, C57BL/6J and HLA-A*0201 transgenic H-2 class I KO mice (HLA-A2 mice)18 were bred at Charles River Laboratories (L'Arbresle, France) and HLA-A*0201/DRB1*0101 transgenic H-2 class I/class II KO mice (HLA-A2/DR1 mice)19 were bred at the Institut Pasteur (Paris, France). Animals receiving the adeno-associated virus (AAV)-HBV were manipulated in an A3 confinement facility (see online supplement).
TG1050 adenovirus-based vector
TG1050 is based on an E1, E3 deleted non-replicative human adenovirus serotype 5 (Ad5). It encodes a large fusion protein composed of, from its N-term part, a truncated form of the HBV Core protein fused to a deleted and mutated HBV Polymerase and to two HBV Envelope (or HBsAg) domains, as shown in figure 1 (derived from GenBank # Y07587). This vector was constructed, produced, purified and titrated as described elsewhere.20–22 An Ad5 encoding no antigen (AdTG15149, referred to as Empty Ad) was also used.
The HBV fusion protein encoded by TG1050 under a CMV promoter is schematically represented. It is composed of a truncated HBV core (aa 1–148) fused to a deleted and mutated Polymerase (aa 1–832, Δ aa 538–544 and Δ aa 710–742 and mutations D689H, V769Y, T776Y, D777H) in which two selected domains of HBsAg are inserted (Env1 aa 14–51 inserted in deletion Δ aa 538–544 of Pol and Env2 aa 165–194 inserted in deletion Δ aa 710–742 of Pol).
AAV2/8 HBV- based vector
The recombinant AAV2/8 vector carrying 1.2 copies of the HBV genome was produced and titrated (virus genome (vg)/mL) as described elsewhere.17
Synthetic peptides and peptide library
Synthetic peptides corresponding to HLA-A2 or H-2b or H-2d restricted epitopes from HBV and adenovirus proteins as well as 15-mers included in HBV proteins and previously found to be reactive (unpublished data) were purchased from Eurogentec (Belgium) or Pro-immune (UK). Peptide amino acid sequences were identical to those of the TG1050 (see online supplementary table S1) and used at a concentration of 10 µM.
Synthetic peptide libraries were composed of 15-mers overlapping by 11 aa covering the HBV Core, Polymerase and the two Env domains included in TG1050. Pools were tested at a concentration of 5 µg/mL/peptide.
Mouse immunisation
Subcutaneous immunisation of BALB/c, C57BL/6J, HLA-A2 or HLA-A*0201/DRB1*0101 transgenic mice was performed with 108 IU (2×109 viral particles) of TG1050 or empty adenovirus.
For AAV2/8-HBV experiments, mice received a single tail vein injection of 1×1010 or 5×1010 vg of the AAV2/8-HBV as described previously.17 At day 32, 33, 54, 62 and 69 they were immunised with TG1050 or empty adenovirus.
Monitoring of induced T cell responses
IFNγ ELISPOT and intracellular cytokine staining (ICS) assays
IFNγ ELISPOT and double or triple IFNγ/tumour necrosis factor α (TNFα) and interleukin 2 (IL-2) intracellular cytokine staining (ICS) assays were performed on splenocytes of BALB/C, C57BL/6J and HLA-A2 mice as described in the online supplement and published previously.23 ,24 Assays were also performed as described for HLA A2/HLA DR1 mice.17 ,23
Pentamer and activation and memory marker staining
Whole blood collected on EDTA was incubated with PE-labelled MGL-H2Kb-pentamer (ProImmune, UK) and anti-CD8α-APC (clone 53-6.7), anti-CD44-FITC (clone IM7, BD Bioscience) and anti-CD62L-PE-Cy7 (clone MEL-14) or anti-CD8α-PerCP (clone 53-6.7, BD, Bioscience), anti-CD27-APC (Clone L.G.3A10, Biolegend) and anti-CD43-PE-Cy7 (Clone 1B11, Biolegend) antibodies as described previously.23 Red blood cells were lysed (FACS Lysing solution, BD, Bioscience) and collected cells were washed, fixed and assessed by flow cytometry (FACS Canto II, BD Bioscience). The data were analysed using the BD DIVA software.
Viral and biochemical parameters in AAV2/8-HBV injected mice
HBsAg was assayed with commercial ELISA kit (Bio-Rad, France) as previously described17 and HBV viraemia was assessed using an Architect Abbot automat or direct quantitative RT-PCR (see online supplement).
Results
TG1050 composition and in vitro characterisation of HBV protein expression via TG1050
Figure 1 depicts the organisation of the large HBV encoded polyprotein. This organisation resulted from the choice of HBV proteins known to be targeted by T cells during infection, in particular resolutive infection (eg, Core) and/or containing clusters of T cell epitopes (eg, Env1 and Env2), together with the need to generate a genetically stable vaccine fit for manufacturing (mutations in POL). In vitro expression of the HBV fusion protein was analysed by western blotting (see online supplementary figure S1A) and revealed a faint band corresponding to the Core-Pol-Env fusion protein (about 115 kDa, the calculated molecular weight (MW) being 113.9 kDa) as well as several additional bands corresponding to lower MW (ranging from 15 to 50 kDa). Products between 15 and 26 kDa were only detected with a Core-specific antibody and with a Pol-specific antibody targeting the N-terminus of the protein, while they were not detected with a Pol-specific antibody targeting aa 225–250 of the Polymerase (data not shown), suggesting that the observed products correspond to N-terminus cleavage products of the HBV fusion protein.
