Article Text

Abstract
Objectives Chronic-plus-binge ethanol feeding activates neutrophils and exacerbates liver injury in mice. This study investigates how recent excessive drinking affects peripheral neutrophils and liver injury in alcoholics, and how miR-223, one of the most abundant microRNAs (miRNAs) in neutrophils, modulates neutrophil function and liver injury in ethanol-fed mice.
Designs Three hundred alcoholics with (n=140) or without (n=160) recent excessive drinking and 45 healthy controls were enrolled. Mice were fed an ethanol diet for 10 days followed by a single binge of ethanol.
Results Compared with healthy controls or alcoholics without recent drinking, alcoholics with recent excessive drinking had higher levels of circulating neutrophils, which correlated with serum levels of alanine transaminase (ALT) and aspartate transaminase (AST). miRNA array analysis revealed that alcoholics had elevated serum miR-223 levels compared with healthy controls. In chronic-plus-binge ethanol feeding mouse model, the levels of miR-223 were increased in both serum and neutrophils. Genetic deletion of the miR-223 gene exacerbated ethanol-induced hepatic injury, neutrophil infiltration, reactive oxygen species (ROS) and upregulated hepatic expression of interleukin (IL)-6 and phagocytic oxidase (phox) p47phox. Mechanistic studies revealed that miR-223 directly inhibited IL-6 expression and subsequently inhibited p47phox expression in neutrophils. Deletion of the p47phox gene ameliorated ethanol-induced liver injury and ROS production by neutrophils. Finally, miR-223 expression was downregulated, while IL-6 and p47phox expression were upregulated in peripheral blood neutrophils from alcoholics compared with healthy controls.
Conclusions miR-223 is an important regulator to block neutrophil infiltration in alcoholic liver disease and could be a novel therapeutic target for the treatment of this malady.
- INFLAMMATION
- ETHANOL
- CYTOKINES
- FATTY LIVER
- LEUKOCYTES
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Chronic-plus-binge ethanol feeding synergistically induces hepatic neutrophil infiltration, elevation of peripheral blood neutrophils and liver injury in mice.
Neutrophils contribute to liver injury in mice fed with chronic-plus-binge ethanol.
What are the new findings?
Alcoholics with recent excessive drinking have significantly higher levels of circulating neutrophils, serum alanine transaminase (ALT) and aspartate transaminase (AST) compared with those without recent drinking and healthy controls.
Circulating neutrophils correlate positively with serum ALT and AST levels only in alcoholics with recent drinking but not in those without recent drinking or healthy controls.
miR-223, the most abundant microRNA in neutrophils, is downregulated in neutrophils of alcoholics with recent excessive drinking when compared with healthy controls.
Genetic deletion of miR-223 gene exacerbates hepatic neutrophil infiltration, neutrophil reactive oxygen species (ROS) production and liver injury in chronic-plus-binge ethanol-fed mice.
miR-223 inhibits the interleukin-6–p47phox–ROS pathway in neutrophils, thereby protecting against alcohol-induced liver injury.
How might it impact on clinical practice in the foreseeable future?
Physicians should pay attention to recent drinking history when reviewing complete blood count results from alcoholics. Significant elevation of peripheral neutrophils may indicate recent drinking in these subjects.
miR-223 plays a critical role in limiting neutrophil infiltration in alcoholic liver disease and could be a novel therapeutic target for the treatment of this malady.
Introduction
Alcoholic liver disease (ALD), a leading cause of chronic liver disease, comprises a spectrum of disorders ranging from simple fatty liver to steatohepatitis, cirrhosis and liver cancer.1–3 While fatty liver develops in almost all subjects with heavy alcohol use, steatohepatitis with hepatic neutrophil infiltration occurs in only 20–40% of these subjects. Once developed, steatohepatitis can progress to cirrhosis and liver cancer.1 ,3 ,4 It is generally accepted that hepatic neutrophil infiltration is an important factor in promoting the progression of ALD.5 ,6 However, the mechanism triggering neutrophil infiltration and the roles of neutrophils in the pathogenesis of ALD are still elusive. In the past, the mechanistic study on the roles of neutrophils in ALD was limited due to negligible hepatic neutrophil infiltration in the widely used chronic ad libitum ethanol-feeding models. The recently developed chronic-plus-binge ethanol feeding model mimics acute-on-chronic alcohol-induced liver injury with significant hepatic neutrophil infiltration.7–9 Weekly ethanol binge also causes a shift in hepatic infiltration from macrophages to neutrophils in chronically intragastric ethanol-fed mice.10 By using these feeding models, several recent studies suggest that acute ethanol binge promotes hepatic neutrophil infiltration leading to exacerbation of liver injury in chronically ethanol-fed mice.7 ,11 The contribution of neutrophil infiltration in alcohol-induced liver injury in animal models is an important observation with significant clinical implications. Neutrophils are the major circulating leucocytes in human, accounting for 60–70% or ∼3–6×109 cells/L of total white blood cells (WBCs). On the contrary, only 10–15% (∼1–2×109 cells/L) of the circulating WBCs in mice are neutrophils. It is plausible that neutrophils may play a more important role in the pathogenesis of ALD in humans than that in rodents, given the higher proportion of circulating neutrophils. In the present study, we analysed the haematogram of 300 alcoholics and found the elevation of circulating neutrophils in correlation with serum transaminases, notably in 140 alcoholics with recent excessive drinking. Our observation suggests that neutrophils may contribute to early liver injury in these subjects.
