Article Text

Abstract
Objective The short isoform of thymic stromal lymphopoietin (TSLP), a cytokine constitutively expressed by epithelial cells, is crucial in preserving immune tolerance in the gut. TSLP deficiency has been implicated in sustaining intestinal damage in Crohn's disease. We explored mucosal TSLP expression and function in refractory and uncomplicated coeliac disease (CD), a T-cell-mediated enteropathy induced by gluten in genetically susceptible individuals.
Design TSLP isoforms—long and short—and receptors—TSLPR and interleukin (IL)-7Rα—were assessed by immunofluorescence, immunoblotting and qRT-PCR in the duodenum of untreated, treated, potential and refractory patients with CD. The ability of the serine protease furin or CD biopsy supernatants to cleave TSLP was evaluated by immunoblotting. The production of interferon (IFN)-γ and IL-8 by untreated CD biopsies cultured ex vivo with TSLP isoforms was also assessed.
Results Mucosal TSLP, but not TSLPR and IL-7Rα, was reduced in untreated CD and refractory CD in comparison to treated CD, potential CD and controls. Transcripts of both TSLP isoforms were decreased in active CD mucosa. Furin, which was overexpressed in active CD biopsies, was able to cleave TSLP in vitro. Accordingly, refractory and untreated CD supernatants showed higher TSLP-degrading capacity in comparison to treated CD and control supernatants. In our ex vivo model, both TSLP isoforms significantly downregulated IFN-γ and IL-8 production by untreated CD biopsies.
Conclusions Reduced mucosal TSLP expression may contribute to intestinal damage in refractory and untreated CD. Further studies are needed to verify whether restoring TSLP might be therapeutically useful especially in refractory patients with CD.
- COELIAC DISEASE
- GUT IMMUNOLOGY
- MATRIX METALLOPROTEINASE
- MUCOSAL IMMUNITY
- SMALL INTESTINE
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Epithelial-derived thymic stromal lymphopoietin (TSLP) is an immunoregulatory cytokine playing a crucial role in immune tolerance in the gut via conditioning dendritic cells to expand regulatory T cells (Treg), with consequent inhibition of T-cell-derived cytokines, such as interferon (IFN)-γ and interleukin (IL)-17A.
Mucosal TSLP is reduced in Crohn's disease, a chronic intestinal disorder characterised by an abnormal T helper (Th)1/Th17 response, and this defect contributes to gut inflammation.
In coeliac disease (CD), a chronic enteropathy caused in genetically susceptible individuals by the ingestion of gluten, villous atrophy is induced by a T-cell-derived cytokine upregulation, abnormal dendritic cell activation and increased matrix-metalloproteinase (MMP) production.
What are the new findings?
In addition to the marked reduction in the transcript levels of both long and short TSLP, TSLP protein levels are markedly decreased in the duodenal mucosa of untreated and refractory patients with CD.
The serin protease furin, which is overexpressed in active CD mucosa, potently degrades long TSLP.
Short TSLP modulates inflammation in CD mucosa, as shown by the ex vivo effect in inhibiting IFN-γ and IL-8 production by untreated CD biopsies.
Untreated CD myofibroblasts may be the target of TSLP action, as proven by their expression of TSLPR and IL-7 receptor (IL-7R)α, and respond to long TSLP by producing lower amounts of MMP-3 and IL-8.
How might it impact on clinical practice in the foreseeable future?
Our findings strengthen the key role of TSLP in modulating immune and damaging mechanisms in the inflamed gut, and may open new therapeutic perspectives in patients with CD aimed at re-inducing immune tolerance towards gluten via restoring TSLP function especially in refractory patients with CD, who no longer respond to a gluten-free diet.
Introduction
Thymic stromal lymphopoietin (TSLP) is a cytokine produced mainly by epithelial cells and expressed in the skin, lungs, thymus and intestinal mucosa.1 TSLP exerts its biological activities by binding to a heterodimer formed by interleukin (IL)-7 receptor (IL-7R)α and TSLP receptor (TSLPR), leading to the phosphorylation and activation of signal transducer and activator of transcription (STAT)5.1–4 During homeostasis, enterocyte-derived TSLP promotes the development of tolerogenic dendritic cells,5 which in turn induce the differentiation of naive T cells into Foxp3-positive regulatory T cells (Treg) and block the development of T helper (Th)1 and Th17 cells.6 ,7 TSLP expression and function are impaired in chronic intestinal inflammation, as shown by the observations that epithelial TSLP expression is markedly reduced in inflamed Crohn's disease mucosa, and that normal dendritic cells cultured with supernatants of Crohn's disease epithelial cells show only a limited ability to induce Treg development.5 Recently, two human transcript variants of TSLP have been identified, namely the long and short TSLP isoforms.8 ,9 Short TSLP is the main TSLP isoform expressed under steady-state conditions and has anti-inflammatory and antimicrobial properties.8 ,9
Coeliac disease (CD) is an enteropathy caused in genetically susceptible individuals by the ingestion of gluten.10 Rather than a single-disease entity, CD is considered a polymorphic spectrum of disorders ranging from potential CD, referring to patients with normal small bowel mucosa and positive coeliac serology, to refractory CD, a rare condition characterised by the persistence of villous atrophy despite strict adherence to gluten-free diet (GFD).10 ,11 The abnormal immune response in active CD mucosa is sustained by the aberrant activation and differentiation of naive T cells into Th1 and Th17 cells, which respectively release high amounts of interferon (IFN)-γ and IL-17A,12 ,13 finally causing, via upregulating the intraepithelial lymphocyte cytotoxicity against enterocytes14 and the production of tissue-damaging matrix metalloproteinases (MMPs),15 increased enterocyte apoptosis and subsequent villous atrophy.10 ,16
The presence of a marked T-cell-derived cytokine upregulation in active CD mucosa,17 together with the increased dendritic cell infiltration, activation and maturation observed in the lamina propria of untreated patients with CD,18 led us to hypothesise that a dysregulation of TSLP expression could be implicated in this condition. On this basis, we explored TSLP expression along the multifaceted clinical spectrum of CD, and then functionally tested the ex vivo and in vitro effects of TSLP isoforms on the immune response and the extracellular matrix remodelling processes in CD mucosa.