Analysis by bidimensional electrophoresis followed by immunoblot together with LC-MS/MS analysis was performed to characterise the different protein species generated (see online supplementary figure S1B and C). Data confirmed that products around 26 kDa are composed of the N-terminal part of the fusion protein—that is, the HBV Core protein and N-term amino acids of the Polymerase protein.
TG1050 induces high percentages of polyfunctional CD8+ T cells producing IFNγ and TNFα and displaying cytolytic activity in naïve mouse models
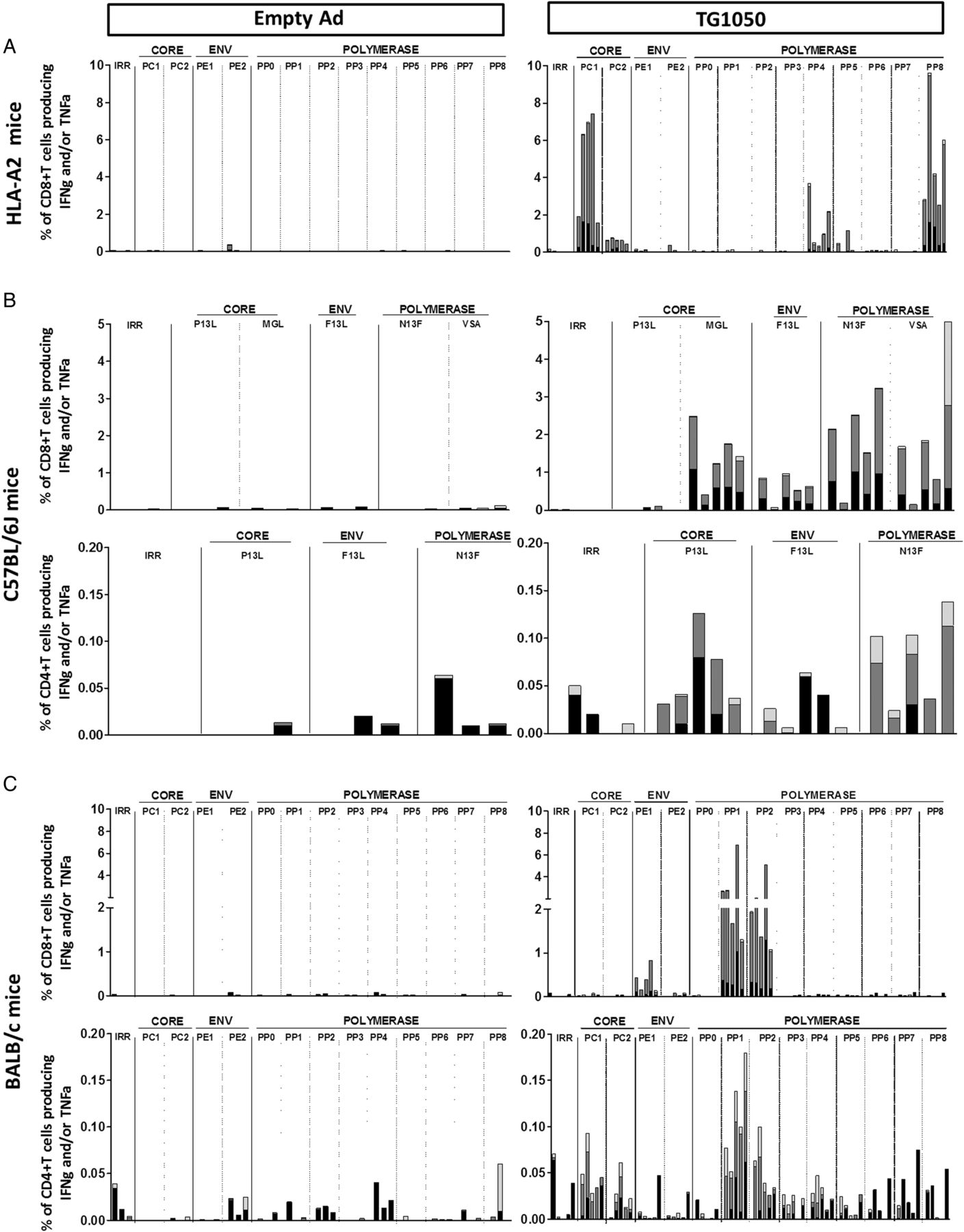
The ability of TG1050 to induce HBV-specific polyfunctional T cells was assessed by ICS assay in three mouse strains following one injection.