Hepatic neutrophil infiltration, a hallmark of alcoholic steatohepatitis, functions against bacterial infection and also exacerbates hepatocyte injury, liver inflammation and fibrosis via the generation of reactive oxygen species (ROS), release of proteases and production of proinflammatory mediators.5 ,12 ROS production in neutrophils is controlled by the multimeric transmembrane enzyme complex nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX), which generates superoxide (O2•−) and hydrogen peroxide (H2O2) from molecular oxygen using NADPH as an electron donor.13 ,14 The mammalian NOX family contains seven isoforms: NOX1–5, DUOX1 and DUOX2. In the liver, the main ROS-producing NOXs are NOX1, NOX2 and NOX4.13 ,14 In addition, several NOX-associated molecules are required for NOX activation by stabilising the NOX proteins and docking cytosolic factors, including phagocytic oxidase (phox) units: p47phox (also called neutrophil cytosolic factor-1: NCF-1), p67phox (NCF-2), p40phox (NCF-4), p22phox and gp91phox.13 ,14 Emerging evidence suggests that ROS production is probably an important mechanism by which neutrophils induce tissue damage including liver injury and fibrosis.14–17 However, how NOXs and their related proteins are regulated in neutrophils remains obscure.
MicroRNAs (miRNAs, small non-coding RNA molecules with 19–25 nucleotides) play an important role in a variety of cellular functions by inducing RNA silencing and involving in post-transcriptional regulation of gene expression. In recent years, numerous studies have demonstrated that miRNAs are involved in the pathogenesis of various types of liver diseases,18–21 including ALD.22–28 For example, a subset of miRNAs have been identified to regulate the lipopolysaccharide (LPS)-mediated inflammatory signalling cascades in hepatocytes and Kupffer cells in rodents chronically fed with ethanol.22–26 miR-182 was found to be expressed at high levels and correlates with disease severity in patients with alcoholic hepatitis.29 All of these previous studies largely focused on the miRNAs in the liver and hepatocytes.22–29 In the current study, we performed miRNA microarray analysis of serum miRNAs from alcoholics and found that serum levels of miR-223, one of the most abundant miRNAs in neutrophils,30–32 were significantly elevated in alcoholics compared with healthy controls. The role of miR-223 was examined in miR-223−/− mice by using a model of chronic-plus-binge ethanol feeding. Our data suggest that miR-223 plays an important role in regulating neutrophil function and liver injury via the inhibition of the interleukin (IL)-6–p47phox pathway in ethanol-fed mice and alcoholics.
Materials and methods
Human subject cohort
Forty-five healthy controls were recruited at Roudebush Veterans Administration Medical Center (Indianapolis, Indiana, USA). Three hundred excessive chronic drinkers were recruited from Fairbanks Drug and Alcohol Treatment Center (Indianapolis, Indiana, USA). Based on the information from the Time Line Follow-Back questionnaire, we dichotomised excessive drinkers into alcoholics with recent excessive drinking (excessive drinking within 10 days before enrollment) and alcoholics without recent alcohol drinking (no drinking within 10 days before enrolment) (table 1). The detailed information about the study cohort is described in online supplementary materials. The study was approved by the Indiana University Purdue University Institutional Review Board, the Research and Development Committee of the Roudebush VA, and Fairbanks Drug and Alcohol Treatment Center. All participants provided written informed consent.
Supplementary materials
Demographic, clinical characteristics and laboratory results of the study cohort
Mice
Ten-week to 12-week-old female mice were used. C57BL/6J, miR-223−/− mice, p47phox−/− (also called NCF-1−/−) mice and elastase−/− mice were purchased from Jackson Laboratory (Bar Harbor, Maine, USA). The study was approved by the NIAAA ACUC Committee.
Mouse model of chronic-plus-binge ethanol feeding
Mouse model of short-term chronic (10 days)-plus-binge ethanol feeding was described previously.33 In some experiments, mice were subjected to long-term chronic (8 weeks)-plus-multiple binges of ethanol, as described previously.8
Real-time PCR analyses and the primer sequences are described in online supplementary materials and table S1.
Other methods and statistical analyses are described in online supplementary materials.
Results
The level of circulating neutrophils correlates with serum ALT and AST in alcoholics with recent excessive drinking
Recent data from chronic-plus-binge ethanol feeding model suggest that binge drinking elevates the levels of circulating and hepatic neutrophils, leading to liver injury in chronically ethanol-fed mice.7 To test the impact of binge drinking on circulating neutrophils and liver injury in humans, we carefully analysed the haematogram and hepatic panels of 300 alcoholics. We found that the level and percentage of circulating neutrophils, serum levels of alanine transaminase (ALT), aspartate transaminase (AST) and bilirubin were higher in alcoholics than those in healthy controls (table 1), suggesting some degree of liver injury in these subjects. We further dichotomised these subjects into alcoholics without recent drinking (representing chronic alcohol drinking) and alcoholics with recent excessive drinking (representing chronic-plus-binge drinking) (table 1). Interestingly, the level and percentage of circulating neutrophils, serum levels of ALT, AST and bilirubin were much higher in the latter group. Moreover, the level of circulating neutrophils positively correlated with serum ALT and AST levels in alcoholics with recent drinking, but no such correlation was found in alcoholics without recent drinking and healthy controls (figure 1).

The level of peripheral blood neutrophils correlates positively with serum ALT and AST levels in alcoholics with recent excessive drinking. Forty-five healthy controls and 300 alcoholics were enrolled, and alcoholics were stratified into two groups: alcoholics with (n=140) and without (n=160) recent excessive drinking (see table 1). The correlation analyses between the levels of circulating neutrophils and serum ALT and AST levels were performed. Significant correlations (p values) are labelled in the figure.
It is generally believed that females are more susceptible to ALD than males;2 however, there were no comprehensive studies that compared the early stages of ALD between male and female alcoholics. The detailed demographic, clinical characteristics, haematograms and hepatic panel results were stratified by genders (223 males and 77 females) and are shown in online supplementary tables S2–S4. The levels of neutrophils were positively correlated with serum ALT or AST levels only in male alcoholics with recent drinking. Such correlation was not observed when we analysed the data in females likely due to a smaller sample size (see online supplementary figure S1). It is interesting that the levels of serum AST and ALT were lower in female alcoholics, despite the comparable neutrophil counts in both genders (see online supplementary table S4). Further studies to recruit more female alcoholics are needed to explore the gender differences on the susceptibility to early ALD.
