Article Text

Abstract
OBJECTIVE The purpose of this study was to investigate the therapeutic potential of the anti-CD2 mAb OX34 first with regard to bone protection in established rat adjuvant arthritis (AA) and secondly with regard to prevention of AA induction.
METHODS Established AA was treated with dexamethasone (1 mg/kg body weight) for two days plus OX34 mAb or control mAb over three days (2 mg and then 1 mg) starting at different time points of the disease. For prevention studies animals were injected as above with mAb before induction of AA. Arthritis score (AS), hindpaw thickness, and body weight were blindly measured three times per week. Flow cytometry and hindpaw radiography were performed at the end of the study (day 29).
RESULTS Treatment of early AA with OX34 mAb combined with dexamethasone but not dexamethasone plus control mAb dramatically suppressed established AA as assessed by AS and hind paw thickness (>65% and >80% reduction, respectively; p < 0.05). Most importantly, early treatment in the course of AA almost completely prevented bone destruction in established AA. When given before AA induction OX34 alone prevented the initiation of arthritis compared with controls (AS reduction 83-95%, p < 0.05). In addition, OX34 plus dexamethasone treatment resulted in depletion of CD4+ T cells but not CD8+ T cells. IL2R+ and CD45RC-(‘memory’) T cells were significantly reduced.
CONCLUSIONS Anti-CD2 mAb treatment prevents AA induction confirming the role of CD4+ T cells in the induction phase of AA. In addition, early OX34 plus dexamethasone treatment resulted in pronounced clinical improvement and joint protection. OX34 treatment therefore inhibits the initiation and the perpetuation of rat AA.
- adjuvant arthritis
- mAb therapy
- CD2
- bone destruction
Statistics from Altmetric.com
Rheumatoid arthritis (RA) is characterised by mononuclear cell infiltrates in the synovium leading eventually to joint destruction. Rat adjuvant arthritis (AA) serves as an animal model for RA. Although the clinical course of AA is somewhat different it shares important features with RA—that is, polysynovitis, characteristic histological findings, and ultimately erosive bone destruction.1 ,2 In addition, the clinical response to anti-inflammatory agents is similar in RA and established AA. Therefore, this model has been widely used for therapeutic and pathogenetic studies of chronic forms of arthritis.1 ,3-6
Currently available anti-rheumatic agents (that is, corticosteroids, non-steroidal anti-inflammatory drugs (NSAIDs), and disease modifying anti-rheumatic drugs (DMARDs)) are known for their potent anti-inflammatory effectiveness. However, none of these drugs are able to stop joint destruction, although some DMARDs, particularly methotrexate and parenteral gold, have been shown to somewhat slow disease progression.7-9 New therapeutic strategies for chronic forms of arthritis have to aim at both, suppression of inflammation and bone protection. To achieve these goals it is important to understand the different stages of disease progression and to identify relevant targets—that is, most relevant cell type(s) or cytokine(s), or both.
We previously reported on the anti-inflammatory effects of an anti-CD2 mAb OX34 in established AA.10 This mAb was found to deplete transiently CD4+ but not CD8+ T cells. We now investigate whether treatment with OX34 has an influence on bone destruction as assessed by radiography and, in an attempt to further understand different phases of arthritis development, whether OX34 mAb treatment can prevent the induction of arthritis. Our data demonstrate that OX34 almost completely prevents AA development and, more importantly, when used with a corticosteroid pulse treatment effectively suppresses established AA leading to joint protection. Therefore, OX34 treatment interferes with the initiation and perpetuation of this chronic form of arthritis.
Methods
ANIMALS
Inbred female Lewis rats were obtained from the Abteilung Versuchstierkunde of the Medizinische Hochschule. The animals were kept under specified pathogen free conditions, were housed at two to three animals per cage, and fed standard rodent chow and water ad libitum.