In HLA-A2 mice (figure 2A), TG1050 induced IFNγ-producing CD8+ T cells targeting the Core and Polymerase proteins (up to 7.5% and 9.5% of IFNγ+ CD8+T cells), most of them also producing TNFα. In C57BL/6J mice (figure 2B), IFNγ-producing CD8+ T cells targeting the three HBV antigens were detected (up to 2.4%, 1% and 3.2% of IFNγ+ CD8+ T cells specific for Core, Env and Polymerase respectively), with at least half of these cells producing both IFNγ and TNFα. Finally, in BALB/C mice (figure 2C), TG1050 induced IFNγ-producing CD8+ T cells specific of Env1 domain and Polymerase (up to 0.7% and 6.8% of IFNγ+CD8+ T cells specific for Env1 and Polymerase, respectively), most of them also producing TNFα. Overall, no (HLA-A2) or weak (C57BL/6J and BALB/c) CD4+ T cells were detected in these experiments. Frequencies of TG1050-induced IFNγ-producing cells were also assessed using IFNγ ELISPOT assay (see online supplementary figure S2). Responses targeting epitopes from all three encoded antigens were detected with variable magnitude and hierarchy depending on the mouse strain.

Analysis of induced T cell responses following single injection of TG1050 using interferon γ (IFNγ)/tumour necrosis factor α (TNFα) ICS assay in (A) HLA-A2 transgenic mice, (B) C57BL/6J mice and (C) BALB/c mice. HLA-A2 transgenic mice or C57BL/6J or BALB/c mice were immunised once subcutaneously with TG1050 or an empty Adenovirus (negative control) and HBV-specific immune responses were monitored on spleen cells using IFNγ/TNFα intracellular cytokine staining assays using single peptides or pools of peptides derived from Core, Env or Polymerase HBV proteins for stimulation. The results are presented as the percentage of IFNγ and/or TNFα producing CD8+ or CD4+ T cells for each group. Each bar corresponds to an individual mouse. For each mouse, the white bar represents the percentage of CD8+ or CD4+ T cells producing TNFα alone, the grey bar represents the percentage of CD8+ or CD4+ T cells producing IFNγ and TNFα and the black bar represents the percentage of CD8+ or CD4+ T cells producing IFNγ alone.
In vivo cytotoxic T lymphocyte (CTL) assays were performed to assess functional capacity of TG1050-induced CD8+ T cells in vivo in the three different mouse lines (see online supplementary figure S3). Cytolytic CD8+ T cells targeting Core and Polymerase (HLA-A2 and C57BL/6J, see online supplementary figure S3A and B) or Core, Pol and Env (BALB/c, see online supplementary figure S3C) were detected with percentages of lysis ranging from 6% to 83%. Of note, all three Pol and Core HLA-A2 epitopes described in patients with HBV were recognised by the induced T cells including the Core18–27 epitope (referred to as FLP) associated with resolution of infection.7 ,25 ,26
Overall, these data illustrate the capacity of TG1050 to induce potent, multi-antigenic, polyfunctional CD8+ T cells displaying in vivo cytolytic activity specific to all encoded HBV antigens.
TG1050 induces long-lasting functional HBV-specific T cells displaying recall potential
In C57BL/6J mice, HBV-Core pentamer staining showed a peak of Core-specific T cells in blood 14 days post-injection (mean of 3.4%), followed by a decrease up to day 96 (mean value around 0.6% at day 96, figure 3A, left panel). The percentage of Core-specific T cells progressively decreased but was still clearly detectable when measured at day 310 and in some mice up to day 400. Analysis of the CD44/CD62L phenotype—which allows effector memory cells to be distinguished from central memory cells—on Core-specific T cells showed that a single injection of TG1050 induces mostly effector memory cells (CD44+/CD62L− cells, percentages ranging from 95% at day 14 to 60% at day 400), but with the appearance of a significant proportion of central memory cells observed from day 310 (28% and 35% of Core-specific T cells being CD44+/CD62L+ at day 310 and day 400, respectively) (figure 3A right panel). Analysis of CD27/CD43 phenotypes on Core-specific T cells, in particular the CD27+/CD43− population which has been described as having good recall potential,27 showed that 20% of induced Core-specific cells were CD27+/CD43− during the first 100 days post-injection of TG1050. This proportion increased to 29% and 36% at the latest analysed time points.


Analysis of long-term HBV-specific T cells following a single injection of TG1050 up to 400 days post-injection. C57BL/6J mice were immunised once subcutaneously with TG1050 or an empty Ad (negative control). (A) HBV-specific cells were monitored and quantified along time in whole blood of injected mice by pentamer staining of HBV Core-specific T cells. An example of a dot plot obtained for one representative mouse injected with TG1050 at each time point is also shown on the right side of the graphs. Phenotypes of these Core-specific T cells were assessed by following CD44 and CD62L as well as CD27 and CD43 markers. Examples of dot plots obtained for CD62 L/CD44 and CD27/CD43 staining experiments in TG1050-injected mice are also shown at the top of the figure (shown for day 400). Pie charts representative of the mean percentages of HBV Core (MGL epitope)-specific CD8+ T cells displaying the indicated phenotypes are shown. (B, C). Interferon γ (IFNγ) ELISPOT (B) and intracellular cytokine staining (ICS) (C) assays realised at 400 days post-injection were performed using spleen cells and pools of overlapping peptides covering, respectively, the HBV Core protein (PC1, PC2), the 2 Env domains (PE1, PE2) and the HBV polymerase protein (PP4) or irrelevant peptides (IRR). For ELISPOT assay (3B), the horizontal dotted line represents the threshold of ELISPOT positivity. Each individual mouse is represented by an empty (empty Ad injected mice) or a plain (TG1050 injected mice) circle and the black line represents the mean of each group. For IFNγ/tumour necrosis factor α (TNFα) ICS assay (C), the results are presented as the percentage of IFNγ- and/or TNFα-producing CD8+ T cells for each group. For each mouse, the white bar represents the percentage of CD8+ T cells producing TNFα alone, the grey bar represents the percentage of CD8+ T cells producing IFNγ and TNFα and the black bar represents the percentage of CD8+ T cells producing IFNγ alone.