Regulation of miR-223 levels in serum and neutrophils from alcoholics and ethanol-fed mice
By screening serum miRNA microarray profiling, we found that serum levels of miR-223, the most abundant miRNA in neutrophils,30–32 were significantly elevated in alcoholics compared with those in healthy controls. Elevation of serum miR-223 in alcoholics was further validated by measuring miR-223 in serum (figure 2A). Elevation of serum miR-223 was also observed in two models of ethanol feeding compared with their pair-fed groups (figure 2B and online supplementary figure S2A).
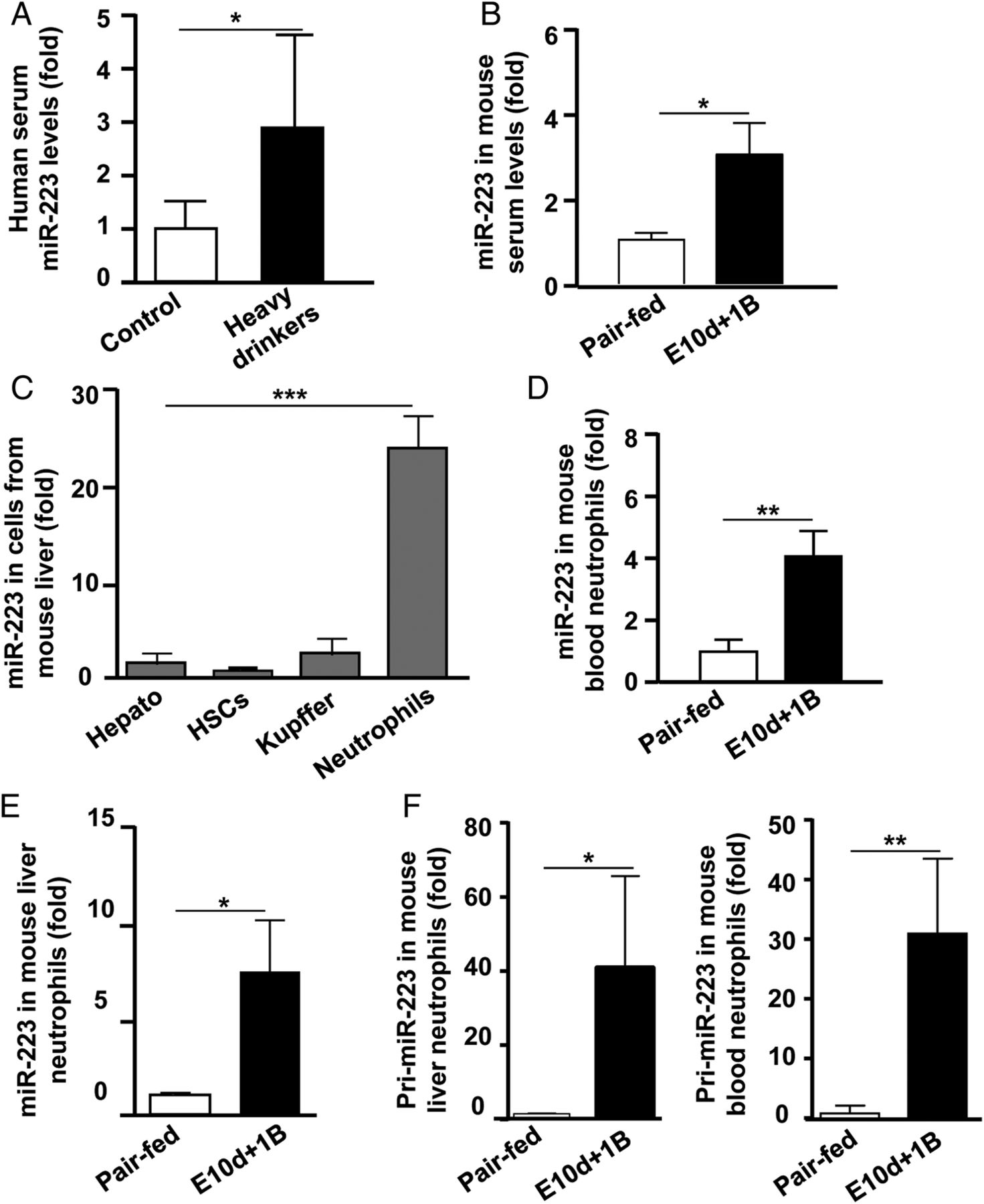
miR-223 levels in serum and neutrophils from alcoholics and ethanol-fed mice. (A) Blood samples were collected from healthy controls and excessive drinkers. miR-223 levels in serum were measured by real-time quantitative PCR. (B) C57BL/6J mice were pair-fed or fed an ethanol-containing diet for 10 days plus one binge (E10d+1B). Mice were euthanized 9 hours postgavage, and serum was collected for miR-223 measurement. (C) Various types of cells were isolated from naïve C57BL6/J livers and subjected to RT-qPCR for miR-223. (D–F) Mice were subjected to control and ethanol feeding as described in (B). miR-223 and pri-miR-223 levels in mouse peripheral and liver neutrophils were measured. Values represent means±SEM (n=6–14). *p<0.05; **p<0.01; ***p<0.001. HSC, hepatic stellate cells.
miR-223 is the most abundant miRNA in neutrophils,30–32 but its expression in hepatocytes, non-parenchymal cells and leukocytes in the liver is elusive. Here, we found that neutrophils had the highest levels of miR-223, which is about 15-fold to 24-fold higher than those in hepatocytes, hepatic stellate cells and Kupffer cells (figure 2C). Thus, we focused primarily on miR-223 in neutrophils in our current study. As illustrated in figure 2D, E, the expression of miR-223 in peripheral blood and hepatic neutrophils was upregulated after chronic-plus-binge ethanol feeding. In vitro incubation of neutrophils with ethanol or its metabolite, acetaldehyde, did not increase miR-223 levels, indicating that the upregulation of miR-223 was not due to the direct effect of ethanol or its metabolite (see online supplementary figure S2B–C). Furthermore, the expression of primary miR-223 (pri-miR-223) was significantly upregulated in hepatic and peripheral neutrophils from ethanol-fed mice than that from pair-fed controls (figure 2F), suggesting that transcriptional regulation is involved in the upregulation of miR-223 expression in neutrophils.
miR-223−/− mice are more susceptible to chronic-plus-binge-induced liver injury
To define the roles of miR-223 in ALD, wild-type (WT) and miR-223−/−, mice were subjected to chronic-plus-binge feeding. As illustrated in figure 3A, miR-223−/− mice had higher levels of serum ALT and AST after pair or ethanol feeding, compared with WT counterparts. In contrast, either chronic (10-day) or binge ethanol feeding alone did not elevate serum ALT and AST in both WT and miR-223−/− mice (see online supplementary figure S3A).