MAB, DEXAMETHASONE
The hybridoma cells producing mouse anti-rat CD2 mAb (OX34, IgG2a) were kindly provided by A N Barclay, University of Oxford, UK, the mouse antihuman LFA-3 mAb AICD58.6 (IgG2a) was used as control mAb (a gift from S C Meuer, Heidelberg, Germany). Purified mAb was produced in spinner flask followed by affinity chromatography using protein A sepharose beads. Purified mAb were filtered (0.25 μm) and administered intraperitoneally. OX34 was given at a loading dose of 2 mg followed by 1 mg on the subsequent two days unless stated otherwise. Dexamethasone was diluted in phosphate buffered saline (PBS) giving a dose of 1 mg/kg body weight and injected intraperitoneally.
INDUCTION OF AA, CLINICAL AND RADIOGRAPHIC ASSESSEMENT
AA was induced by intradermal injection into the tail base with 0.1 ml of FCA containing 10 mg/ml Mycobacterium tuberculosis (R73Ra) diluted in incomplete Freund’s adjuvant (both Difco Laboratories, Detroit, MI, USA). AA developed between day 10 and 15. The severity of arthritis was evaluated by a blinded investigator (JCH or JW) three times per week on a scale of 0 to 4 per limb based on increasing swelling and periarticular erythema as previously described.10 The maximum arthritis score (AS) was therefore 16. At the same time the body weight was measured. Hindpaw thickness was measured as previously described using calipers.11 Established AA was arbitrarily defined as clear arthritis (AS per limb ⩾ 1) in at least two limbs (total AS ⩾ 2). Using these criteria 75% of all animals had established AA by day 12 and 100% by day 15. At the end of the study the animals were killed and both hindlimbs were amputated, frozen at −80°C, and radiographed using mammography technique at 20 mAs. Radiographic scoring was performed by assessing soft tissue swelling, periosteal new bone formation, joint space narrowing, periarticular osteoporosis, and bone destruction on a scale of 0 (normal) to 3 (maximum) per hindlimb.12 Scoring was performed by two independent, blinded investigators (JCH and CH). The maximum radiographic score was 6 per animal.
SUBSET ANALYSIS OF BLOOD LYMPHOCYTES BY FLOW CYTOMETRY
Peripheral blood was obtained by venopuncture from the retro-orbital plexus. Numbers of leucocytes were determined in duplicate samples using a Coulter counter and three differential blood smears were prepared. Red blood cells were lysed using a hypotonic solution as previously described.13 Hybridoma cell lines were kindly provided by A N Barclay, University of Oxford, UK, and T Hünig, University of Würzburg, Germany. Double staining was performed using pairs of biotinylated or FITC conjugated mouse anti-rat mAb and streptavidin red 670 as secondary antibody giving the following lymphocyte subsets10: B lymphocytes (κ light chain+, OX12), CD4+ T cells (W3/25high) CD8+ T cells (R73+, OX8+), natural killer cells (R73-, OX8+), monocytes (W3/25low), CD45RC+ CD4+ T cells (W3/25high, OX22high), CD45RC- CD4+ T cells (W3/25high, OX22low). Interleukin 2 receptor expression was determined by a single staining technique using OX39, and goat antimouse polyclonal antibodies coupled to FITC. MAb and isotype matched irrelevant antibodies served as controls. Only viable cells as determined by forward and side scatter characteristics were analysed using a FACScan flow cytometer. Absolute subset numbers were calculated using leucocyte counts, differential blood count, and percentage of subsets as determined by flow cytometry.
STATISTICAL ANALYSIS
All measurements are shown as mean (SEM) unless otherwise stated. To compare two different groups the Mann-Whitney U test was applied. Differences were considered significant if the p value was ⩽0.05. Three group comparison was done by the Kruskal-Wallis test followed by pairwise comparison using the Mann-Whitney U test if the Kruskal-Wallis test was significant (significance level ⩽ 0.05). For comparison of radiographic scores (comparison of seven groups) first the Kruskal-Wallis test was performed and, if significant, the multiple comparison procedure by Tukey-Kramer, which controls experiment wise error rate.14
Results
SUPPRESSION OF ESTABLISHED AA BY COMBINED TREATMENT WITH THE ANTI-CD2 MAB OX34 AND DEXAMETHASONE
We have previously shown that OX34 treatment has an anti-inflammatory effect on established AA.10 However, this treatment does not affect bone destruction as demonstrated by similar radiographic scores for OX34 or control mAb treated AA rats (fig 1). In an attempt to achieve joint protection we combined OX34 treatment with a one day dexamethasone pulse. Indeed, a strong reduction of the AS was found in the OX34 plus dexamethasone treated group compared with dexamethsone plus control mAb treated group (each group n = 5, p < 0.05 from day 17 to day 27)(fig 2). However, the reduction of bone destruction was not significant when compared with AA rats receiving dexamethasone plus control mAb (fig 1).