Materials and methods
Patients and tissues
Well-oriented endoscopic biopsies were collected from the second part of the duodenum of 64 patients with untreated CD (mean age 35.7 years, range 19–66) and 50 patients with CD after at least 12 months of GFD (mean age 42.3 years, range 21–70). Diagnosis was based on the positivity of serum anti-endomysial and anti-tissue transglutaminase antibodies associated with typical histopathological lesions, namely villous atrophy, increased intraepithelial lymphocyte infiltration and crypt hyperplasia.10 Among the 64 untreated patients with CD, 59 showed a B2 lesion and 5 showed a B1 lesion, according to Corazza-Villanacci grading.19 Histological improvement was documented in all treated patients with CD. Duodenal biopsies were also obtained from eight patients with refractory CD (three type 1 and five type 211; mean age 62.8 years, range 38–76) (table 1), four patients with potential CD (mean age 47.2 years, range 31–71) (table 2) and 43 control subjects (mean age 43.6 years, range 19–73) undergoing endoscopy for functional dyspepsia, negative for anti-endomysial and anti-tissue transglutaminase antibodies and with normal histology. Potential CD was diagnosed in the presence of an architecturally normal small bowel mucosa despite persistent positivity of serum coeliac antibodies.10 All potential patients with CD were on a gluten-containing diet at the time of the biopsy, and they are currently under a yearly endoscopic follow-up. Some of the biopsies were processed for routine histology or embedded in OCT Tissue-Tek (Sakura Finetek, Torrance, California, USA) and snap frozen; others were used for organ culture or cell isolation or homogenised for immunoblotting. Heparinised peripheral venous blood of six healthy volunteers (mean age 38.6 years, range 30–46) was collected.
Clinical and pathological features of eight patients affected by refractory coeliac disease (RCD)
Clinical and pathological features of four patients affected by potential coeliac disease (PCD)
Immunofluorescence
Four 5-μm-cryostat sections were fixed in cold acetone or in paraformaldehyde 4% for 20 min and incubated overnight at 4°C with rabbit anti-TSLP (3 μg/mL; manufactured and kindly provided by Maria Rescigno's lab; please see online supplementary methods for details) or rabbit anti-TSLPR antibodies (1:50 dilution; Santa Cruz Biotechnology, Santa Cruz, California, USA). Cy3-conjugated donkey anti-rabbit (1:400 dilution; Jackson, West Grove, Pennsylvania, USA) and FITC-conjugated goat anti-rabbit (1:500 dilution; Abcam, Cambridge, UK) were used as secondary antibodies. Sections were counterstained by 4′,6-diamidino-2-phenylindole (1:1000 dilution; Life Technologies, Paisley, UK), and analysed with a laser scanning confocal microscope (FluoView FV1000; Olympus, Center Valley, Pennsylvania, USA). Images (1024×1024 pixels) were acquired with an oil immersion lens (60×1.4 NA Plan-Apochromat; Olympus).
Counting of epithelial cells
For cell counts, 8–24 crypts were evaluated only if aligned along the longitudinal axis such that the lumen of the crypt could be seen along its length. Observation of epithelial cells was performed using conventional light microscopy in a blinded manner by the same expert observer. Counts were performed at a constant magnification (20×) by counting with an ocular linear graticule the number of epithelial cells placed in the epithelial layer overlying a fixed unit length (200 μm) of muscularis mucosae, defined by the length of the graticule (60 μm).
RNA extraction and analysis of mRNA expression by quantitative RT-PCR
Reverse transcription (Im-PromII, Promega, Southampton, UK) on 1 μg total RNA extracted from biopsies was performed, and cDNA was used for PCR. Primer sequences were as follows: TSLP forward, 5′-CCCAGGCTATTCGGAAACTCAG-3′, and reverse, 5′-CGCCACAATCCTTGTAATTGTG-3′ (these primers do not distinguish between the two TSLP isoforms); long TSLP forward, 5′-CACCGTCTCTTGTAGCAATCG-3′, and reverse, 5′-TAGCCTGGGCACCAGATAGC-3′; short TSLP forward, 5′-CCGCCTATGAGCAGCCAC-3′, and reverse, 5′-CCTGAGTAGCATTTATCTGAG-3′; TSLPR forward, 5′-AGAGCAGCGAGACGACATTC-3′, and reverse, 5′-CCGGTACTGAACCTCATAGAGG-3′. Typically, 40 cycles of 20 s at 95°C and 20 s at 60°C, followed by the thermal dissociation protocol for Fast SYBR green detection. PCR reactions were normalised by expression analysis of GAPDH or cytokeratin 18 (CK18) with the following primers: GAPDH forward, 5′-ATCAGCAATGCCTCCTGCAC-3′, and reverse, 5′-TGGCATGGACTGTGGTCATG-3′; CK18 forward, 5′-TGATGACACCAATATCACACGAC-3′, and reverse, 5′-TACCTCCACGGTCAACCCA-3′.20
Organ culture
Biopsies were placed in 24-well plates (VWR International, Lutterworth, UK) in 300 μL serum-free HL-1 medium (Cambrex BioScience, Wokingham, UK) supplemented with 100 U/mL to 100 μg/mL penicillin-streptomycin solution (Life Technologies), and cultured at 37°C, 5% CO2, with or without 50 ng/mL long TSLP (aa 29–159, mw 15 kDa, Uniprot: Q969D9; R&D Systems, Abingdon, UK) or 50 ng/mL short TSLP (manufactured and kindly provided by Maria Rescigno's lab; please see online supplementary methods for details). After 24 h culture, biopsies and supernatants were stored at −70°C.
Lamina propria mononuclear cell isolation
Lamina propria mononuclear cells (LPMCs) were isolated from duodenal biopsies, as previously described.18 Cells were not used if viability did not exceed 90%.
ELISPOT
The 96-well polyvinylidene difluoride-backed ELISPOT plates (Millipore, Bedford, Massachusetts, USA) were coated with 15 μg/mL anti-IFN-γ antibody (Mabtech, Stockholm, Sweden) and incubated overnight at 4°C. LPMCs resuspended in AIM-V medium (Life Technologies) were added (2×105 cells/well, in triplicate) to the pre-coated plates with or without 50 ng/mL long TSLP (R&D Systems) in the absence or presence of soluble anti-CD3 (Mabtech) and anti-CD28 (eBioscience, San Diego, California, USA) antibodies, both at 1 μg/mL. Plates were incubated overnight at 37°C, 5% CO2. Next, 1 μg/mL biotinylated anti-IFN-γ antibody (50 μL/well; Mabtech) was added. After 2 h, streptavidin alkaline phosphatase conjugate (1:1000 dilution; Mabtech) was added to the wells and plates were incubated at room temperature for 2 h. Next, 100 μL/well of chromogenic alkaline phosphatase substrate (1:25 dilution; Bio-Rad Laboratories, Hercules, California, USA) was added. The colorimetric reaction was stopped after 30 min with water, then plates were air-dried. Spot-forming cells were counted using an ELISPOT counter (Millipore).