Deletion of the miR-223 gene exacerbates chronic-plus-binge ethanol-induced liver injury, elevation of circulating and hepatic neutrophils. Wild-type (WT) mice and miR-223−/− mice were subjected to E10d+1B or pair feeding, and mice were euthanised 9 hours postgavage. (A) Serum ALT, AST and hepatic triglyceride (TG) levels were measured. (B) The percentage and number of neutrophils were determined by a haematology analyser (two upper panels). The percentage of Gr1+CD11b+ circulating neutrophils were analysed by flow cytometer (lower panel). (C) Liver tissues were subjected to immunostaining with an anti-MPO antibody. The number of MPO+ cells was counted. (D) Liver tissues were subjected to real-time PCR analyses of Ly6G (a marker for neutrophils). (E) Liver leucocytes were isolated and subjected to flow cytometric analyses. The percentage of neutrophils (Gr1+CD11b+) cells was analysed. Values represent means±SEM (n=4–10). *p<0.05; **p<0.01; ***p<0.001.
Interestingly, although serum ALT and AST levels were higher in chronic-plus-binge ethanol-fed miR-223−/− mice, hepatic triglyceride levels were lower in these mice compared with ethanol-fed WT mice (figure 3A). The lesser degree of hepatic steatosis in ethanol-fed miR-223−/− mice may be due to higher levels of IL-6 (see results below), a cytokine known to ameliorate steatosis, than those in WT mice.34
The above data in figure 3 suggest miR-223 deficiency exacerbates alcohol-induced liver injury. We next tested whether overexpression of miR-223 ameliorates alcohol-induced liver injury via the injection of pre-miR-223 lentivirus through tail vein. As illustrated in online supplementary figure S3B, the ethanol-fed mice with pre-miR-223 lentivirus injection had lower serum ALT levels than those with control lentivirus injection. Injection of pre-miR-223 lentivirus resulted in an increase in expression of miR-223 in bone marrow neutrophils and in the liver. There was a trend in an increase in miR-223 expression in blood neutrophils from pre-miR-223 lentivirus injected mice but did not reach statistical difference (one possibility for this small increase was due to high basal levels of miR-223 in neutrophils).
As illustrated in figure 3B, both the ratio and the level of circulating neutrophils were increased after chronic-plus-binge feeding with higher levels in miR-223−/− mice than those in WT mice. This finding was also confirmed by flow cytometric analyses of CD11b+Gr-1+neutrophils (figure 3B). Furthermore, the data from online supplementary figure S4 suggest that higher levels of neutrophils in ethanol-fed miR-223−/− mice are likely due to the increase in neutrophil release from the bone marrow, rather than increased proliferation or decreased neutrophil apoptosis. Finally, the ratio and level of lymphocytes and monocytes decreased after chronic-plus-binge ethanol feeding with greater reduction in miR-223−/− mice than those in WT mice (see online supplementary figure S5A).
miR-223−/− mice are more susceptible to chronic-plus-binge feeding-mediated induction of hepatic neutrophil infiltration and reduction of Kupffer/macrophages
Hepatic neutrophils were examined by immunohistochemistry, real-time quantitative PCR and flow cytometric analyses. As illustrated in figure 3C and online supplementary figure S5B, the number of myelopeoxidase (MPO)+ neutrophils in the liver was elevated in both WT and miR-223−/− mice after chronic-plus-binge feeding, with higher number in miR-223−/− mice. Hepatic expressions of Ly6G (a marker for neutrophils) were also significantly increased after ethanol feeding, and such upregulation was twofold higher in miR-223−/− mice than that in WT mice (figure 3D). Finally, flow cytometric analyses revealed that the percentage of hepatic Gr1+CD11b+ neutrophils was higher in miR-223−/− mice compared with that in WT mice (figure 3E).
