Article Text

Abstract
OBJECTIVES The significance of the mast cell in the pathogenesis of rheumatic diseases has become more evident. Although mast cell hyperplasia is a feature of rheumatoid arthritis, the nature of mast cell chemoattractants involved in the recruitment of mast cells in joint diseases has not been studied in any detail. In this study the presence of mast cell chemotactic activity in synovial fluids was examined.
METHODS Synovial fluids from seven rheumatoid patients were tested in a modified Boyden chamber, where a human mast cell line was used as responder. The presence of stem cell factor (SCF) and transforming growth factor β (TGFβ) was measured by enzyme linked immunosorbent assay (ELISA).
RESULTS Six of the seven synovial fluids tested exhibited mast cell chemotactic activity. Two characterised human mast cell chemotaxins, SCF and TGFβ, were highly expressed in the synovium. Soluble SCF could be detected in all fluids analysed. Blocking antibodies against SCF or TGFβ almost completely blocked the activity in one fluid, partially blocked the activity in three, and did not affect the activity in two. Treatment of the responder cells with pertussis toxin reduced the migratory response against seven fluids, indicating the presence of chemoattractants mediating their effect through Gi coupled receptors.
CONCLUSION These data demonstrate the presence of multiple factors in synovial fluid acting as mast cell chemoattractants, two of which are SCF and TGFβ that contribute to the effect. These findings may be of importance for developing new strategies to inhibit mast cell accumulation in rheumatic diseases.
- mast cells
- migration
- stem cell factor
Statistics from Altmetric.com
Mast cells are inflammatory cells with importance for both acute and chronic inflammatory processes.1 These cells store a number of mediator molecules in their granules, such as histamine, heparin, and proteases, and can release these mediators into the extracellular space upon activation.2 Activated mast cells also synthesise prostaglandins and leucotrienes, and release both preformed and newly synthesised cytokines such as tumour necrosis factor α.2 ,3 Thus mast cells can release an array of mediators with the potential to cause inflammation and tissue remodelling. Therefore the mast cell is now considered to have a pivotal role in arthritis.4-6
Several histological studies have reported that mast cells are present in normal human synovia.7-9 Furthermore, mast cell hyperplasia has been described in rheumatoid synovial membranes.10-12 The mast cells are distributed throughout the synovial tissue as well as being present in the synovial fluid.13 The degree of mast cell hyperplasia in synovial tissues seems to be related to the degree of clinical synovitis and lymphocytic infiltration.14 The localisation of mast cells at the cartilage-pannus junction and in areas of matrix metalloproteinase deposition suggests a role for these cells in matrix remodelling.15 ,16 In addition, levels of mast cell mediators, including histamine, heparin, chymase, and tryptase, are increased in synovial fluids, indicating continuing mast cell activation.4 ,17 ,18 Administration of corticosteroids results in clinical improvement and can be accompanied by a decrease in mast cell numbers.14 These findings suggest that mast cells and their mediators may contribute to the initiation and progression of the destructive inflammatory process of rheumatoid arthritis.
The cause of mast cell hyperplasia in rheumatoid arthritis is not known. One possibility is an increased recruitment of mast cells and mast cell progenitors from other tissues into the inflamed joint. The recent reports that stem cell factor (SCF) and transforming growth factor β1 (TGFβ1) are mast cell chemoattractants have provided support for this hypothesis.19-21 The present study aimed at determining whether synovial fluid contains mast cell chemotactic activity.
Materials and methods
PATIENTS
Patients with active rheumatoid arthritis, as defined by the American College of Rheumatology Criteria,22 were recruited from the open care unit at the Department of Rheumatology, Karolinska Hospital, Stockholm. All were newly diagnosed (less than one year from appearance) and with active arthritis (joint pain and swollen joints). Table 1 summarises details of the patients. Biopsy specimens from patients with hallux valgus served as controls.23
Description of patients
SYNOVIAL FLUIDS AND BIOPSY SPECIMENS
Synovial fluids and biopsy specimens were taken from the knee joint. The fluids were obtained in heparinised vials, centrifuged at 1400×g for 10 minutes, and supernatants frozen at −70°C until investigation. Biopsy specimens were obtained by arthroscopy guided techniques and immediately snap frozen in isopentane on dry ice. All tissues were stored at −70°C until sectioned.