Functional HBV-specific T cells producing IFNγ remained detectable more than 1 year after one immunisation (figure 3B). The frequency of detected Pol-specific T cells was the highest (mean 290 spots/106 cells), followed by Core-specific T cells (mean 160 spots/106 cells) and Env1-specific T cells (mean 75 spots/106 cells). Half or more of HBV-specific CD8+ T cells detected 400 days after one immunisation were still able to produce both IFNγ and TNFα. These T cells targeted mainly Polymerase and Core antigens (figure 3C).
TG1050 is able to induce functional T cells and antiviral responses in HBV-persistent HLA-A2/DR1 transgenic mice following a single administration
TG1050 immunogenicity was evaluated in the recently described AAV-HBV-persistent model.17 Both short-term and long-term studies were performed.
Short-term monitoring
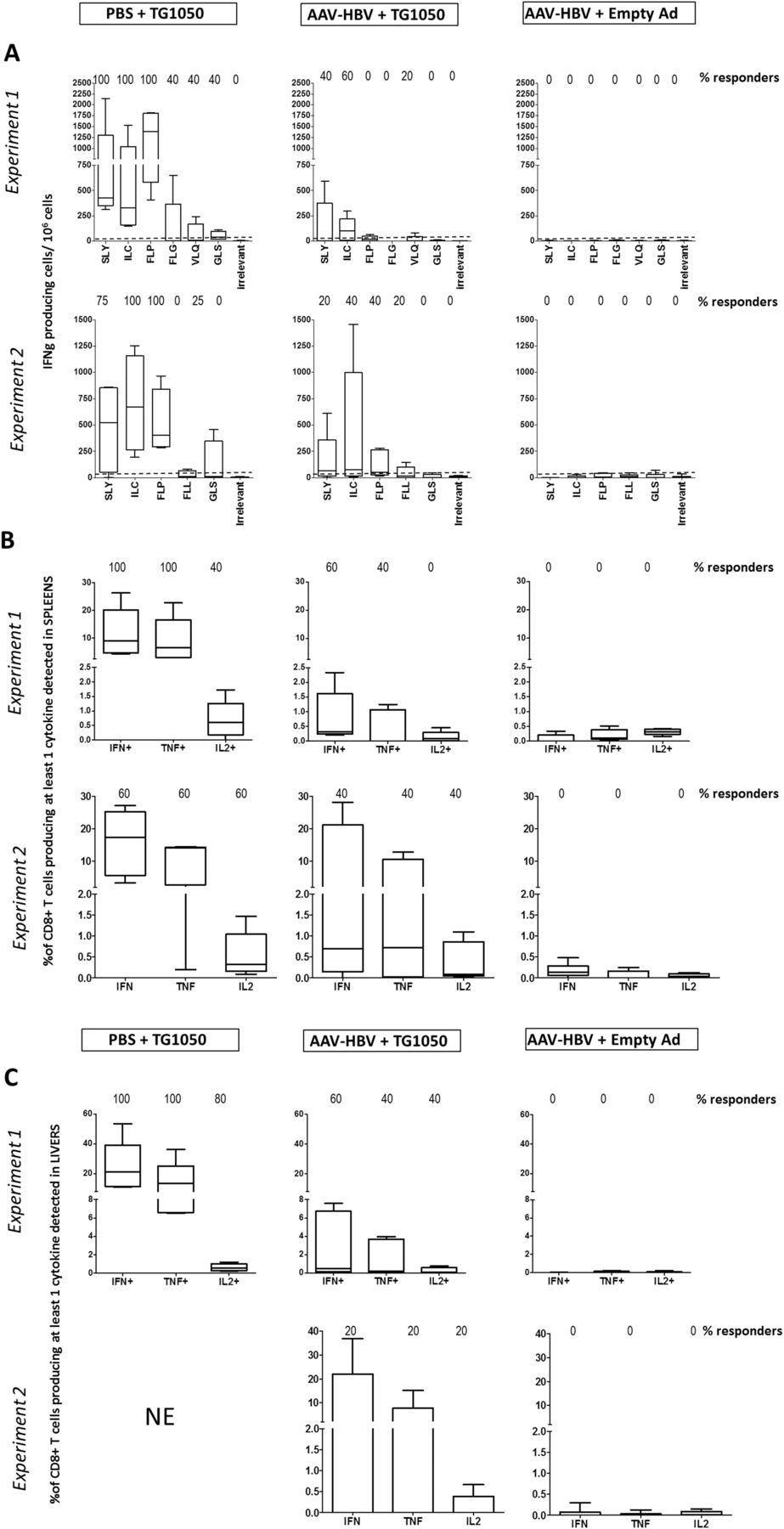
Two experiments were performed in order to closely assess the kinetics of appearance of TG1050-induced T cells. The ability of TG1050 to induce functional T cells producing cytokines was assessed 2 weeks after TG1050 immunisation. Analysis performed on spleen cells of AAV-HBV mice showed that TG1050 was able to induce IFNγ-producing cells specific for HLA-A2 epitopes located within Polymerase, Core and Env to a lower extent (figure 4A). The frequencies of IFNγ-producing cells and of responding mice were notably detectable but lower in these mice (median values up to 105 and 73 spots/106 splenocytes in experiments 1 (p<0.01) and 2 (p=NS), respectively), than in HBV-free mice (median values up to 1389 and 605 spots/106 splenocytes in experiments 1 and 2, respectively). Splenic CD8+ T cells producing at least one cytokine could be detected in 60% and 40% of AAV-injected mice with percentages of detection reaching 0.32% and 0.7% (experiments 1 and 2 respectively, figure 4B). In comparison, median values of CD8+ T cells producing at least one cytokine were up to 9.1% and 17.4% and found in up to 100% and 60% of HBV-free responding mice (experiments 1 and 2, respectively). In experiment 1, detected cells produced mainly one or two cytokines (mostly IFNγ+ or IFNγ+/TNFα+) in HBV-free and AAV-HBV injected mice (data not shown). Liver-infiltrating HBV-specific T cells were also monitored (figure 4). In AAV-HBV injected mice, cytokine-producing cells were detected in both experiments, albeit with distinct frequencies (percentage of responding mice ranged