Myofibroblast isolation and culture
Myofibroblasts were isolated from biopsies as previously described21 and were used at passage 4–6. Subconfluent monolayers of myofibroblasts seeded in 12-well plates (1×105 cells/well) were starved for 24 h at 37°C, 5% CO2 before incubation for 24 h at 37°C, 5% CO2 with 50 ng/mL long TSLP (R&D Systems).
ELISA
IFN-γ and IL-8 in culture supernatants were measured using ELISA kits (R&D Systems).
TSLP cleavage assay
Long TSLP (1 μg/mL; R&D Systems) was co-incubated for 24 h at 37°C, 5% CO2 with 0.2–200 U/mL furin (Sigma-Aldrich, Poole, UK), or with complete RPMI-1640 (Sigma-Aldrich) or serum-free HL-1 (Cambrex BioScience) medium, or with supernatants of biopsies cultured ex vivo for 24 h. Effects on TSLP integrity and function were evaluated in cleavage reaction products by immunoblotting for TSLP and by bidirectional mixed lymphocyte reaction (see online supplementary methods for details), respectively.
Immunoblotting and immunoprecipitation
Immunoblotting was performed according to a modified method described previously.22 Proteins (100 μg) or cell supernatants (15 μL) were loaded and subjected to 10% SDS-PAGE under reducing conditions, followed by nitrocellulose (Bio-Rad Laboratories) transfer. The following anti-human primary antibodies were used: rabbit polyclonal anti-TSLP (0.2 μg/mL; Abcam), mouse anti-TSLPR (1:200 dilution; Santa Cruz Biotechnology), rabbit anti-IL-7Rα (1:1000 dilution; Abcam), rabbit anti-phospho-STAT5 (1:1000 dilution; Cell Signaling Technology, Danvers, Massachusetts, USA), mouse anti-T-bet (1:800 dilution; Abcam), rabbit anti-MMP-3 (1:500 dilution; Abcam), rabbit anti-MMP-9 (1:1000 dilution; Abcam) and rabbit anti-tissue inhibitor of matrix metalloproteinase (TIMP)-1 (dilution 1:1000; Abcam). Appropriate horseradish peroxidase-conjugated antibodies (DAKO, High Wycombe, UK) were used as secondary antibodies, and the reaction was developed with the ECL plus kit (Amersham Biosciences, Little Chalfont, UK). When required, blots were stripped and analysed for internal loading controls using rabbit anti-STAT5 (1:1000 dilution; Cell Signaling Technology) and rabbit anti-β-actin antibodies (1:5000 dilution; Abcam). Bands were quantified using an LKB Ultrascan XL Laser Densitometer (Kodak, Hemel Hempstead, UK). For immunoprecipitation experiments, total proteins were immunoprecipitated with a rabbit polyclonal anti-TSLP (0.2 μg/mL; Abcam), then analysed by immunoblotting for TSLP.
Statistical analysis
Data were analysed in the GraphPad Prism statistical PC program (GraphPad Software, San Diego, California, USA) using the non-parametric Mann–Whitney U (for comparison of the in vivo data obtained in different groups of patients) or the paired t test (for comparison of the ex vivo and in vitro data obtained in different culture conditions). p<0.05 was considered statistically significant.
Results
In vivo mucosal TSLP expression
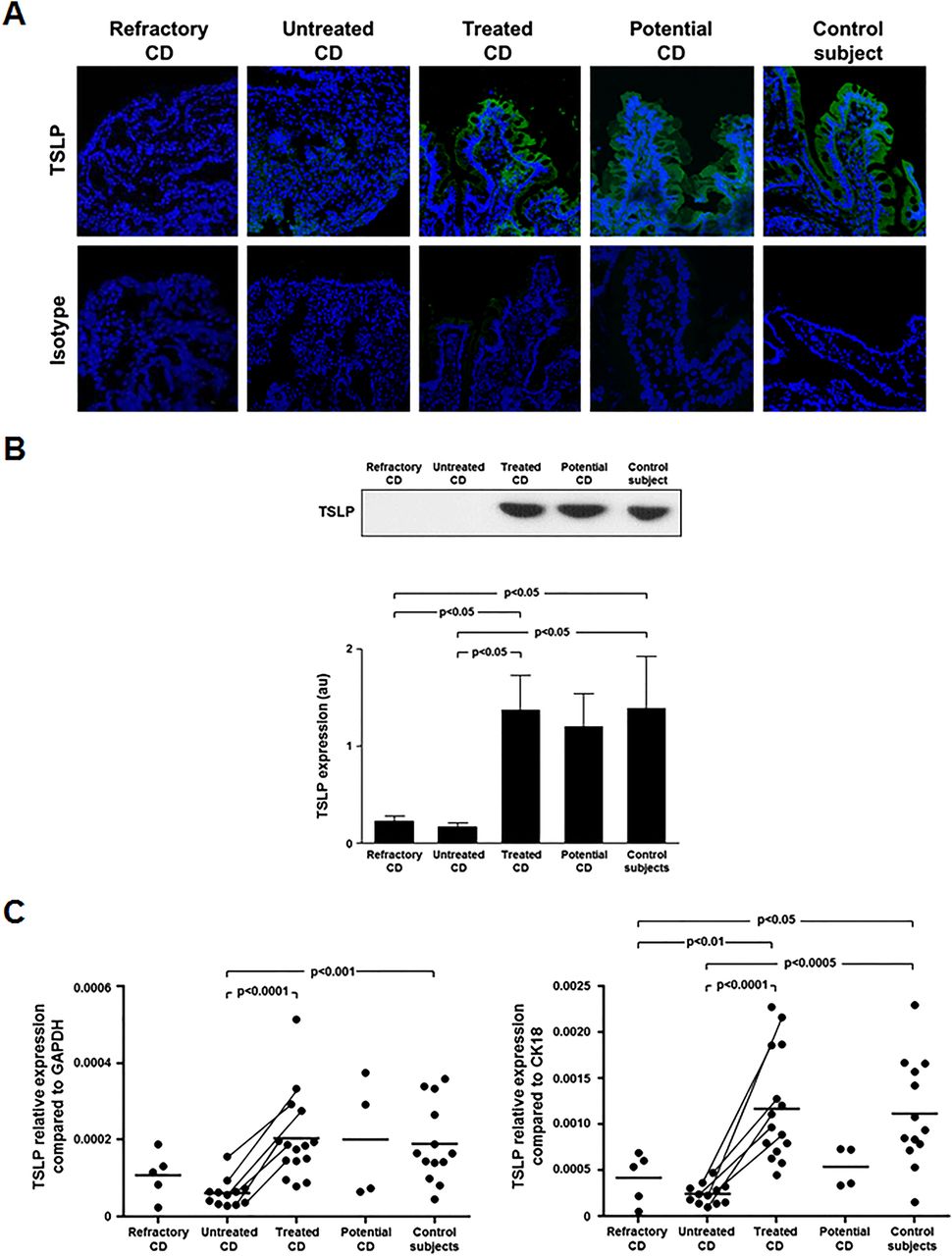
We first determined duodenal TSLP expression by immunofluorescence in untreated, treated, refractory, potential patients with CD and controls (figure 1A). TSLP was absent in the epithelium and lamina propria of untreated and refractory CD. Conversely, in treated CD, potential CD and controls the majority of enterocytes both at the epithelial surface and crypt level expressed TSLP. Several TSLP-positive LPMCs were also found in treated CD, potential CD and control mucosa. This is consistent with the capacity of dendritic cells to produce TSLP.23 ,24 No difference in TSLP expression was observed between treated CD, potential CD and controls. We performed epithelial cell counts per unit length of muscularis mucosae, and we observed a significantly (p<0.05) reduced number of epithelial cells in refractory (median 214, range 195–234) and untreated CD (median 215, range 187–247) compared with treated CD (median 246, range 205–266) and controls (median 247, range 221–274). Immunoprecipitation confirmed immunofluorescence data. A