PREPARATION OF TISSUE SECTIONS AND IMMUNOSTAINING
For each monoclonal antibody two cryostat sections, 6–8 μm thick, were mounted on gelatin coated glass slides (Cel-Line, Newfield, USA). The slides were fixed in 50% cold acetone for 30 seconds, followed by 100% cold acetone. Endogenous peroxidase activity was blocked by 0.3% hydrogen peroxide in phosphate buffered saline (PBS) for one hour in the dark. After three additional washes in PBS the slides were incubated overnight at room temperature in a humid chamber with 100 μl of specific monoclonal antibody (antitryptase, monoclonal antibody 1222 from Chemicon International, London, UK, and anti-SCF, Genzyme Diagnostics, Cambridge, MA) dissolved in PBS containing 1% bovine serum albumin and 0.02% NaN3. The presence of TGFβ isoforms was determined as previously described24using affinity purified rabbit polyclonal antibodies: TGFβ1, No 96, TGFβ2, No 94, and TGFβ3, No 95 (kindly provided by Dr Keiko Funa, Gothenburg University, Sweden). Control staining was performed in parallel with isotype control (Dakopatts, Copenhagen, Denmark). After incubation with primary antibodies, slides were washed three times with PBS and incubated with 1% normal horse serum in PBS for 15 minutes at room temperature. Secondary antibodies (biotin-horse-antimouse IgG1, Vector Laboratory, Burlingame, CA), were added for 30 minutes at room temperature. After three washes with PBS, avidin-biotin-peroxidase (ABC) complexes (Vectastain ABC-HP kit standard, Vector Laboratory) were added for 45 minutes at room temperature. The colour reaction was developed for 15 minutes with 2.5 mg/ml AEC (3-amino-9-ethylcarbazole in 0.02 M sodium acetate pH 5.5 with 0.003% H2O2), generating a red colour. Counterstaining was performed with Mayer's haematoxylin for 1–5 minutes and the sections were mounted in glycerin-gelatin (Kaisers glycerin-gelatin, Merck, Darmstadt, Germany).
To test the specificity of the staining of the cytokine producing cells, blocking experiments were also performed for all antibodies used in the experiments. Recombinant cytokines were incubated with primary antibody at 10 times excess of the antibody concentration (20 μg/ml) at 4°C overnight. Subsequent staining was performed as described.
COMPUTER AIDED IMAGE ANALYSIS
Stainings were examined with a Polyvar II microscope (Reichert-Jung, Vienna, Austria). Tissue sections were evaluated by computer aided image analysis as previously described.25Briefly, the microscope was equipped with a three-chip charged couple device colour camera (DXC-750P, Sony Corporation, Tokyo, Japan) that digitised the microscope images to be processed in a Quantimet 600S image analyser (Leica Cambridge, Cambridge, UK) linked to a PC computer. The images were simultaneously transferred by the videocamera to a monitor, where the analytical decisions made by the computer were displayed in discriminating pseudocolours. This enabled the operator to accept or discard the result of analysis of a given field. A special software program (Posarea) was written in the high level language QUIPS and used to detect positively stained areas of the synovial tissue.25 Each microscope field was quantified using selected colour detection threshold values of single pixels in the digital image. The computer was then able to quantify the detected pixels to measure the area occupied by a specific staining of cytokines or of cells and express the results as a percentage of the total tissue area that was studied. A reading of an entire tissue section typically involved 50–200 microscope fields using a magnification power of ×250.
MEASUREMENTS OF CYTOKINE LEVELS IN SYNOVIAL FLUID
Measurements of TGFβ and SCF concentrations in synovial fluids were made by an enzyme linked immunoassay (ELISA) kit (sensitivity TGFβ 5 pg/ml and SCF 4 pg/ml) (R&D Systems, Abingdon, UK).
MAST CELL CULTURES
The human mast cell line, HMC-1, was established from a patient with mast cell leukaemia,26 and exhibited a phenotype of immature mast cells.27 HMC-1 cells were cultured in Iscove's modified Dulbecco's medium supplemented with 10% fetal calf serum (FCS), 2 mM l-glutamine, 100 IU/ml penicillin, 50 μg/ml streptomycin, and 1.2 mM α-thioglycerol. Cells were passaged every three to four days.