from 20% to 60%). The percentages of HBV-specific cells producing IFNγ and/or TNFα and/or IL-2 and the percentages of responding mice were lower in AAV-HBV mice than in HBV-free mice (21% vs 0.46% of CD8+ T cells producing at least one cytokine; experiment 1, median values). Some IL-2 producing cells were detected in the liver of AAV-HBV injected mice, hence the overall proportion of multiple cytokine producer cells (double and triple) approximated 80%, with a slightly lower proportion in HBV-free mice (data not shown). In experiment 2, due to the high background observed within the liver, no CD8+ T cells producing cytokines (single or multiple) could be detected in the control HBV-free mice. In AAV-HBV injected mice, one mouse injected with TG1050 displayed detectable CD8+ T cells producing at least one cytokine (figure 4C).

Analysis of short-term HBV-specific functional T cells in HLA-A2/DR1 AAV-HBV-persistent mice following a single TG1050 immunisation. HLA-A2/HLA-DR1 transgenic mice were intravenously injected with PBS or AAV-HBV (day 0) and immunised 1 month later (day 32) with either TG1050 or empty adenovirus. Induced HBV-specific immune responses were assessed 2 weeks after TG1050 or empty Ad immunisation through (A) IFNγ ELISPOT assay on spleen cells and interferon γ (IFNγ)/tumour necrosis factor α (TNFα) intracellular cytokine staining assays using (B) spleen cells and (C) liver-infiltrating lymphocytes. The analytical conditions are as described in figure 3, except that HLA-A2-restricted peptides were used (see online supplementary table S1). Data from two experiments are presented. In experiment 2, data from the group PBS+TG1050 could not be exploited due to high background. NE, non-evaluable.
As shown in figure 5, circulating HBV DNA remained stable (experiment 1) or increased (experiment 2) in the control groups that received an empty Ad vector (experiment 1, HBV DNA increased 0.7–1.7-fold from 6 to 14 days post-Empty Ad injection; experiment 2, HBV DNA increased 4-fold and 9-fold at 6 and 10 days post TG1050 injection, respectively; figure 5A). In contrast, TG1050-injected mice controlled HBV DNA increase from 6 to 10 or 14 days post-injection. A strong trend was observed in both experiments, and statistical significance was reached in experiment 2 at day 38 (p=0.0174). Initial levels of HBV DNA were different in both experiments, a factor that may have influenced the effect of TG1050.