significantly (p<0.05) lower TSLP protein expression was observed in untreated and refractory CD compared with treated CD and controls. TSLP protein did not differ between untreated and refractory CD, and between treated CD and controls (figure 1B). We did not observe any difference in TSLP expression between type 1 and type 2 refractory CD. We also evaluated mucosal TSLP transcripts (total: short+long TSLP isoforms) by qRT-PCR (figure 1C). Upon normalisation for GAPDH, TSLP mRNA was significantly reduced in untreated CD compared with treated CD (p<0.0001) and controls (p<0.001). No significant difference was found in TSLP transcripts between refractory CD and all the other groups. In order to neutralise the possible bias due to the marked enterocyte loss occurring in active CD, we normalised TSLP transcripts for the epithelial cell marker CK18 (figure 1C). This normalisation confirmed the statistically significant reduction of TSLP transcripts in untreated CD compared with treated CD (p<0.0001) and controls (p<0.0005), and showed a significantly lower amount of TSLP mRNA in refractory CD compared with treated CD (p<0.01) and controls (p<0.05). The decrease in TSLP expression in six patients with CD evaluated before and after GFD supports the concept that gluten withdrawal normalises TSLP mucosal levels. We also measured transcripts of long and short isoforms of TSLP in the duodenal mucosa of untreated, treated CD and controls, and found that both the variants are defective in active CD mucosa (see online supplementary figure S1).

Reduced thymic stromal lymphopoietin (TSLP) expression in untreated and refractory coeliac disease (CD) mucosa. (A) TSLP expression was evaluated by immunofluorescence in the duodenum of a refractory, an untreated, a treated, a potential patient with CD and a control subject. While in untreated and refractory CD mucosa TSLP expression was absent, in treated and potential CD and controls TSLP was expressed both at epithelial and lamina propria level. Data are representative of staining performed in five refractory (two type 1 and three type 2), eight untreated, eight treated, three potential patients with CD and eight controls. An isotype antibody was used as negative control. (B) TSLP protein, analysed by immunoprecipitation in the duodenum of 5 refractory (two type 1 and three type 2), 16 untreated, 14 treated, 4 potential patients with CD and 14 controls. Examples in the upper panel are representative of all patients. Lower panel shows immunoblot densitometry. Results are mean±SEM. (C) TSLP transcripts (total: short+long TSLP isoforms), quantified by qRT-PCR, in the duodenum of 5 refractory (2 type 1 and 3 type 2), 12 untreated, 15 treated, 4 potential patients with CD and 13 controls. Changes in transcript levels are normalised for either GAPDH or cytokeratin 18 (CK18). Horizontal bars are mean values. au, arbitrary units.
In vivo mucosal furin expression and ex vivo TSLP cleavage
Protease upregulation is common in active CD mucosa.15 ,25 Furin is a ubiquitously expressed serine endopeptidase that has been shown to cleave TSLP.26–28 Thus, we evaluated furin expression by immunoblotting in mucosal homogenates (figure 2A). A significantly (p<0.05) higher furin expression was found in untreated CD compared with treated CD and controls. Subsequently, we assessed the effect of biopsy supernatants from refractory, untreated, treated patients with CD and controls on TSLP integrity (figure 2B). Furin degraded TSLP generating ∼10 and ∼3 kDa fragments. TSLP degradation by furin was concentration-dependent, as displayed by the increased intensity of the 10 kDa fragment and the parallel decrease in intensity of the intact TSLP band. Accordingly, at 24 h the band corresponding to intact TSLP was lower after co-incubation with refractory and untreated CD supernatants than with treated CD and control supernatants. A higher intensity of the cleaved TSLP band after 2 h was observed in refractory and untreated CD.