Influence of dexamethasone or OX34 treatment, or both, on radiographic bone destruction of AA rats. Indicated is the mean radiographic score of both hindpaws, the SEM for each treatment group, and the levels of significance. Radiographic scores of two groups of 14 rats receiving either OX34 or control mAb are included from a previous study (Hoffmann et al, unpublished data). All mAb treatments were started with 2 mg followed by 1 mg intraperitoneally per injection day, mAb were given over three consecutive days unless stated otherwise; dexamethasone was given intraperitoneally at 1 mg/kg body weight either for two consecutive days starting on day 12 or 18 or once on day 15. * = mAb given two days and one day before induction of AA and on the day of induction. † = mAb given on day 15, 17, 20, and 23 after induction of AA.
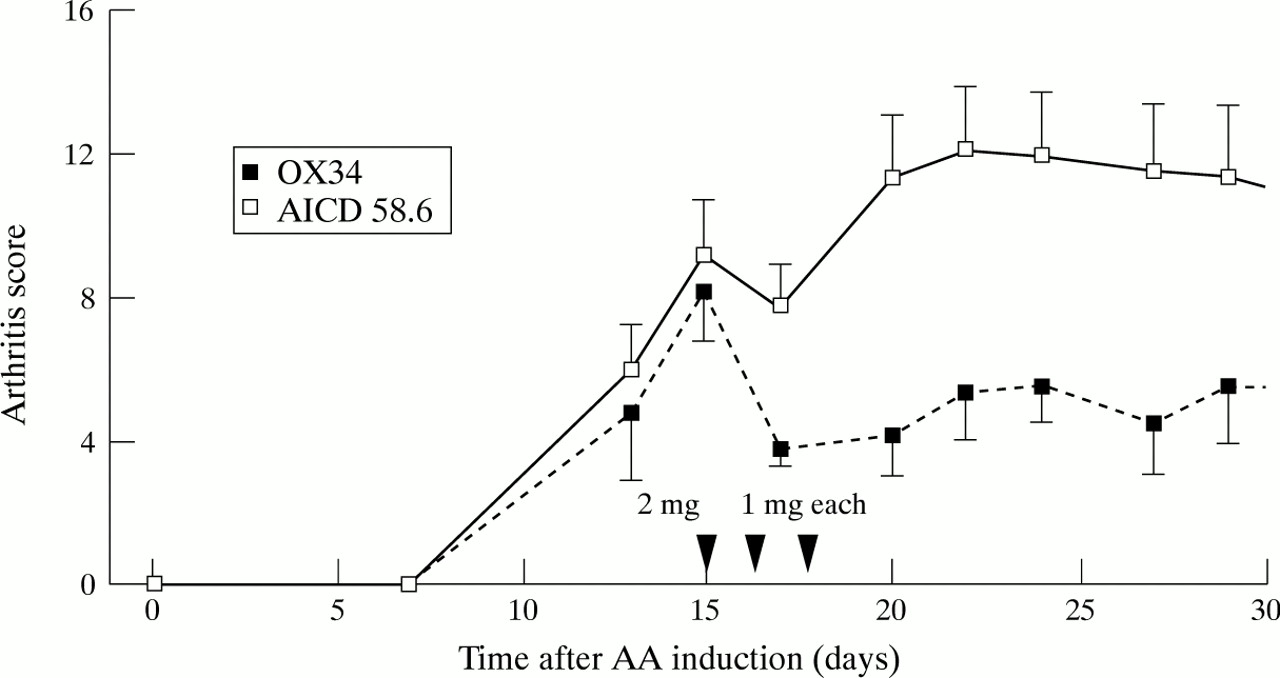
Amelioration of established AA by dexamethasone plus OX34 mAb therapy. Ten animals with established AA were randomised to intraperitoneal injections of OX34 (n = 5) or istotype matched control mAb (AICD58.6, n = 5) over three consecutive days starting with a loading dose of 2 mg followed by 1 mg each at indicated time points (arrows). Dexamethasone was given intraperitoneally on day 15 at a dose of 1mg/kg body weight. Arthritis score was measured by a blinded investigator at indicated time points. Shown are means (SEM), arrows indicate mAb injections.