CHEMOTAXIS ASSAY
The migration of HMC-1 cells was assayed in a modified Boyden chamber by the leading front technique as previously described.20 Synovial fluids to be tested were diluted in medium. Thirty microlitres of the attractant was added to each well below the filter, and 50 μl of the cell suspension (1.5 x 106 cells/ml) was added above the filter. After 150 minutes at 37°C and 5% CO2, the filters were fixed, stained with Mayer's haemalum solution and mounted. The migration of mast cells against medium with 10% FCS served as control and was designated as 100% migration. Cell chemotaxis was assayed as the migration of the two furthest migrating cells visible in focus of one high power field (10 × 20). The migration distance on each filter was calculated as the mean of the readings of three different areas of the filter. The assay was always performed in triplicate at least.
INHIBITION OF ACTIVITY OF SYNOVIAL FLUID USING ANTI-CYTOKINE ANTIBODIES
Anti-TGFβ (pan-specific TGFβ neutralising antibody, R&D Systems Europe Ltd, Abingdon, UK) and anti-SCF (goat antihuman rSCF, R&D Systems) were added to the substrate suspension just before their addition to the chemotaxis assay. After a dose-response study of the two antibodies to determine optimal inhibitory concentrations for each agent the following concentrations were used: pan-specific TGFβ neutralising antibody (10 μg/ml), goat antihuman rSCF (10 μg/ml). Species-specific IgG (goat IgG for SCF and rabbit IgG for TGFβ, R&D Systems Europe Ltd) were used as controls at 10 μg/ml.
TREATMENT WITH INHIBITORS
HMC-1 cells were treated with 0.5 μg/ml pertussis toxin (Sigma Chemicals Co, St Louis, MO) or 0.5 μg/ml genistein (Calbiochem-Novabiochem Co, San Diego, CA). Inhibitor treatment was performed by incubating 2.5×106 cells/ml for 90 minutes at 37°C and 5% CO2, with each inhibitor in complete medium. The cells were then washed and resuspended at 1.5×106 cells/ml in complete medium before chemotaxis assay.
STATISTICAL ANALYSIS
The data in the text and figures are expressed as means (SEM), unless otherwise stated. Analysis of variance was used for analysis of significant differences. Differences were considered significant at p<0.05. The t test for correlation was used for calculation of significance.
Results
STAINING OF MAST CELLS AND STEM CELL FACTOR IN SYNOVIAL MEMBRANE
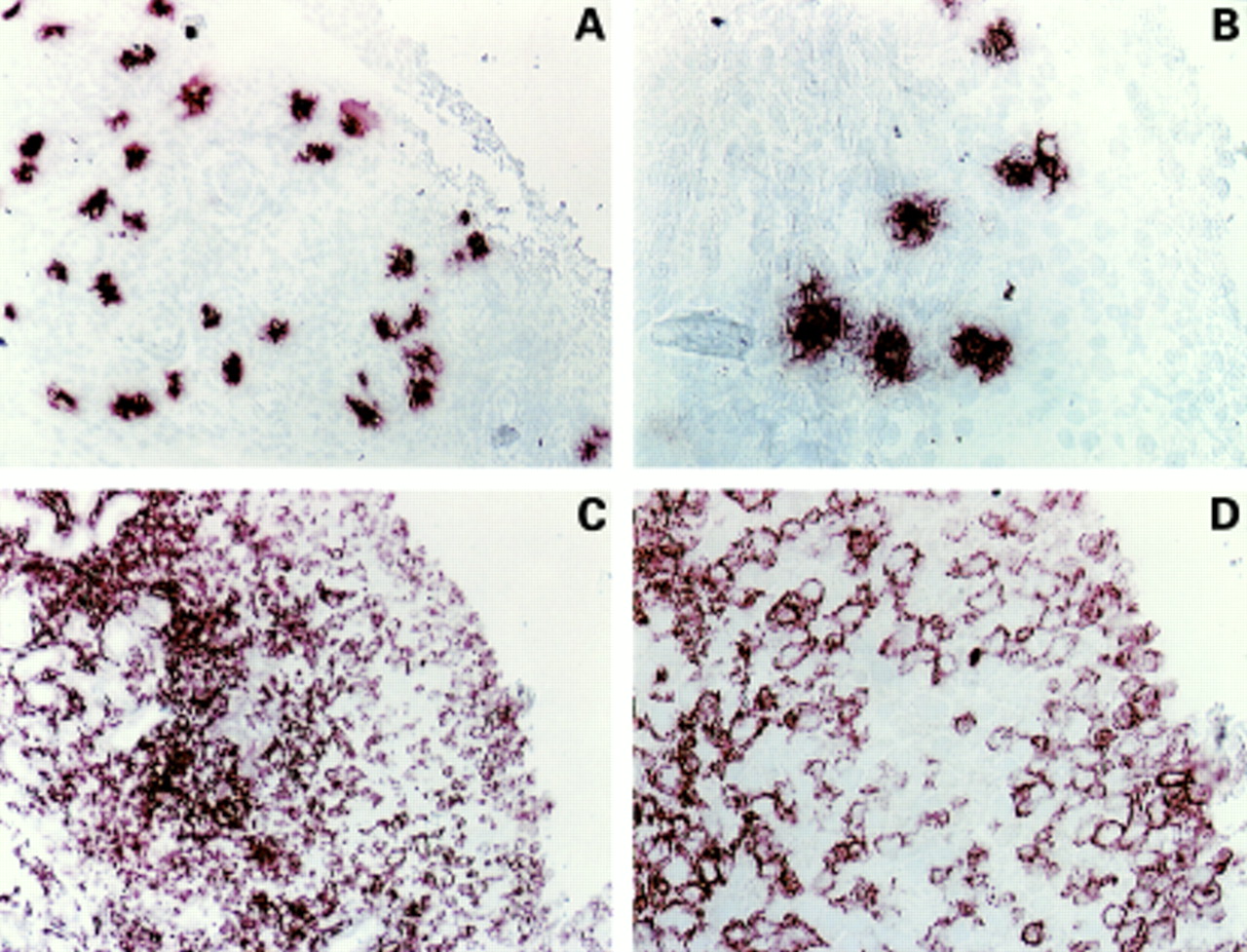