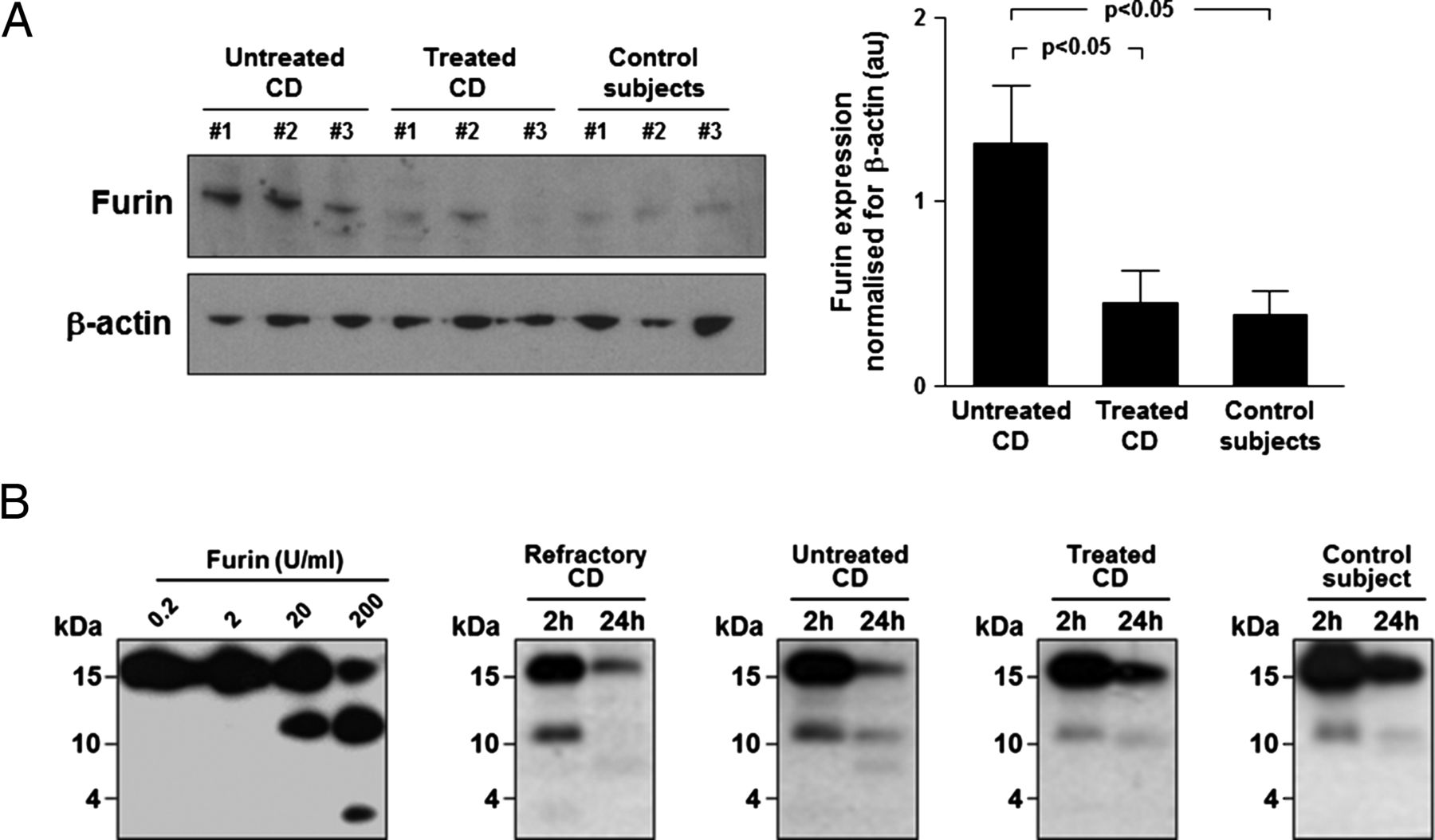
Furin overexpression and increased thymic stromal lymphopoietin (TSLP) degradation in untreated coeliac disease (CD) mucosa. (A) Furin protein was evaluated by immunoblotting in the duodenum of seven untreated and seven treated patients with CD, and seven controls. Examples in the left panel are representative of all patients. Blots were stripped and analysed for β-actin. Right panel shows immunoblot densitometry. Results are mean±SEM. (B) Effects of either recombinant human furin or supernatants of biopsies from three refractory, five untreated and five treated patients with CD and four controls on TSLP integrity. 1 μg/mL recombinant human TSLP was co-incubated with either furin (0.2–200 U/mL) or biopsy supernatants, then TSLP integrity was evaluated by immunoblotting on reaction products. Furin-mediated TSLP cleavage occurred in a concentration-dependent manner and was mirrored by decreasing intensity of the band corresponding to intact TSLP. At the highest furin concentration, a further ∼3 kDa TSLP fragment appeared. At 24 h, the band corresponding to intact TSLP is lower after co-incubation with refractory and untreated CD supernatants than with treated CD and control supernatants. In parallel, we observed a higher intensity of the band corresponding to cleaved TSLP after 2 h in refractory and untreated CD. Examples shown are representative of all patients. au, arbitrary units.
In vivo mucosal TSLPR and IL-7Rα expression
We then evaluated mucosal TSLPR expression by immunofluorescence (figure 3A). Some cells in the lamina propria were TSLPR-positive without differences among all the patient groups. These data were confirmed by TSLPR immunoblotting on mucosal homogenates. To better understand the cellular target of TSLP, we assessed the co-localisation of TSLPR with dendritic cell/macrophage (CD11c, CD14) and T cell (CD3) markers in untreated CD mucosa (see online supplementary figure S2). TSLPR was expressed mainly by enterocytes, whereas it was undetectable on CD3-positive intraepithelial lymphocytes and LPMCs. We also found that some CD11c/TSLPR-positive LPMCs were evident just underneath the epithelium. Moreover, a few mononuclear cells co-expressed TSLPR and CD14 in the lamina propria. IL-7Rα was expressed in the duodenal mucosa of all the patient groups without differences among the different conditions (figure 3B). TSLPR transcripts were not significantly different between all the patient groups, either upon normalisation for GAPDH or CK18, and TSLPR expression did not change before and after GFD in the six patients evaluated both at diagnosis and after GFD (figure 3C). TSLPR mRNA and protein expression did not differ between type 1 and type 2 refractory CD.

Thymic stromal lymphopoietin receptor (TSLPR) and interleukin (IL)-7Rα expression in coeliac disease (CD) mucosa. (A) Mucosal TSLPR, evaluated by immunofluorescence, in the duodenum of a refractory, an untreated, a treated, a potential patient with CD and a control subject. No difference was found in TSLPR expression both at epithelial and lamina propria level in all groups studied. Data are representative of staining performed in five refractory (two type 1 and three type 2), eight untreated, eight treated, three potential patients with CD and eight controls. An isotype antibody was used as negative control. (B) TSLPR and IL-7Rα protein, evaluated by immunoblotting, in the duodenum of 5 refractory (2 type 1 and 3 type 2), 16 untreated, 14 treated, 4 potential patients with CD and 14 controls. Examples in the upper panel are representative of all patients. Blots were stripped and analysed for β-actin. Lower panels show immunoblot densitometry. Results are mean±SEM. (C) TSLPR transcripts, quantified by qRT-PCR, in the duodenum of 5 refractory (2 type 1 and 3 type 2), 12 untreated, 15 treated, 4 potential patients with CD and 13 controls. Changes in TSLPR transcripts are shown after normalisation with either GAPDH or cytokeratin 18 (CK18). Horizontal bars are mean values. au, arbitrary units.