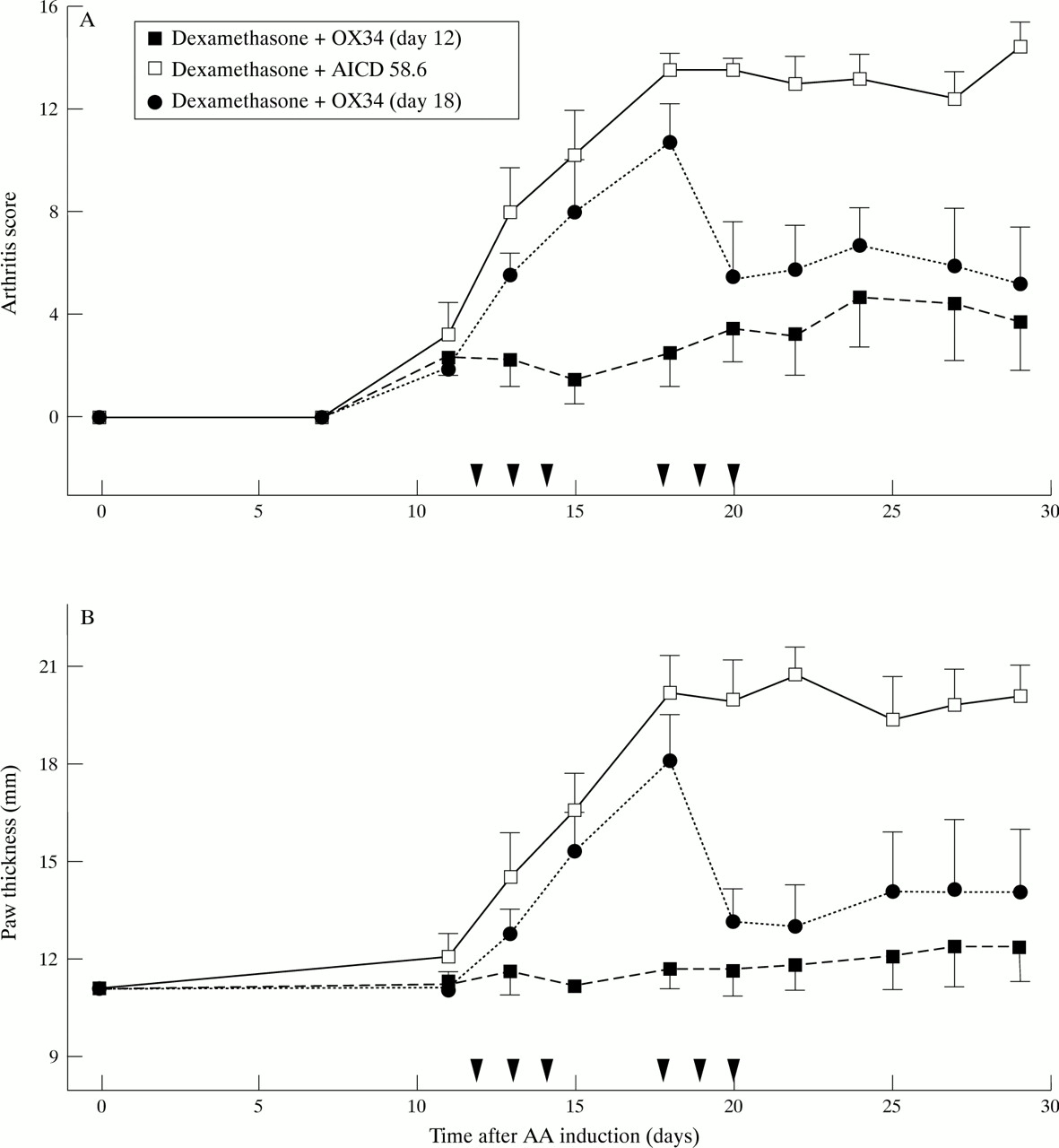
To further improve the effect on bone destruction we next modified the treatment protocol by giving dexamethasone on two consecutive days and OX34 starting just after the onset of arthritis—that is, day 12 after AA induction. This modification resulted in almost complete joint protection (mean radiographic score 0.25 versus 3.75, p < 0.05)(fig 1) and significant clinical improvement (AS and hindpaw thickness) when compared with a control group receiving dexamethasone over two days plus control mAb starting on day 12 (fig 3A and B). When OX34 plus dexamethasone was started on day 18 (same regimen as above) the reduction of AS and hindpaw thickness was also significant compared with dexamethasone plus control mAb starting on day 12 (p < 0.05), however the reduction in bone destruction was not (mean radiographic score 1.25 versus 3.75, p = 0.08). As no radiographic difference was found between AA rats treated with dexamethasone plus control mAb on day 12 or day 15 these animals were pooled in fig1.
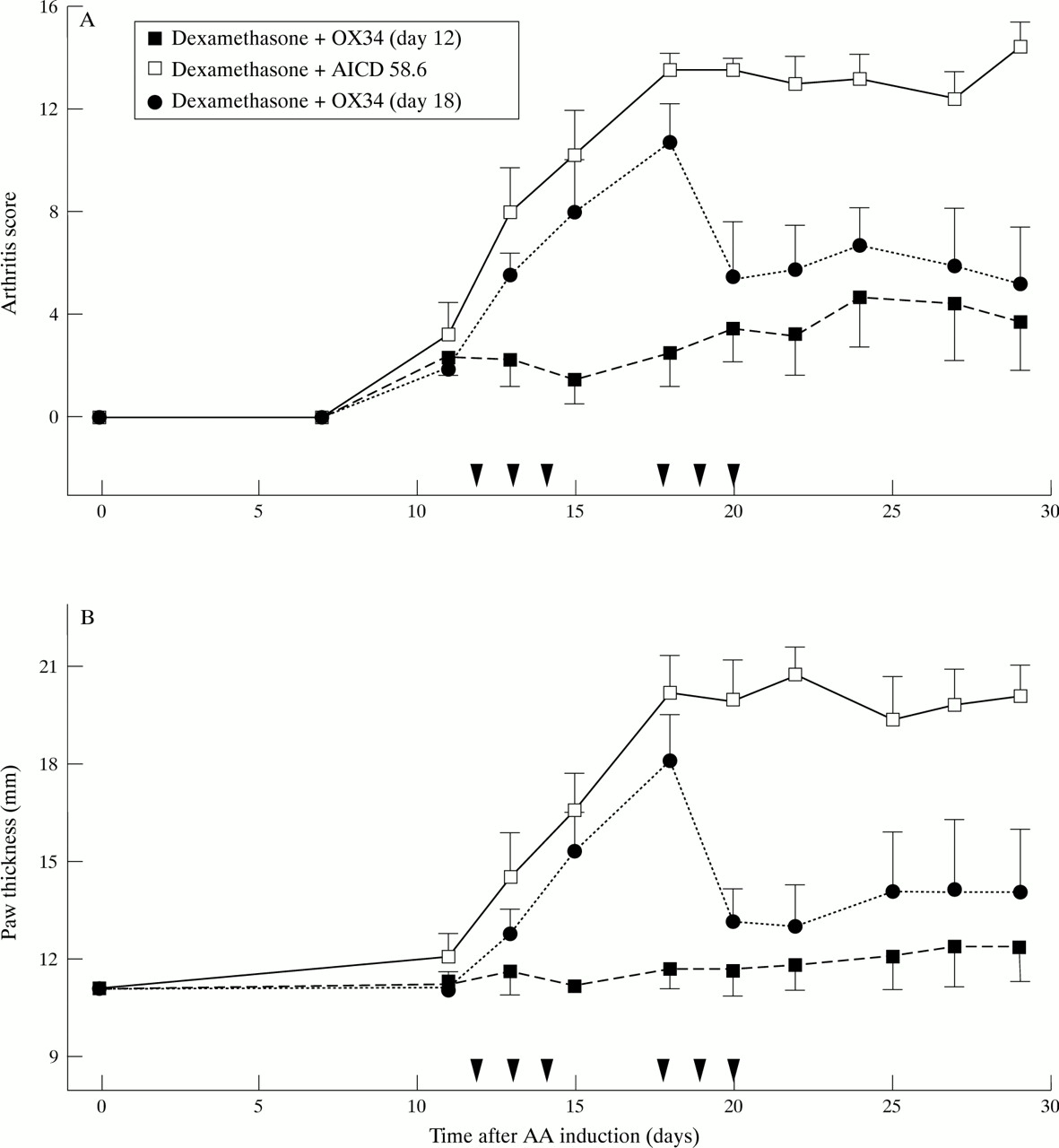