The number of mast cells is known to increase in the inflamed synovial tissue in arthritis. The most prominent factor for mast cells, important for several aspects of mast cell functions including migration, is stem cell factor. We therefore stained synovial tissue from patients with rheumatic diseases for the presence of mast cell tryptase and stem cell factor. All specimens of rheumatoid synovium indicated the presence of mast cells (table 2), but their distribution and local concentrations were variable. Mast cells were particularly located in the sublining area and in areas around vessels (figs 1A and B). Similarly, all analysed specimens had high expression of SCF (table2), in both the sublining and the lining areas, and around vessels (figs 1C and D). The cells immunopositive for SCF were identified as synoviocytes. Application of isotype control did not give any staining (data not shown).
Quantification of staining for mast cell tryptase and stem cell factor (SCF) in the synovium

Immunolocalisation of mast cells and stem cell factor (SCF) in synovial tissue. Tissues were stained for tryptase (A and B) or SCF (C and D). The figures are from the same patient (RD1), but different sections. The stainings are presented at two different magnifications from the same section. Magnification for A and C is ×80, and for B and D ×200.
We also investigated the presence of mast cells and the expression of SCF in the joints from five patients with hallux valgus, who lack signs of chronic inflammation.23 None of these specimens showed an increase in mast cell numbers or pronounced expression of SCF (negative or slightly positive, data not shown).
Transforming growth factor β has been implicated in the pathogenesis of rheumatic diseases28 and is also a prominent mast cell chemotaxin.21 The specimens were therefore analysed for the expression of TGFβ isoforms (TGFβ1, TGFβ2, and TGFβ3) and found to be positive (data not shown).
MAST CELL CHEMOTACTIC ACTIVITY IN SYNOVIAL FLUID
To address the issue of migration of mast cells into the inflamed synovial fluid in rheumatic disease, the presence of mast cell chemotactic activity in synovial fluid was analysed using the human mast cell line, HMC-1, in a bioassay. The synovial fluids were tested undiluted or diluted in medium (5–500 times) in a modified Boyden chamber. All except one (RD5) of the synovial fluids showed mast cell chemoattractant activity (fig 2). Five of them had strongest activity at ×5 dilution and one at ×125 dilution. Most known chemotaxins induce migration in a bell shaped dose-response curve, where higher concentrations inhibit migration. Thus the results obtained in this study indicate that the synovial fluids contain high concentrations of mast cell chemotactic activity. The optimal dilution of each fluid was used in all subsequent experiments. As a comparison of the migratory response to synovial fluids the efficacy of some of the identified mast cell chemotaxins is shown in table 3. Thus it is evident that the chemotactic activity in the synovial fluids gives a chemotactic response as strong or stronger than most of the identified mast cell chemotaxins.