Ex vivo effects of TSLP on pSTAT5, T-bet and cytokines
Since TSLP exerts its function by STAT5 phosphorylation,1 we evaluated whether TSLP was functional in our system by measuring STAT5 phosphorylation in untreated CD biopsies cultured ex vivo with long or short TSLP (figure 4A). Long TSLP significantly (p<0.05) enhanced STAT5 phosphorylation ex vivo, whereas short TSLP did not have any effect on STAT5 phosphorylation. Since TSLP inhibits Th1 cell development,5 we evaluated the expression of the Th1-specific transcription factor T-bet on biopsies from untreated patients with CD cultured ex vivo with long or short TSLP (figure 4B). Both TSLP isoforms significantly (p<0.05) downregulated T-bet expression. Compared with medium alone (mean 215.3±56.6 pg/mL), both long and short TSLP significantly (p<0.05) downregulated IFN-γ production by untreated CD biopsies cultured ex vivo (mean 61.2±19.6 pg/mL and 70.3±23.8 pg/mL, respectively) (figure 4C). Long and short TSLP induced a significant (p<0.05) downregulation of IL-8 (mean 4052±1444 pg/mL and 4408±1250 pg/mL, respectively) compared with unstimulated conditions (mean 9829±1442 pg/mL) (figure 4C). Additionally, IL-8 was significantly decreased in culture supernatants of treated CD (mean 3227±1133 pg/mL, p<0.005) and controls (mean 4383±1155 pg/mL, p<0.05) compared with untreated CD (see online supplementary figure S3A).

Ex vivo effects of long and short thymic stromal lymphopoietin (TSLP) on cytokine production by untreated coeliac disease (CD) mucosa. Recombinant human long or short TSLP were cultured with biopsies from six untreated patients with CD and after 24 h culture phospho-signal transducer and activator of transcription (pSTAT)5 (A) and T-bet expression (B) was evaluated in mucosal homogenates by immunoblotting, while interferon (IFN)-γ and interleukin (IL)-8 production (C) was evaluated on biopsy supernatants by ELISA. Blots for pSTAT5 and T-bet were stripped and respectively analysed for STAT5 and β-actin. Examples shown are representative of all patients. Right panel shows immunoblot densitometry. Results are mean±SEM. Cytokine levels are mean±SEM. au, arbitrary units.
In vitro effects of TSLP on IFN-γ release by LPMCs and peripheral blood mononuclear cells
We then addressed whether LPMCs are target of TSLP. Untreated CD LPMCs express both TSLPR and IL-7Rα (figure 5A). We then cultured untreated CD LPMCs in ELISPOT plates with long TSLP in the presence of anti-CD3/CD28 antibodies and evaluated IFN-γ spot-forming cells. Long TSLP significantly (p<0.05) reduced IFN-γ spot-forming cells (mean 187.6±55.1/106) compared with medium (mean 457.8±100.6/106). Anti-CD3/CD28 induced a significant increase in mean IFN-γ spot-forming cells (827.8±89.8/106) compared with medium (p<0.05) and long TSLP (p<0.0005), and the addition of long TSLP significantly (p<0.05) downregulated IFN-γ spot-forming cells (mean 390.2±120.1/106) compared with culture with anti-CD3/CD28 antibodies (figure 5B). To explore the effects of furin-exposed long TSLP on IFN-γ production, we co-cultured peripheral blood mononuclear cells in bidirectional mixed lymphocyte reactions. Long TSLP, at the concentration of 100 and 200 ng/mL, induced a significant (p<0.05) upregulation of IFN-γ production, whereas furin-exposed long TSLP, at the concentration of 25 and 50 ng/mL, significantly (p<0.05) downregulated IFN-γ production compared with medium alone. Furin alone did not have any effect on IFN-γ release. As expected, vitamin D3 induced a dose-dependent reduction in IFN-γ concentration in culture supernatants. Short TSLP significantly (p<0.05) reduced IFN-γ production (see online supplementary figure S4).

In vitro down-regulatory effects of long thymic stromal lymphopoietin (TSLP) on interferon (IFN)-γ spot-forming untreated coeliac disease (CD) lamina propria mononuclear cells (LPMCs). (A) TSLP receptor (TSLPR) and interleukin (IL)-7Rα protein, detected by immunoblotting, on lysates of LPMCs isolated from two untreated patients with CD. Blots were stripped and analysed for β-actin. (B) Proportion of IFN-γ-secreting LPMCs from the duodenum of five untreated patients with CD evaluated through ELISPOT assay after in vitro stimulation with recombinant human TSLP. The photographs displayed in the upper panels, showing IFN-γ ELISPOTs derived from LPMCs unstimulated or stimulated with TSLP in the absence or presence of anti-CD3/CD28 antibodies (αCD3/CD28), are representative of all experiments performed. The frequency of IFN-γ-secreting LPMCs is shown in the lower panel as the mean number of IFN-γ spot-forming cells (SFCs) per 106 LPMCs corrected for background. Values are mean±SEM.