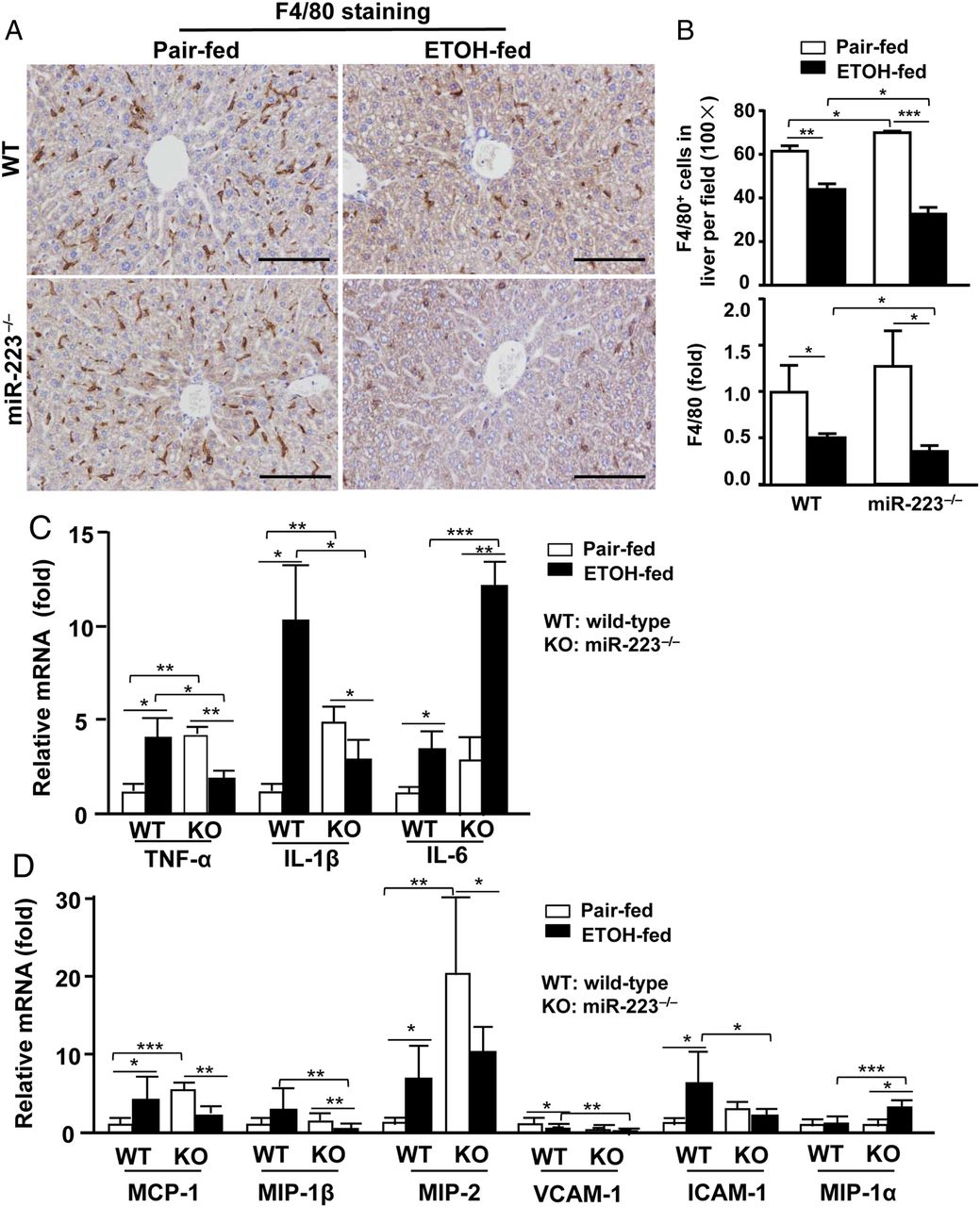
In contrast to the elevation of hepatic neutrophils, the number of F4/80+ Kupffer cells/macrophages and hepatic expression of F4/80 were decreased in WT and miR-223−/− mice after chronic-plus-binge feeding with a greater reduction in miR-223−/− mice (figure 4A, B). This greater reduction of macrophages in ethanol-fed miR-223−/− mice was probably due to increased apoptosis as demonstrated by the higher number of apoptotic TUNEL+F4/80+ cells in miR-223−/− mice than in WT mice (see online supplementary figure S6).

Macrophages and hepatic expression of proinflammatory mediators in ethanol-fed and pair-fed wild-type (WT) and miR-223−/− mice. WT mice and miR-223−/− mice were subjected to E10d+1B or pair feeding, and mice were euthanised 9 hours postgavage. (A and B) Liver tissues were subjected to immunostaining with an anti-F4/80 antibody. Representative photographs are shown in panel A (scale bar: 100 μm). The number of F4/80 was counted in panel B. Hepatic F4/80 mRNA was detected by real-time PCR and is shown in panel B. (C and D) Liver tissues were subjected to real-time PCR analyses. Values represent means±SEM (n=5–10). *p<0.05; **p<0.01; ***p<0.001.
Differential regulation of hepatic proinflammatory mediators in WT and miR-223−/− mice post chronic-plus-binge feeding
Hepatic expression of several proinflammatory mediators was examined and shown in figure 4C,D. These results are summarised as follows. First, chronic-plus-binge feeding resulted in the upregulation of several hepatic proinflammatory mediators in WT mice, which are consistent with previous findings.7 Second, hepatic expression of several mediators (eg, tumor necrotic factor-α (TNF-α), IL-1β, monocyte chemoattractant protein-1 (MCP-1) and macrophage inflammatory protein-2 (MIP-2)) was higher in pair-fed miR-223−/− mice than in pair-fed WT counterparts, but were reduced in miR-223−/− mice after chronic-plus-binge challenge. Third, hepatic expression of IL-6 was elevated in both miR-223−/− and WT mice after ethanol feeding with much higher levels in miR-223−/− mice than that in WT mice.
MiR-223−/− mice are more susceptible to chronic-plus-binge feeding-induced ROS production in the liver and neutrophils
Hepatic levels of lipid peroxide including malonaldehyde (MDA) and 4-hydroxynonenal (4-HNE) were examined by immunohistochemistry. The data in figure 5A, B show that the levels of hepatic MDA and 4-HNE expression in WT and miR-223−/− mice were elevated after chronic-plus-binge feeding, with a greater increase in miR-223−/− mice.

miR-223−/− mice are more susceptible to chronic-plus-binge ethanol-induced oxidative stress in the liver and circulating neutrophils. WT mice and miR-223−/− mice were subjected to E10d+1B or pair feeding, and mice were euthanised 9 hours postgavage. (A and B) Liver tissues were subjected to immunostaining with an anti-malonaldehyde (MDA) or anti-4-hydroxynonenal (HNE) antibody. Representative images are shown in panel A. Relative staining is quantified and shown in panel B (scale bar: 200 μm). (C and D) Hepatic and peripheral neutrophils were isolated and stimulated with or without phorbol 12-myristate 13-acetate (PMA). The reactive oxygen species (ROS) levels were measured by flow cytometric analyses. The ROS levels are calculated and shown. Values represent means±SEM (n=4–8). *p<0.05; **p<0.01; ***p<0.001.