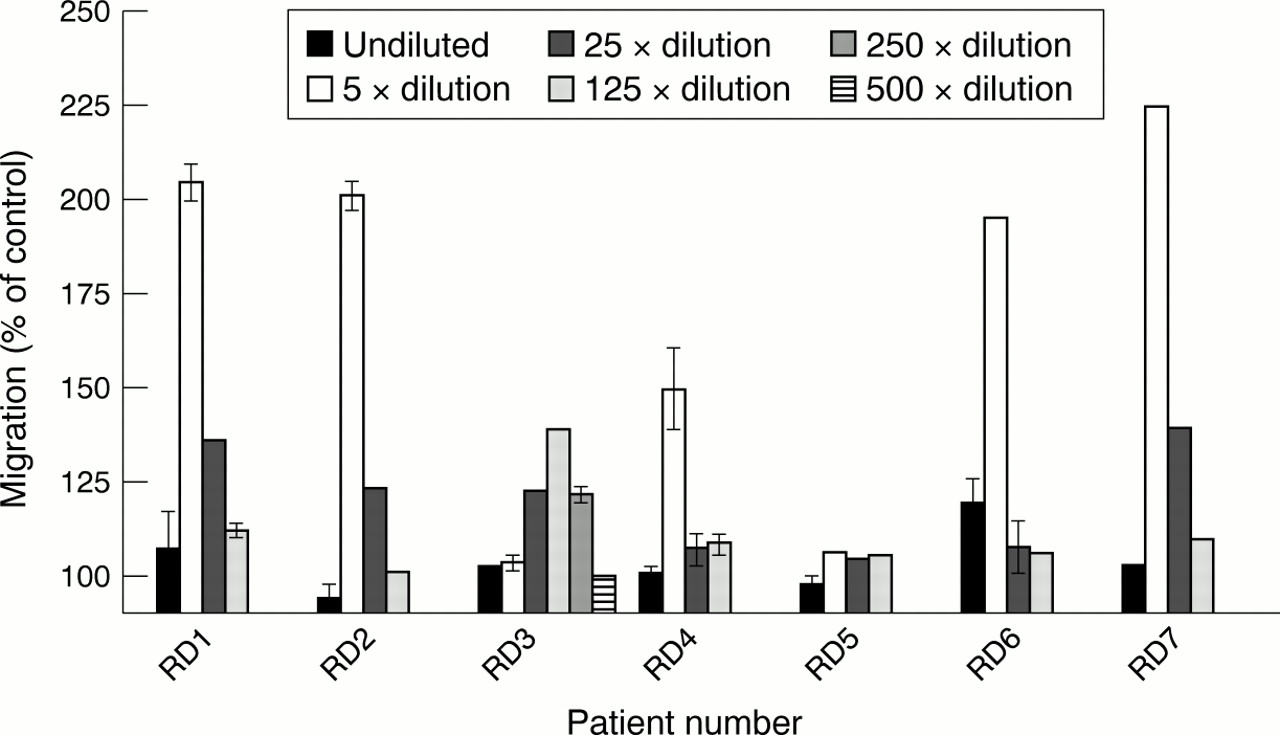
Migratory response of human cultured mast cells to synovial fluid. The fluid was diluted in medium and tested for mast cell chemotactic activity. Dilutions: undiluted, 5×, 25×, 125×, 250×, and 500×. Migration was measured after 150 minutes. The results depicted are from three experiments performed in triplicate. Results are given as means (SEM).
Comparison of mast cell migration in response to different chemoattractants
SCF AND TGFβ IN SYNOVIAL FLUID
We and others have earlier reported that SCF and TGFβ can induce mast cell migration in vitro.19-21 We therefore suggested that the mast cell chemotactic activity in synovial fluids might, at least in part, be attributed to these factors. The presence of SCF and TGFβ in synovial fluid was analysed by ELISA. Table 4 shows that SCF could be detected in all fluids tested. In contrast, TGFβ could not be detected in any of the fluids. However, TGFβ is effective as a chemotaxin at very low concentrations, with an ED50 of 10 fM.29 Thus TGFβ might be present in the synovial fluids at levels below the detection limit for the ELISA and still act as a chemoattractant for mast cells.