Ex vivo and in vitro effects of TSLP on MMP-3, MMP-9 and TIMP-1
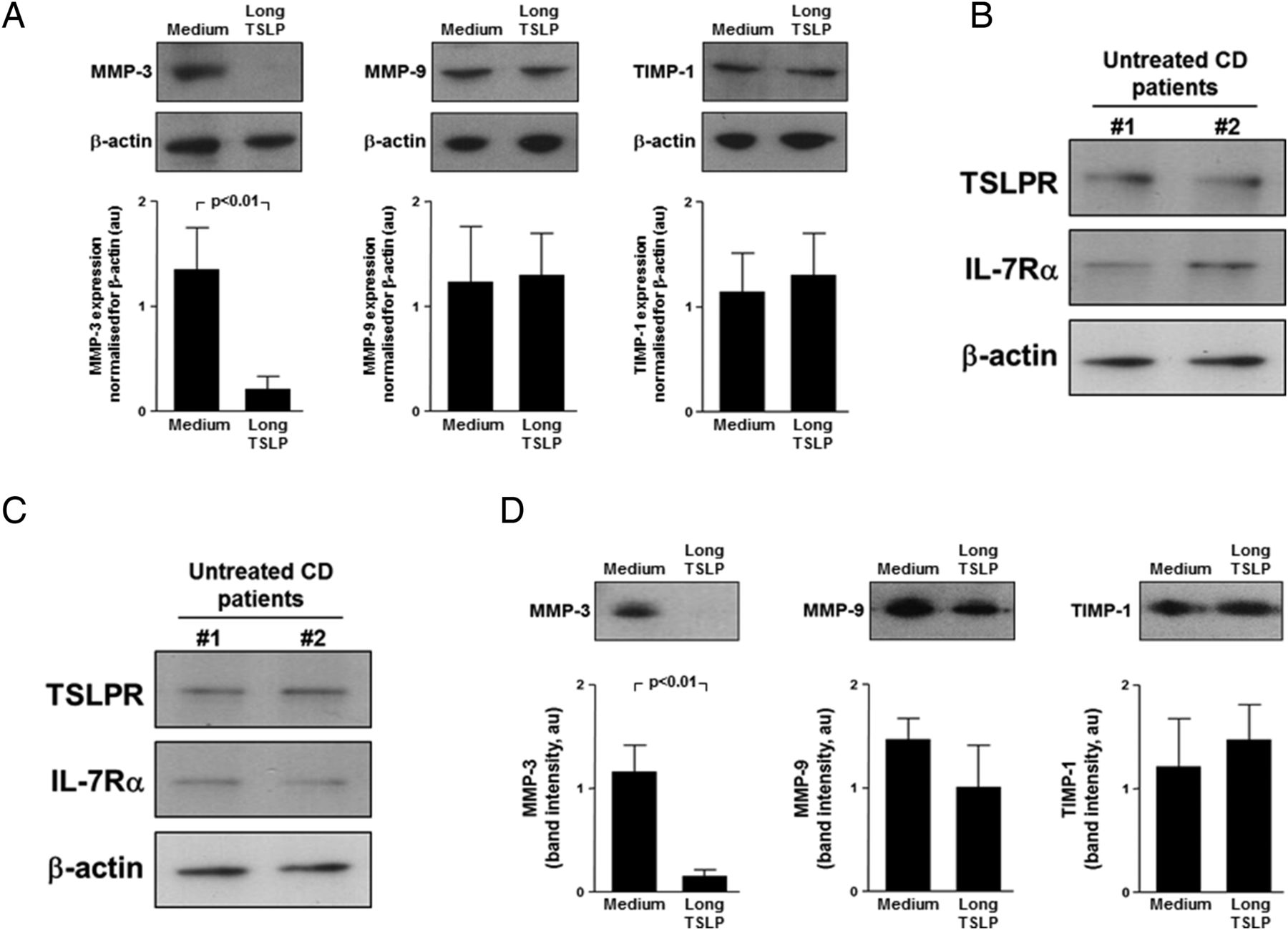
In untreated CD, there is increased MMP activation,15 ,25 thus we analysed the ex vivo effects of long TSLP on MMP and TIMP-1 production by untreated CD biopsies. Long TSLP significantly (p<0.01) downregulated MMP-3, whereas it did not change MMP-9 or TIMP-1 production (figure 6A). As intestinal myofibroblasts are an important MMP source,29 ,30 we assessed the in vitro effects of long TSLP on untreated CD myofibroblast production of MMPs and TIMP-1. Untreated CD myofibroblasts expressed TSLPR and IL-7Rα, both when freshly isolated (figure 6B) and at passage 4 (figure 6C). Supernatants of myofibroblasts cultured with long TSLP showed significantly (p<0.01) lower MMP-3 levels than unstimulated myofibroblasts, whereas long TSLP did not induce any significant change in MMP-9 and TIMP-1 production (figure 6D). Long TSLP significantly (p<0.005) downregulated IL-8 production by untreated CD myofibroblasts compared with medium only (from mean 218.5±37.2 pg/mL to 13.5±11.9 pg/mL) (see online supplementary figure S3B).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Ex vivo and in vitro effects of long thymic stromal lymphopoietin (TSLP) on matrix-metalloproteinase (MMP) production by untreated coeliac disease (CD) mucosa and myofibroblasts. (A) Recombinant human long TSLP was cultured with biopsies from six untreated patients with CD and after 24 h culture MMP-3, MMP-9 and tissue inhibitor of matrix metalloproteinase-1 (TIMP-1) production was evaluated by immunoblotting in biopsy supernatants. Examples shown in the upper panel are representative of all experiments. Blots were stripped and analysed for β-actin. Lower panel shows immunoblot densitometry. Results are mean±SEM. (B) TSLP receptor (TSLPR) and interleukin (IL)-7Rα protein, detected by immunoblotting, on lysates of mucosal myofibroblasts freshly isolated from two untreated patients with CD. Blots were stripped and analysed for β-actin. (C) TSLPR and IL-7Rα protein, detected by immunoblotting, on lysates of mucosal myofibroblasts isolated from two untreated patients with CD and cultured until passage 4. Blots were stripped and analysed for β-actin. (D) Effect of TSLP on MMP-3, MMP-9 and TIMP-1 release by untreated CD myofibroblasts. Examples shown in the upper panel are representative of six independent experiments on two different cell lines. Blots were stripped and analysed for β-actin. Lower panel shows immunoblot densitometry. Results are expressed as mean band intensity (arbitrary units)±SEM. au, arbitrary units.
Discussion
TSLP, an immunoregulatory cytokine constitutively expressed by enterocytes, plays an important role in preserving mucosal tolerance during homeostasis in the gut.31 Conversely, mucosal TSLP deficiency has been reported in Crohn's disease, and this contributes to chronic inflammation due to the consequent impairment in tolerogenic dendritic cell differentiation.5 ,20 In keeping with the findings by Fornasa et al,9 we observed that both long and short TSLP mucosal transcripts are reduced in active patients with CD compared with treated CD and controls. In the atrophic duodenal mucosa of both untreated and refractory patients with CD, we observed a marked TSLP reduction both at gene and protein levels. In parallel, we found a reduced number of epithelial cells per unit length of muscularis mucosae in refractory and untreated CD mucosa, suggesting that reduced TSLP may be a consequence of enterocyte loss. However, TSLP reduction in active CD was confirmed upon normalising TSLP transcripts for the epithelium-specific gene CK18, and this indicates that this deficiency is at least in part irrespective of the epithelial damage. There was no difference in mucosal TSLP expression both at protein and mRNA levels between type 1 and 2 refractory CD. TSLP expression normalised in treated CD, suggesting that TSLP deficiency might not be a primary defect in CD. This concept is further supported by unchanged TSLP content in architecturally normal potential CD mucosa.