Next, we used flow cytometer analysis to examine ROS release by both circulating and liver neutrophils from pair-fed and ethanol-fed mice. As illustrated in figure 5C, in pair-fed groups, the phorbol 12-myristate 13-acetate (PMA)-stimulated ROS levels were significantly higher in hepatic neutrophils from miR-223−/− mice than those from WT mice. Chronic-plus-binge feeding further increased PMA-stimulated ROS levels in neutrophils from WT mice but did not further increase in those from miR-223−/− mice.
Figure 5D shows ROS levels in circulating neutrophils. PMA-stimulated ROS levels were significantly higher in pair-fed miR-223−/− mice than that in pair-fed WT mice. Ethanol feeding further increased ROS levels in WT and miR-223−/− mice with higher levels in the miR-223−/− mice than those in WT mice.
ROS levels were also measured in hepatic macrophages, which were much lower than those in neutrophils. Ethanol feeding decreased PMA-stimulated ROS in macrophages from miR-223−/− mice (see online supplementary figure S7).
miR-223 indirectly downregulates p47phox in neutrophils via the inhibition of IL-6
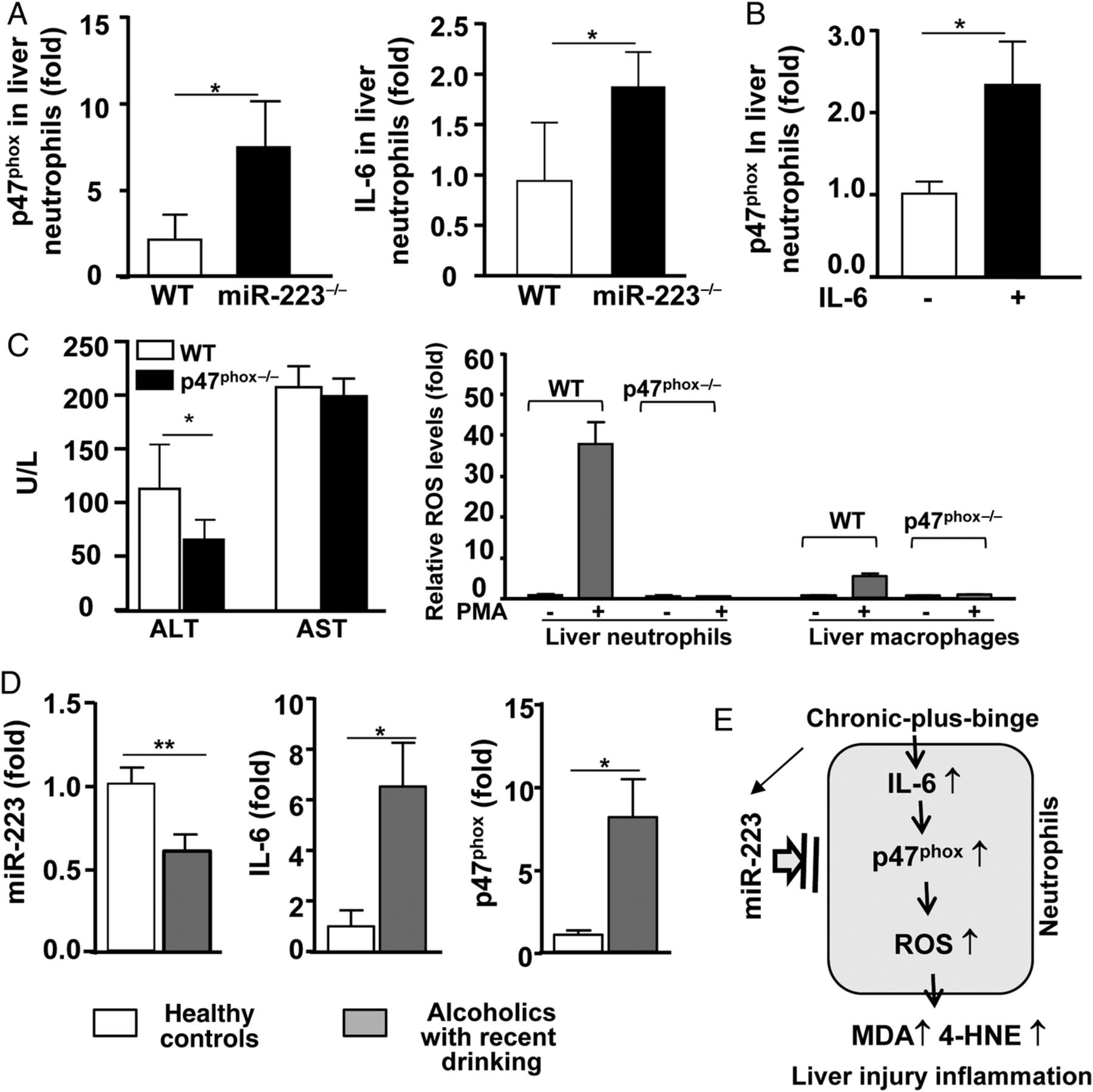
To understand the mechanisms underlying greater ROS in miR-223−/− mice versus WT mice, we examined hepatic expression of NADPH-associated molecules, which consist of five phagocytic oxidase (phox) units: p47phox, p67phox, p40phox, p22phox and gp91phox.14 As illustrated in figure 6A, chronic-plus-binge feeding increased hepatic p47phox expression in miR-223−/− mice compared with the pair-fed groups, while such feeding did not alter hepatic p47phox expression in WT mice. Interestingly, the basal levels of hepatic p47phox expression were higher in pair-fed miR-223−/− mice than those in pair-fed WT mice. In contrast, hepatic expression of other phox genes (p67, p40, p22 and gp91) was either not altered or elevated post ethanol feeding. Moreover, hepatic expression of Nox1, Nox3 and Nox4 was upregulated after chronic-plus-binge feeding with much higher levels in miR-223−/− mice compared with WT mice (figure 6B).

MiR-223 regulates reactive oxygen species (ROS)-associated genes in the liver from ethanol-fed mice. WT mice and miR-223−/− mice were subjected to E10d+1B or pair feeding, and mice were euthanised 9 hours postgavage. Liver tissues were subjected to real-time PCR analyses of ROS-associated genes. Values represent means±SEM (n=4–11). *p<0.05; **p<0.01;***p<0.001.