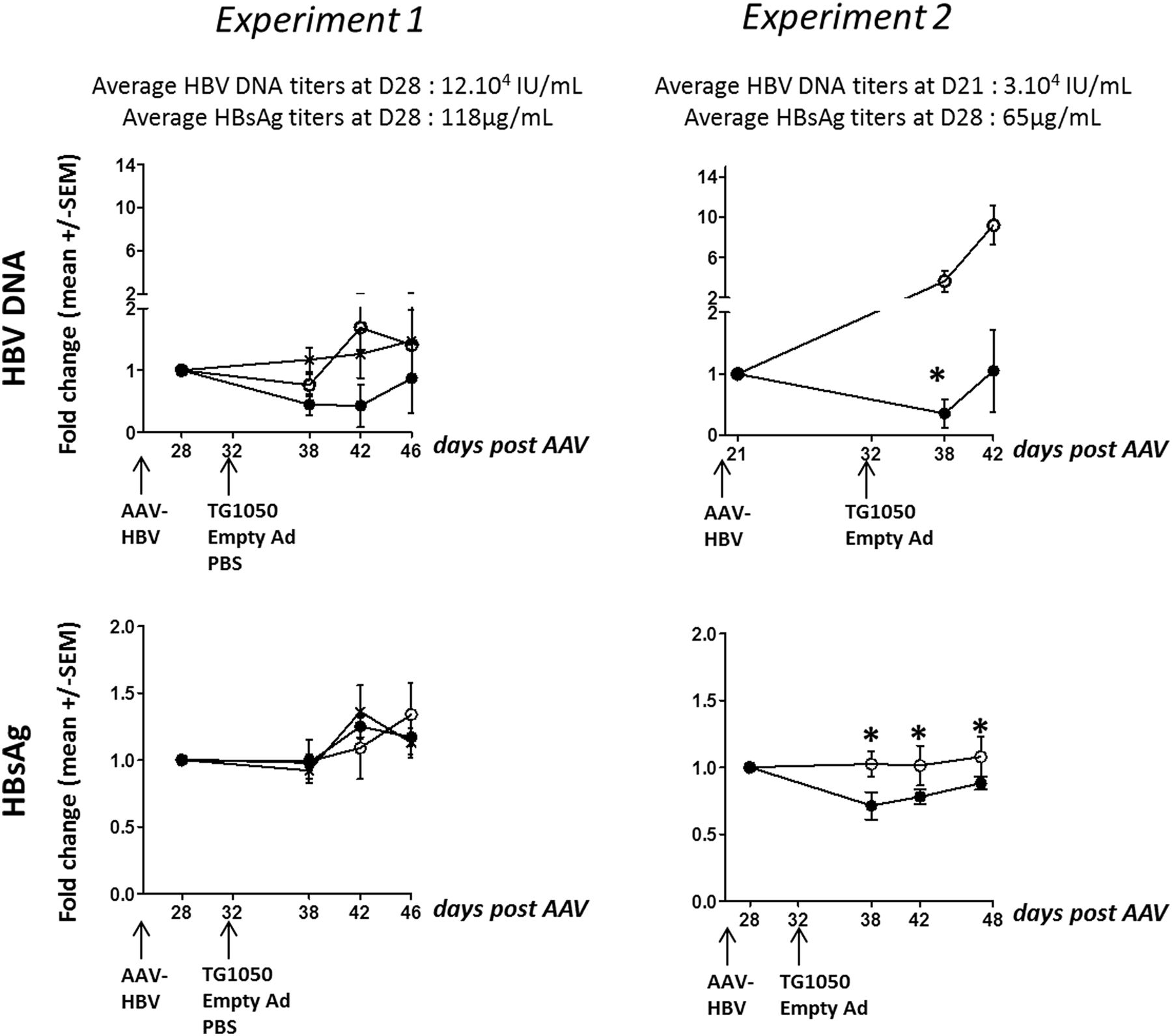
Monitoring of viral parameters in HLA-A2/DR1 AAV-HBV-persistent mice. HLA-A2/HLA-DR1 transgenic mice were intravenously injected with PBS or AAV-HBV (day 0) and immunised 1 month later (day 32) with either TG1050 or empty adenovirus. (A) HBV DNA and (B) HBsAg data are represented as mean fold changes (DNA and HBsAg) compared with their respective mean level at day 28 before TG1050 (plain circle curve) or empty Ad (empty circle curve) immunisation or in non-immunised mice (cross curve, for experiment 1 only). Statistical analyses were performed using mixed models and ANOVA for repeated measures. *p<0.05.
Injection of TG1050 did not affect HBsAg levels in experiment 1, although the levels were consistently lower. A statistically significant effect was seen in experiment 2 (figure 5B). In this experiment, a decrease was maintained over three consecutive time points, (p=0.0296, p=0.0319 and p=0.022 between AAV-HBV+Empty Ad and AAV-HBV+TG1050 groups for days 38, 42 and 48, respectively).
No increase in ALAT levels over normal values was detected in either experiment, independently of the analysed groups (data not shown).
Long-term monitoring
Monitoring of spleen cells up to 4 months after TG1050 injection (see online supplementary figure S4A, IFNγ ELISPOT assay) showed induction of high frequencies of IFNγ-producing cells targeting Core, Polymerase and Env domains. Frequencies of IFNγ+ cells and percentages of responding mice were comparable between TG1050 immunised HBV-free (PBS) and AAV-HBV mice. A high percentage of HBV-specific CD8+ T cells producing IFNγ and/or TNFα and/or IL-2 was detected in HBV-free and AAV-HBV mice (see online supplementary figure S4B). The percentages of cells producing one, two and three cytokines were almost identical between the two groups, whether tested in the spleen or the liver (see online supplementary figure S4C).
Viral parameters following TG1050 or empty-Ad administration were low in this experiment at the time of TG1050 administration (mean 7.11 µg/mL HBsAg and <300 IU/mL DNA in 50% of mice), probably due to the use of a lower dose of AAV-HBV injected compared with that used in the short-term experiments (5 times lower ie, 1×1010 vg). Hence, no impact on HBV DNA at day 47 or day160 could be seen (data not shown). Overall, comparison at all time points showed that HBsAg levels were consistently lower in AAV-HBV mice immunised with TG1050 than in mice receiving an empty Ad, and significantly different at day 47 (see online supplementary figure S5).
In conclusion, data collected in HLA-A2/DR1 mice show that TG1050 can educate functional T cells in a HBV-persistent environment and can impact on both HBV DNA and HBsAg levels, albeit in a transient manner.
TG1050 is able to induce functional T cells and sustained antiviral responses in HBV-persistent C57BL/6J mice following multiple administrations
The AAV-HBV-persistent model was set up in C57BL/6J mice and TG1050 was injected up to four times.