As an additional mechanism to explain TSLP defect in CD, we hypothesised an increased TSLP degradation by furin, a calcium-dependent serine endoprotease capable of cleaving TSLP.26–28 Furin was overexpressed in active CD mucosa compared with both treated CD and controls. We then verified in an in vitro cleavage assay whether supernatants of duodenal biopsies could degrade long TSLP. Untreated and refractory CD supernatants showed a higher TSLP-degrading ability compared with treated CD and control supernatants, as shown by the higher intensity of the cleavage fragment after 2 h and the reduced intensity of the band corresponding to intact long TSLP. Therefore, cleavage may represent an additional mechanism accounting for TSLP deficiency in untreated CD.
TSLP exerts its functions by binding to the heterodimer formed by IL-7Rα and TSLPR. We observed comparable levels of both TSLPR and IL-7Rα in the duodenal mucosa of refractory, untreated, treated and potential CD and controls. Moreover, we could detect both TSLPR and IL-7Rα in the lysates from unfractionated duodenal LPMCs and myofibroblasts from the duodenal mucosa of untreated patients with CD. The evidence that TSLPR co-localises with CD11c and CD14 on the surface of LPMCs in untreated CD supports the notion that TSLPR is expressed by mucosal phagocytes, in keeping with the well-known increased lamina propria dendritic cell infiltration in untreated CD.18
TSLP reduction in active CD mucosa may have important immunological consequences. Since TSLP signalling through TSLPR results in downstream STAT5 phosphorylation,32 we evaluated pSTAT5 in untreated CD biopsies challenged with long or short TSLP. Only long TSLP upregulated pSTAT5, whereas short TSLP was unable to induce STAT5 phosphorylation, in keeping with previous observations.8 ,9 We observed that both long and short TSLP downregulate T-bet expression and IFN-γ production by untreated CD biopsies. Moreover, the number of IFN-γ-producing untreated CD LPMCs, which expressed TSLPR, was reduced by the culture with long TSLP, in keeping with the demonstration that TSLP may exert its action directly on mucosal T cells.23 ,33 These findings support the concept that TSLP deficiency in active CD may have a role in upregulating the Th1 mucosal immune response, which plays a crucial role in the development of villous atrophy.10 ,12 ,13 While our results confirm the anti-inflammatory properties of short TSLP, the reduction of IFN-γ production observed upon culture with long TSLP is in apparent contrast with the results of Fornasa et al,9 who reported that the long isoform is responsible for an increased IFN-γ production in human mixed lymphocyte reaction. In the same type of in vitro experiment, we observed that furin-exposed long TSLP induces a reduction of IFN-γ release by peripheral blood mononuclear cells to a similar extent as short TSLP. The effects of long TSLP on IFN-γ production by untreated CD biopsies and LPMCs may derive from TSLP proteolytic cleavage, in keeping with our results on TSLP degradation by active CD organ culture supernatants and furin. Additionally, both long and short TSLP reduced untreated CD biopsy production of IL-8, a chemokine promoting immune cell recruitment into inflamed gut, and which we showed to be produced in excess by untreated CD biopsies grown ex vivo.
Based on recent findings showing a marked TSLP upregulation in several chronic inflammatory disorders characterised by dysregulated tissue remodelling,34–36 a role for TSLP in modulating extracellular matrix turnover was proposed. Since abnormal extracellular matrix turnover is implicated in promoting villous atrophy in CD,15 ,25 ,37 we assessed the ex vivo influence of long TSLP on MMP-3, MMP-9 and TIMP-1 release.38 TSLP addition to active CD biopsies led to a significant MMP-3, but not MMP-9 and TIMP-1, reduction. As intestinal myofibroblasts are among the most important MMP-producing cell types,29 ,30 upon having verified their expression of TSLPR and IL-7Rα in CD, we evaluated long TSLP effect on production of MMPs by untreated CD myofibroblasts. TSLP markedly reduced MMP-3 production. These findings suggest that TSLP deficiency may contribute to tissue damage by upregulating MMP-3, confirming the pathogenic role of this protease in active CD.15 ,25 Finally, as intestinal myofibroblasts produce IL-8,39 we measured IL-8 production by untreated CD myofibroblasts following TSLP stimulation. Long TSLP reduced IL-8 production by untreated CD myofibroblasts, highlighting a new role for such cells, considered to have almost exclusively architectural functions so far. Remarkably, our results on the effects of long TSLP in active CD were obtained on biopsies and cell types, such as LPMCs and myofibroblasts, which are an important source of proteolytic enzymes; therefore, we cannot exclude that the anti-inflammatory properties of the long TSLP are the result of a protease-cleaved peptide.
In conclusion, defective TSLP content in active CD mucosa may contribute to the upregulation of T-cell-derived cytokines and myofibroblast-released matrix-degrading enzymes. The potent action demonstrated by TSLP in dampening tissue inflammation and damage in CD suggests that restoring TSLP function might be therapeutically useful at least in refractory patients with CD, who no longer respond to GFD.
Acknowledgments
The authors would like to thank all patients that agreed to give biopsies and tissue samples without whom this study would not have been possible.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
Footnotes
Correction notice This article has been corrected since it published Online First. Figures 1 and 3 have been replaced with colour images.
Contributors PB and ADS contributed equally to this manuscript and should be considered joint first authors. PB, PG, GF, KT, SLFP, AP, CU and AV performed the experimental work. PB, ADS, PG and GF performed data analysis. ADS, CP, AF and EW provided clinical samples. PB, ADS, MR, TTM and GRC designed the study. PB and ADS drafted the manuscript. All authors approved the final version of the manuscript.
Funding This project was partly supported by the Associazione Italiana Celiachia and by the European Union–funded project TORNADO (FP7-KBBE-222720).
Competing interests PG was supported by a grant from Ghislieri College (Pavia, Italy).
Patient consent Obtained.
Ethics approval Ethical Committee, St. Matteo Hospital, Pavia.
Provenance and peer review Not commissioned; externally peer reviewed.