Because p47phox is predominately expressed in neutrophils and plays a key role in generating oxidative burst by neutrophils,35 ,36 we further examined the p47phox in neutrophils in this study. As illustrated in figure 7A, p47phox mRNA expression in neutrophils from ethanol-fed miR-223−/− mice was higher compared with WT mice. To understand the mechanism by which gp47phox is upregulated in miR-223−/− neutrophils, we examined miR-223 target sequence; however, no target sequences were found in p47phox gene, suggesting that miR-223 may indirectly regulate gp47phox. Because IL-6 was reported to be a target gene of miR-22337 ,38 and hepatic IL-6 gene expression was highly elevated in ethanol-fed miR-223−/− mice (figure 4C), we hypothesised that miR-223 indirectly inhibits p47phox expression in neutrophil via the downregulation of IL-6. First, we confirmed that IL-6 is a miR-223 target gene in online supplementary figure S8, showing that the 3′ untranslated region (UTR) region of IL-6 gene contains a miR-223-binding region, and transfection of miR-223 inhibited the activity of IL-6–3′ UTR gene reporter. Second, IL-6 expression was higher in neutrophils from ethanol-fed miR-223−/− mice compared with that in WT mice (figure 7A). Third, in vitro treatment with IL-6 markedly upregulated p47phox expression in neutrophils (figure 7B).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
MiR-223 regulates circulating neutrophils by targeting the IL-6–p47phox axis in ethanol-fed mice and alcoholics with recent drinking. (A) WT mice and miR-223−/− mice were subjected to E10d+1B or pair feeding, and mice were euthanised 9 hours postgavage. Circulating neutrophils were isolated and subjected to real-time PCR analyses. (B) Circulating neutrophils were isolated, and IL-6 (50 ng/mL) was added to the culture medium for 6 hours, and expression of p47phox was determined by real-time PCR analyses. (C) WT and p47phox−/− mice were subjected to E10d+1B feeding, and mice were euthanised 9 hours postgavage. Serum ALT and AST were measured (left panel). Liver neutrophils were isolated and treated with or without phorbol 12-myristate 13-acetate (PMA), and reactive oxygen species (ROS) was measured and calculated (right panel). (D) Circulating neutrophils were isolated from healthy controls and alcoholics with recent drinking, followed by real-time PCR analyses. (E) A summarised figure depicting the miR-223 as a modulator to control hepatic neutrophil infiltration and liver injury after chronic-plus-binge ethanol drinking. Values represent means±SEM (n=4–16). *p<0.05; **p<0.01.
The function of p47phox in the chronic-plus-binge feeding model was examined by using p47phox−/− mice. As illustrated in figure 7C, deletion of the p47phox gene diminished chronic-plus-binge-induced liver injury (elevation of ALT), whereas deletion of the elastase, a proteolytic enzyme in neutrophils, did not affect the liver injury in this model (see online supplementary figure S9A). Interestingly, only ALT levels were reduced in p47phox−/− mice, whereas serum AST levels were not reduced in these mice compared with WT mice (figure 7C). It appeared that in chronic-plus-binge feeding model, acute ethanol gavage directly caused hepatocyte mitochondrial injury and mitochondrial AST elevation, while elevated neutrophils contributed predominantly to ALT elevation. Thus, deletion of the p47phox gene did not affect serum AST levels, but inhibited neutrophils, thereby resulting in lower levels of ALT. In addition, compared with ethanol-fed WT mice, the number of peripheral neutrophils remained unchanged and hepatic neutrophils were higher in ethanol-fed p47phox−/− mice (see online supplementary figure S9B). PMA-stimulated ROS production was completely diminished in neutrophils from ethanol-fed p47phox−/− mice (figure 7C). PMA-stimulated ROS production in macrophages, which was eightfold lower compared with that in neutrophils, was also abolished in ethanol-fed p47phox−/− mice. To further define the role of p47phox in immune cells including neutrophils contributes to liver injury, we generated chimeric mice via the bone marrow transplantation. As illustrated in online supplementary figure S9C, the WT mice with p47phox-deficient bone marrow had much lower serum ALT compared with the WT mice with WT mouse bone marrow post chronic-plus-binge ethanol feeding.
The above data suggest that miR-223 controls IL-6 and p47phox gene expression in neutrophils in mice. To examine the potential effect of miR-223 on IL-6 and p47phox in circulating neutrophils in human alcoholics, blood neutrophils were isolated from another cohort of 16 alcoholics with recent excessive drinking and 12 healthy controls. As illustrated in figure 7D, the expression of miR-223 was lower, while IL-6 and p47phox expression was higher in circulating neutrophils from alcoholics compared with those from healthy controls.
Discussion
Recent studies suggest that binge ethanol feeding increases the levels of circulating and hepatic neutrophils, contributing to liver injury in chronically ethanol-fed mice.7 In the present study, by analysing haematological and hepatic profiles in 300 alcoholics, we demonstrated that serum ALT and AST levels were much higher in alcoholics with recent drinking compared with those without recent drinking or healthy controls, which is in agreement with the results in mice showing the effect of chronic-plus-binge feeding on the elevation of serum ALT and AST.7 One of the key features from the mouse model of chronic-plus-binge feeding is the elevation of peripheral and hepatic neutrophils.7 ,39 ,40 Interestingly, in alcoholics with recent drinking, circulating peripheral neutrophils were substantially increased, accounting for approximately 80% of total WBC counts, which are significantly higher than those without recent drinking and healthy controls (approximately 60–67% of total WBC). Though we were unable to obtain liver biopsies to examine hepatic neutrophil infiltration due to ethical concerns, we found that the level of circulating neutrophils positively correlated very well with serum ALT and AST levels in alcoholics with recent drinking. These findings suggest that excessive drinking likely causes the increase in hepatic neutrophil infiltration, which leads to liver injury in these subjects.