Mice were immunised once with TG1050 at day 32 post AAV-HBV injection; half of them were monitored for viral parameters and sacrificed for immune monitoring at day 46/47 (figure 6A left panel and C). The remaining mice were kept alive, injected an additional three times at 1-week intervals and followed for viral parameters up to day 105 (figure 6A right panel and B). A strong reduction in HBV-DNA titres was quickly observed (2 and 3 weeks after first TG1050 immunisation) compared with control mice (26-fold and 44-fold, figure 6A). Following additional injections, the HBV-DNA titre remained significantly reduced until the end of monitoring. HBsAg titres also decreased 2–3 weeks after first TG1050 immunisation and remained low up to day 105 (10 weeks after first immunisation) in mice receiving three additional injections of TG1050 compared with control mice.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Monitoring of viral and immunological parameters in C57BL/6J AAV-HBV-persistent mice. C57BL/6J naïve mice were intravenously injected with PBS or AAV-HBV (day 0) and immunised 1 month later (day 32) with either TG1050 or empty adenovirus. (A) HBV DNA and (B) HBsAg data are represented as mean fold changes (DNA and HBsAg) compared with their respective mean level before TG1050 (plain circle curve) or empty Ad (empty circle curve) immunisation. Statistical analyses were performed using mixed models and ANOVA for repeated measures. The analysis was done on changes from baseline in percentage. For DNA, the analysis was performed on ranks in order to fulfil the mixed model hypothesis. *p<0.05. Induced HBV-specific immune responses were assessed 2 weeks after TG1050 or empty Ad immunisation (day 47) with interferon γ (IFNγ)/tumour necrosis factor α (TNFα)/interleukin 2 (IL-2) intracellular cytokine staining assays using liver-infiltrating lymphocytes (C), using the H-2b restricted peptide VSA (polymerase) for in vitro stimulation. The results are presented as the percentage of CD8+ T cells producing at least IFNγ or TNFα or IL-2. The percentage of mice displaying a positive response is indicated at the top of the graph.
The percentages of HBV-specific cells producing IFNγ and/or TNFα and/or IL-2 were monitored 2 weeks after the first TG1050 injection and were found to be lower in HBV-persistent mice than in HBV-free mice (8.4% vs 1.9% of IFNγ+ CD8+ T cells, figure 6C). However, as a higher number of CD8+ T cells was observed in the liver of AAV-HBV mice (about three times more than in HBV-free mice), absolute numbers of HBV Polymerase-specific CD8T cells induced by TG1050 in the AAV-HBV and HBV-free mice were very close (data not shown). An IFNγ response was detected in all TG1050-injected mice, both HBV-free and persistent, whereas the percentage of HBV persistent mice displaying TNFα+ and/or IL-2+ CD8+ T cells was low (50% and 0%) compared with HBV-free mice (100% and 80%). The percentage of TG1050-induced cells secreting at least two cytokines was higher in HBV-free mice (45%) than in HBV-persistent mice (19%).
Overall, data collected in C57BL/6J HBV-persistent mice show that TG1050 induces a rapid and sustained antiviral effect following multiple injections.
Discussion
Adenovirus vectors are extremely potent at inducing cellular-based immune responses to the encoded immunogens and have been used to develop vaccines against a range of infectious diseases and cancers.28 Ad serotype 5 has been the most widely used Ad vector, displaying a good safety profile and a remarkable capacity to induce long-lasting and broad T cell-based immune responses, in particular those driven by CD8+.29 TG1050 is an Ad-5 vector expressing a unique fusion protein that includes the Core protein described as an effective target of T cell responses in HBV resolvers.30 ,31 We show here in mouse models that a single administration of TG1050 induces mainly CD8 T cells (2–5%), although HBV-specific CD4 T cells were also detected to a lesser extent (0.05–0.15%). While CD4 T cells are important in HBV control,32 CD8 T cells have been shown to be essential in clearance of HBV, as demonstrated in depletion studies performed in chimpanzees33 and in studies describing functional exhaustion of HBV-specific CD8 T cells in patients with CHB.34 The detection of multispecific T cells targeting all three TG1050-encoded immunogens and producing multiple cytokines was also observed. In addition, in vivo analyses demonstrated the induction of CTL, with significant cell lysis observed for Core- and Polymerase-specific peptides. CTL are expected to be a key component of the mechanism of action of TG1050 as they aim at killing infected hepatocytes and hence clearing cccDNA.35 ,36 Finally, our study shows induction of long-lasting HBV-specific T cells, detectable up to 400 days post-injection. Such induced Core-specific T cells are mainly effector memory cells (CD44+/CD62L−) and some of them displayed markers (CD27+/CD43−) associated with strong recall capacity but also central memory cells (CD44+/CD62L+) that are detected until day 400 post-injection. In contrast to other vaccine vectors, adenoviruses are known to induce a long-lasting effector phase, possibly due to the persistence of a low level of viral transcription in T lymphocytes37 together with a significant memory response.38