Marked suppression of established AA by early, repeated OX34 mAb injections plus a dexamethasone pulse over two days. Sixteen rats were injected with FCA and 12 rats developed AA by day 12 (AS ⩾ 2). These were randomised into three groups, four animals each, and received blindly either dexamethasone plus OX34 starting on day 12 (n = 4), dexamethasone plus control mAb (AICD 58.6) starting on day 12 (n = 4), or dexamethasone plus OX34 starting on day 18 (n = 4). PBS was given on those days when no mAb was injected (for example, day 12 to 14 for dexamethasone plus OX34). Dexamethasone diluted in PBS was given at a concentration of 1 mg/kg body weight on two consecutive days, mAbs were injected for three consecutive days starting with 2 mg followed by 1 mg per day. Arthritis score (A) and hindpaw thickness (B) were measured by a blinded investigator at indicated time points. Symbols and labelling as in figure 2.
OX34 PLUS DEXAMETHASONE TREATMENT REDUCES CD4+ T CELLS AND ACTIVATED T CELLS IN PERIPHERAL BLOOD
OX34 treatment without dexamethasone was previously found to deplete CD4+ but not CD8+ T cells.10 A similar effect was found after treatment with OX34 plus dexamethasone starting on day 12. Therefore, flow cytometry at the end of the study (day 29) showed depletion of CD4+, but not NK cells and CD8+ T cells (table 1). Dexamethasone and control mAb had no significant effect on CD4+ T cell concentrations (data not shown). The following leucocyte subsets were found to be unchanged: neutrophils (control: 4847 (511)/μl; OX34: 5766 (896)/μl), monocytes (control: 781 (60)/μl; OX34: 644 (120)/μl), and B lymphocytes (control: 1175 (168)/μl; OX34: 1523 (141)/μl). No differences were seen between early and late OX34 treatment (data not shown).
Effect of OX34 and dexamethasone administration on peripheral blood leucocyte subsets on day 29
When looking at activation markers after combined dexamethasone plus OX34 therapy starting on day 12, the concentrations of CD45RC- CD4+ (‘memory’) T cells and IL2R+ PMNC were significantly reduced (table 1). Different starting points of OX34 treatment gave similar results when looking at these leucocyte subsets (data not shown). Reduction of numbers of CD4+ T cells, ‘memory’ T cells, or IL2R+ PMNC therefore did not correlate with the degree of bone protection (data not shown).