By screening serum miRNAs, we found that the levels of serum miR-223, which is known to regulate neutrophil functions,30–32 were significantly higher in alcoholics compared with those in healthy controls. In agreement with these human data, serum miR-223 levels were also elevated after chronic-plus-binge ethanol feeding in mice. The role of miR-223 in ALD was tested in miR-223−/− mice. Interestingly, the pair-fed miR-223−/− mice had baseline phenotypes, as indicated by slightly but significantly higher serum ALT levels, peripheral neutrophil counts and expression of hepatic inflammatory mediators, when compared with pair-fed WT mice. This may be because that miR-223−/− mice had elevated circulating peripheral blood neutrophils and marked granulocyte hyperplasia in the bone marrow, which is mediated via the upregulation of the miR-223 targeted gene Mef2c, a transcription factor promoting myeloid progenitor proliferation.41 It is likely that the observed baseline phenotypes in these mice are secondary to the increase in circulating neutrophil levels. It is important to note that neither chronic ethanol feeding nor acute ethanol gavage alone augmented the baseline levels of serum AST and ALT in miR-223−/− mice. The escalation of liver injury as well as hepatic neutrophil infiltration only observed when chronic-plus-binge ethanol feeding was employed. These results suggest that the therapeutic implications of miR-223 may be applicable only to the conditions, which lead to the increase in hepatic neutrophils, such as chronic-plus-binge model, but not in chronic ethanol feeding or acute binge alone when hepatic neutrophil infiltration is not apparent.7
Mechanistic studies suggest that miR-223 attenuates neutrophil function by inhibiting the IL-6–p47phox–ROS pathway, thereby protecting against ALD (see summarised figure 7E). Among several NADPH oxidases and their associated genes related to ROS production, the expression of p47phox in the liver had the highest fold upregulation in ethanol-fed miR-223−/− mice compared with that in WT mice. A previous study reported that the deletion of the p47phox gene abolished chronic ethanol-feeding-induced liver injury through the reduction of ROS production by Kupffer cells.42 The results from our current study suggest a novel role of p47phox in neutrophils in promoting alcohol-induced liver injury. First, chronic-plus-binge model is associated with significant hepatic neutrophil infiltration but reduction in Kupffer cells. Second, neutrophils generated fivefold to eightfold higher levels of ROS than macrophages. Third, p47phox expression was highly elevated in hepatic neutrophils from ethanol-fed miR-223−/− mice compared with that in WT mice. Deletion of the p47phox gene completely abolished ROS production in neutrophils. Thus, we believe that p47phox in neutrophils plays a dominant role in promoting ROS production and liver injury in the chronic-plus-binge ethanol-feeding model.
In contrast to the increase in hepatic neutrophils in ethanol-fed miR-223−/− mice, hepatic F4/80+ macrophages were markedly decreased, when compared with those in pair-fed WT mice. The decrease in the number of macrophages was also consistent with the downregulation of several hepatic macrophage-associated cytokines and chemokines (such as TNF-α, MCP-1, MIP-1β and MIP-2) in ethanol-fed miR-223−/− mice versus the pair-fed group. A recent study reported that transfection of miR-223 mimics attenuated macrophage apoptosis by downregulating the expression of forkhead box O3 (FOXO3),43 an important inducer for macrophage apoptosis.44 Therefore, it is possible that the increased sensitivity of hepatic macrophages in miR-223−/− mice to ethanol-induced apoptosis was due to elevated expression of FOXO3. Finally, although miR-223 has been shown to play an important role in regulating macrophages in several types of diseases,45 ,46 its activation of macrophages may play a less important role in chronic-plus-binge ethanol-induced liver injury because hepatic macrophages were markedly decreased in this model.
Another interesting finding in the present study was that miR-223 expression was differentially regulated in neutrophils from human alcoholics and ethanol-fed mice although serum miR-223 levels were elevated in both groups. We demonstrated that in alcoholics, miR-223 in neutrophils was downregulated, whereas in chronic-plus-binge ethanol-fed mice, miR-223 in neutrophils was upregulated. At present, the mechanisms underlying this opposite regulation in humans and mice remain unknown. In addition, we postulate that upregulation of neutrophilic miR-223 in mice may play a compensatory role in protecting against alcohol-induced liver injury; whereas downregulation of neutrophilic miR-223 in alcoholics may exacerbate alcohol-mediated activation of neutrophils and liver injury in these subjects. Thus, modulation of miR-223 may be a novel strategy for the treatment of ALD through its regulation of neutrophil functions. Indeed, injection of pre-miR-223 lentivirus reduced chronic-plus-binge ethanol-induced liver injury. Because neutrophilic miR-223 ameliorated liver injury via the inhibition of neutrophils as demonstrated in this study, hepatic miR-223 promoted hepatocyte apoptosis;47 therefore, neutrophil-specific delivery of miR-223 may be needed in order to generate better therapeutic outcomes for the treatment of ALD.
References
Footnotes
Contributors ML planned and conducted the animal and cell culture study, and wrote the paper; YH, ZZ, TR, YqG, YhG, HC, YC, MX and DF performed the experiments and analysed the data; PZ designed and analysed bone marrow data; RAR conducted clinical study; SL planned and conducted clinical study, analysed the clinical data and edited the paper; BG designed the whole project, supervised the study and wrote the paper.
Funding This work was supported by the intramural programme of NIAAA, NIH (BG), by the 1I01CX000361-01 from the Veterans Affairs Research and Administration and W81XWH-12-1-0497 from USA Department of Defense (SL). ML was supported by Training Plan of Outstanding Young Medical Talents, Shanghai Municipal Health Bureau (XYQ2013093); YqG was supported by 3-year action plan of development of Traditional Chinese Medicine in Shanghai (ZYSNXD-CC-ZDYJ015).
Competing interests None declared.
Patient consent Obtained.
Ethics approval Indiana University Purdue University Institutional Review Board, the Research and Development Committee of the Roudebush VAMC, and Fairbanks Drug and Alcohol Treatment Center.
Provenance and peer review Not commissioned; externally peer reviewed.