Although anti-adenovirus immune responses were not measured in this study, we have shown that a single subcutaneous administration of HBV-specific Ad vectors (unpublished information and Boukhebza et al23) expectedly results in both T and B Ad-specific immune responses that do not preclude the mounting of HBV-specific responses, including after multiple administrations.23 Pre-existing immunity to Ad5 in humans has been reported to hamper mounting of immunity to encoded immunogens, in particular in the HIV field39 and when E1-deleted Ad5 are used. However, recent papers on malaria and tuberculosis vaccine development have reported no impact of pre-Ad5 immunity to vaccine immunogens, particularly when E1/E3 deleted vectors are used, such as for TG1050.40 ,41 Strategies to formulate Ad5 in order to minimise and/or prevent the impact of anti-Ad5 neutralising antibodies on vaccine immunogenicity can be explored.42
First-generation HBV immunotherapeutics have been disappointing in the clinic. These were based on suboptimal vector platforms with respect to induction of strong T cell-based immunity. They have included peptides, adjuvanted recombinant antigens in single or complex mixtures including existing HBV prophylactic vaccines as well as DNA vaccines administered by conventional means.16 In contrast, TG1050 has a high antigenic complexity covering either full length or major domains of three HBV antigens and is based on an optimal platform for induction of CD8-driven responses. Closest to TG1050, HB-110 is a combination of three plasmid DNAs expressing pre-S/S, Core/Polymerase and the IL-12 cytokine which showed promising results in a therapy arrest clinical trial setting,43 although this was not confirmed in a later study.44
Among the few animal models supporting infection by a HBV,45 studies have been conducted in woodchucks infected by woodchuck hepatitis virus (WHV) including evaluation of combined use of NUC and immunotherapeutics, a potent combination of a DNA and an adenovirus-based vaccine.46 WHV and HBV genomic sequences display a high rate of diversity, ranging from 25% to 60%, limiting the potential of the woodchuck model in the evaluation of immunotherapeutics targeting the human virus. Here we used the recently described alternative HBV-persistent murine model (AAV-HBV)17 to evaluate the capacity of TG1050 to educate HBV-specific functional T cells in face of ongoing HBV antigen expression. We also provide a first insight into the capacity of TG1050 to exert an antiviral effect. Remarkably, in this model a single subcutaneous administration of TG1050 was able to educate broad HBV-specific CD8+ T cells targeting Core, Pol and Env and capable of producing IFNγ, TNFα and, in some cases, IL-2, including in the liver of immunised mice. In parallel, some control of HBV DNA and HBsAg levels in the serum of vaccinated mice was achieved, reaching statistical significance in two of the three reported short-term experiments, both in a HLA-A2/HLA-DR1 and in a C57BL/6J background. In a similar mouse model, Tay et al47 showed that the threshold of antigen expression within the liver is a dominant factor determining the fate of T cells, with a high percentage of cells being silenced. Interestingly, in C56BL/6J AAV-HBV mice, a single administration resulted in a dramatic reduction in both HBV DNA and HBsAg (>1 log decrease for the DNA), and a significant decrease was sustained until day 105. This persistent effect may be linked to the three additional closely-spaced boosting injections administered. HLA-A2/DR1 mice have been shown to harbour a low percentage CD8+ T cells (reaching only 2–3% of total splenocytes48), a feature that may have been suboptimal with respect to mounting of TG1050 antiviral efficacy. In humans a correlation has been reported between T cell exhaustion and viral load and/or antigenic load; the higher the levels of viral parameters carried by the patients, the more immune defects are observed.31 Together our observations suggest that multiple TG1050 administrations may have to be used in the clinic, particularly in patients with higher viral markers. Application of closely spaced administrations of viral-vectored immunotherapeutics, as performed here, has shown clinical efficacy in the treatment of patients with chronic hepatitis C.49 TG1050 is intended to be first developed in the clinic in combination with NUC, hence the ultimate goal will be elimination of HBsAg. At that stage, our preclinical data show the capacity of TG1050 to affect the levels of HBsAg in the absence of a significant elevation of alanine aminotransferase. In patients, long-term treatment with NUC has been shown in in vitro expansion studies to restore some T cell functionality.50 The combination of TG1050 with NUC and possibly with blockers of inhibitory molecules described in HBV T cells of patients with CHB such as PD-1, TIM-3 and SLAM,51 ,52 should be explored.
In conclusion, we have developed a novel active targeted immunotherapeutic that displays a number of immunological features found in HBV-infected resolvers together with the capacity to exert antiviral activity on a stand alone-based approach.
Acknowledgments
We thanks all employees of Plateau de Biologie Expérimentale de la souris in Lyon for taking care of the animals. This work was partially funded by NATHEB program (FEDER/FUI).
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
Contributors All authors listed have contributed to the work described in the article. CD, EJ, AE and HB conducted immunological experiments in HBV naïve mice. RK, KL-S and AE conducted experiments in C57BL6 AAV-HBV mice. PM and GI supervised the experiments and took a major part in writing the paper. SD, MM-B and OG conducted experiments in the HLA-A2/HLA-DR1 AAV-HBV model. M-LM supervised these experiments and made an important contribution to writing the paper. J-FM performed HBV DNA measurements. YS, DV, J-BM, MG, RB and AF performed in vitro analyses. TM, NS, J-BM supervised these and took part in writing of the manuscript. J-MS and AVD performed mass spectrometry analysis.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement Not relevant, no unpublishd data available.