PREVENTION OF AA BY TREATMENT WITH THE ANTI-CD2 MAB OX34
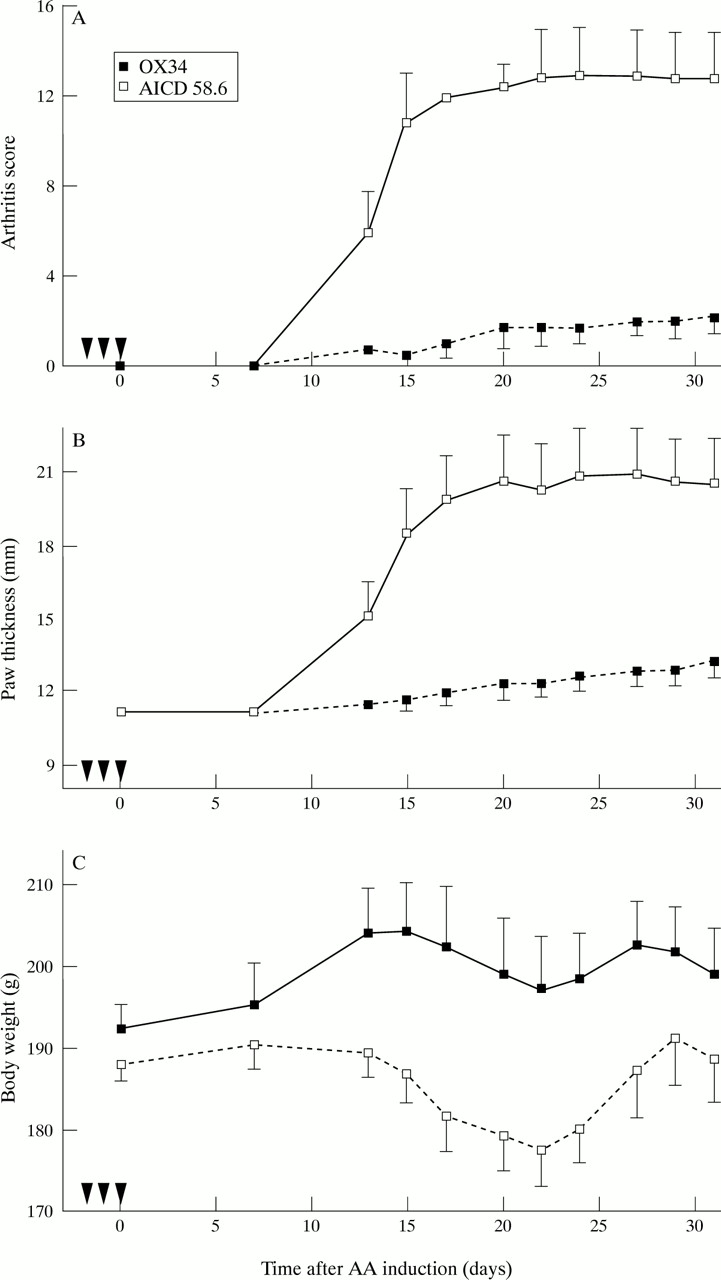
As early dexamethasone plus OX34 mAb treatment suppressed the perpetuation of AA we asked next whether OX34 alone might inhibit the initiation of AA. Twelve female rats received blindly either OX34 or isotype matched control mAb starting two days before AA induction. While all animals receiving the control mAb developed severe polyarthritis by day 13 only two of six rats that had received OX34 mAb had mild arthritis on day 13 (fig 4A). Over the whole observation period the highest mean AS of the OX34 treated group was 2.2 compared with 12.8 in the control group. Similarly, hind paw thickness was significantly reduced and body weight increased in the OX34 treated group in comparison with the control group (fig 4B and 4C). In addition, OX34 treatment drastically reduced bone destruction as judged by hind paw radiography on day 31 (fig 1). No difference was found between animals receiving control mAb before AA induction or on day 15 after AA induction and were therefore pooled in figure 1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Prevention of AA induction by the anti-CD2 mAb OX34. Twelve animals were randomised to intraperitoneal administration of OX34 (n = 6) or istotype matched control mAb (AICD 58.6, n = 6) as in figure 3 before AA induction. Arthritis score (A), hind paw thickness (B), and body weight (C) was measured by a blinded investigator at indicated time points. Symbols and labelling as in figure 2.