Measurements of transforming growth factor β (TGFβ) and stem cell factor (SCF) in synovial fluid
BLOCKING OF MAST CELL CHEMOTACTIC ACTIVITY WITH ANTIBODIES AGAINST SCF OR TGFβ
To test the involvement of SCF and TGFβ in the migration of mast cells towards synovial fluids, we used blocking antibodies against SCF and TGFβ. The antibodies were added to synovial fluids before the migration assay. Mast cell chemotactic activity was almost completely blocked in one of the fluids, while four were partially blocked and one not affected when treated with anti-SCF (fig 3A). Similarly, antibodies against TGFβ completely blocked the activity in one of the fluids, while three were partially blocked and two were not affected (fig 3B). The specificity of the antibodies and the lack of effect of an irrelevant antibody were also tested (data not shown).

Inhibition of activity of synovial fluids in mast cell chemotaxis assay by specific antibodies against (A) SCF (10 μg/ml) and (B) TGFβ (10 μg/ml). Open bars = control, filled bars = with antibody. The results are from three experiments performed in triplicate. Results are given as means (SEM). A significant effect was obtained as indicated: *p<0.05, ***p<0.001.
INHIBITION OF MAST CELL CHEMOTAXIS TO SYNOVIAL FLUIDS
To analyse further the nature of the mast cell chemotactic activity in synovial fluids we decided to treat the responder cells with different inhibitors which interfere with intracellular signalling. We chose to use pertussis toxin for inhibition of Gi protein coupled receptors (such as complement receptors) and genistein for tyrosine kinase receptors (for example, stem cell factor receptor). Each synovial fluid was used at a dilution giving optimal migration. Figure 4A shows that pertussis toxin treatment reduced the migratory response of the mast cells to several of the fluids. Of particular interest are fluids RD6 and RD7, which were unaffected by treatment with antibodies, but which did not induce migration of pertussis toxin treated cells. Genistein abolished the migratory response to fluid RD3 (fig 4B), which is the same fluid that showed SCF and TGFβ dependent migration (fig 3A and B). However, other fluids that also seemed to induce SCF and TGFβ dependent migration could still induce migration of genistein treated cells.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Analysis of signal transduction pathways which play a part in mediating mast cell chemoattractant activity in synovial fluids. Mast cells were treated with (A) pertussis toxin, or (B) genistein for 90 minutes before chemotaxis assay. The fluids were used at dilutions giving optimal migration. Open bars = control; filled bars = with inhibitor. The results are from three experiments performed in triplicate. Results are given as means (SEM). A significant effect was obtained as indicated: **p<0.01, ***p<0.001.
Discussion
Our findings show that (a) synovial fluids collected from patients with rheumatic diseases contain mast cell chemotactic activity, (b) SCF and TGFβ are expressed in the synovium and SCF can be detected in the synovial fluids, and (c) the mast cell chemotactic activity in the synovial fluids is due to multiple chemoattractants, of which SCF and TGFβ are two.
Rheumatoid arthritis is a chronic inflammatory disease characterised by synovial hyperplasia and hypertrophy. Only recently has the mast cell been recognised as an important effector cell of the rheumatoid lesion.6 One possible cause for the observed mast cell hyperplasia might be proliferation and maturation of resident immature mast cell precursors already present in the tissue. However, it was recently reported that mast cells in the synovial membrane of patients with arthritis do not proliferate.30 It is therefore likely that a direct migration of mast cells within tissues is an important mechanism for the increase in mast cell numbers evident in rheumatic diseases. Although mast cells have been considered to be stationary cells in the tissue, with low tendency to migrate, it is now recognised that they can migrate.31 We therefore proposed the hypothesis that cells in the synovium secrete factors that can attract mast cells to the synovium, and that mast cell chemotactic activity should be detectable in the synovial fluid. Indeed, six of seven tested synovial fluids contained mast cell chemotactic activity. As has been reported for lymphocytes, maximum activity was found at a 1:5 dilution of the fluids.32 Thus both lymphocyte and mast cell chemotactic activity can be detected in the fluids.
It was determined that two of the known mast cell chemotaxins, SCF and TGFβ1,20 ,21 were expressed in the synovial membrane, and contributed to mast cell chemotactic activity in the synovial fluids. The cells immunopositive for SCF in the synovial membrane were identified as synoviocytes (macrophages and fibroblasts). It was recently reported by Ceponis et al that SCF is expressed in the synovial membrane in arthritis.30Furthermore, the same group has reported that synovial fibroblasts release SCF in vitro that can induce mast cell migration.33 We confirm their results, but, in addition, we also show that SCF is secreted in vivo and can be detected in synovial fluid. Furthermore, our study points out the complexity of the mast cell chemotactic activity found in synovial fluid. Stem cell factor is probably one part of the activity, but many other factors, as discussed below, are also likely to play a part.
SCF is not only a mast cell chemotaxin but is also important for other mast cell functions, such as growth and differentiation, survival, adhesion, and activation.34 Thus the presence of SCF in the synovium may have important functions both for the activation and survival of the synovial mast cells and might thereby contribute to the increased number of mast cells in the synovium.
Expression of TGFβ has been described in the synovium of patients with rheumatoid arthritis, supposedly to be secreted into the synovial fluid.35 All three isoforms of TGFβ are expressed in arthritis.36 All the specimens analysed in this study expressed TGFβ1, TGFβ2, and TGFβ3. Although the isoforms have the same potency to induce migration of mast cells in vitro, TGFβ3 is the most effective.29 Mast cell hyperplasia in arthritis may therefore, in part, be due to differential expression of TGFβ isoforms in the synovial membrane.
The composition of synovial fluids is complex and it is therefore not surprising that blocking with antibody to single cytokines does not completely block the activity of the fluid. Cytokines other than those studied here have been reported to attract human mast cells, and still more may remain to be discovered.
One approach for delineating the different types of factors mediating mast cell chemotactic activity is to block different receptor signalling pathways using inhibitors. Pertussis toxin was used to block the family of inhibitory guanine nucleotide-binding proteins (Gi). We, and others, have recently reported that factors mediating their effects through receptors sensitive to pertussis toxin treatment, such as C3a, C5a, serum amyloid A, platelet activating factor, and interleukin 8, are mast cell chemotaxins.37-42 Some of these are expressed in the synovial membrane and are present in the synovial fluid.24 ,43 ,44 The response of pertussis toxin treated mast cells was reduced to all except one of the fluids (RD4), indicating the presence of chemotaxins mediating their effects through Gi protein coupled receptors. Similarly, inhibition of tyrosine kinases reduced the response to several of the fluids, even to those that showed a decreased chemotactic activity on pertussis toxin treated cells. These results strongly suggest that several factors are present in the synovial fluid that can act as mast cell chemoattractants. We have previously found a similar complexity of mast cell chemoattractants in other body fluids, including nasal lavage from allergic patients and bronchial lavage fluid from asthmatic patients.45 ,46
One complicating factor in the understanding of the control of cell migration is how different chemotaxins interact with each other. They may be additive or synergistic, but may also counteract each other. We determined that mast cell migration induced by platelet activating factor is attenuated by C5a and SCF (unpublished results).42 This might explain the migratory response to RD4 that is attenuated by anti-SCF, but is active on pertussis toxin as well as genistein treated cells. Furthermore, TGFβ1 has been shown to down regulate the response to SCF in several different systems,47 ,48 including migration (Sundström, Olsson, and Nilsson, manuscript in preparation). Although we have clearly shown that synovial fluids contain strong mast cell chemotactic activity, the cross talk and cross desensitisation among receptors for chemotaxins make it difficult to identify the factors mediating the response in each synovial fluid.
In conclusion, synovial fluids contain mast cell chemotactic activity that can have a role in the mast cell hyperplasia evident in rheumatic diseases. SCF could be detected in the synovial fluid, indicating that SCF may play a part not only in the recruitment of mast cells but also in the perpetuation of chronic inflammation by acting as a mast cell survival and activating factor. There have been reports of a correlation between clinical improvements and a decrease in mast cell numbers,14 ,49 and thus inhibiting mast cell migration might decrease the local mast cell hyperplasia and as a consequence reduce the destructive inflammatory process in arthritic diseases.
Acknowledgments
We thank Anna-Karin Kratz for excellent technical assistance, Drs Staffan Lindblad and Lollo Gröndal for patient material, Drs Lars Klareskog and Ulf Andersson for valuable discussions and critical reading of the manuscript, Dr BA Harris for linguistic advice, and Dr Keiko Funa for antibodies against TGFβ isoforms.
References
Footnotes
This study was supported by the Swedish Association against Rheumatism, the Swedish Foundation for Health Care Sciences and Allergy Research, Ollie and Elof Ericssons Foundation, and King Gustaf V 80-years foundation.