Discussion
Three important findings are described in this report: (1) Dexamethasone pulse therapy over two days plus OX34 over three days effectively suppressed synovitis in established AA when used early in the disease process. More importantly, dexamethasone plus early OX34 therapy prevented bone destruction in established AA. (2) In addition to its clinical effectiveness, OX34 plus dexamethasone reduced activated peripheral blood T cells as assessed by IL2 receptor and CD45RC expression. (3) The anti-CD2 mAb OX34 almost completely prevents the induction of AA.
Similar to human RA, the development of AA can be devided into three phases starting with the induction phase without evidence of synovitis, followed by early synovitis, and finally late synovitis with progressive joint destruction. Our data suggest that T cells and CD4+ T cells in particular are very important in the initiation phase of AA as OX34 selectively depletes CD4+ T cells resulting in almost complete prevention of AA formation. This finding confirms the notion that CD4+ rather than macrophages or fibroblasts are of primary importance for the induction of AA. Still, some groups regard macrophages and pannocytes as initiating cell type in chronic forms of arthritis.15 ,16However, several lines of evidence suggest that T cells, and CD4+ T cell in particular, are the most important cell type in the initiation phase of chronic forms of arthritis. CD4+T cells are among the first cells to reach inflamed joints.6 Neonatal thymectomy or depletion by mAb therapy of T cells or of CD4+ T cells prevents AA induction in Lewis rats.4 ,17 ,18 Therefore, the driving force in the initiation phase of AA, and probably other chronic forms of arthritis, seems to be the CD4+ T cell subset.
In the second phase of AA, early synovitis, the joint becomes infiltrated by macrophages and neutrophils. Accordingly, depleting T cells or even CD4+ T cells by mAb therapy only ameliorates AA for a short period of time—that is, at most one week.4 ,19 In contrast, amelioration of established AA by OX34 and dexamethasone lasts for at least 2.5 weeks when given at an early stage. As dexamethasone is known to inhibit activation and immigration of monocytes and neutrophils5 ,20 ,21 it can be speculated that dexamethasone over two days blocks macrophage and neutrophil infiltration while OX34 targets T cells. These T cells seem to be the driving force in this phase of inflammation because dexamethasone over only two days with the control mAb had no significant effect on AA. This hypothesis is supported by data from Issekutz et al showing that dexamethasone inhibits migration of monocytes and neutrophils but not T lymphocytes to inflamed joints.5 Attacking both, T cells and inflammatory mediators, has previously been reported to be highly effective in established collagen arthritis using anti-TNFα and anti-CD4 mAb.11 Similarly, our approach to target CD4+T cells by OX34 and macrophages/neutrophils by dexamethasone effectively suppressed established AA leading to joint protection. As a result of this immunotherapy peripheral activated T cells were found to be reduced.
The last phase of AA is late synovitis, which is characterised by progressive joint destruction. Although affected joints are infiltrated by mononuclear cells and predominantly CD4+ T cells this phase is dominated by activated macrophages, which destroy the cartilage and stimulate fibroblasts and chondrocytes leading to progressive joint destruction and pannus formation. Therefore, important damage has already occurred in late synovitis. This is reflected in our experiments by some bone destruction when dexamethasone plus OX34 is given at a late stage of disease. Still, the anti-inflammatory effect of this late treatment was almost equivalent to early treatment suggesting that T cells also participated in the maintenance of synovitis during this phase of disease. The anti-inflammatory effect and the albeit non-significant reduction of bone destruction by the late OX34 plus dexamethasone treatment underline, despite the small sample size of four rats per group, the importance of attacking both T cells as the driving force and macrophages plus neutrophils as the effector cells even in late AA. As joint protection plus suppression of synovitis are the ultimate goals of a better RA treatment than the currently available DMARDs future studies have to investigate CD2 directed immunotherapies in human RA.
Different mechanisms might be responsible for the clinical effects of OX34 therapy. Certainly the most obvious effect of the OX34 therapy is the depletion of CD4+ T cells but not CD8+ T cells. This depletion might explain why OX34 prevents the induction of AA. Secondly, OX34 was shown to interfere with T cell activation by inhibition of IL2 secretion, a finding that has also been reported for anti-CD2 mAb in the human system.22 ,23 Thirdly, OX34 inhibits the CD2/CD48 interaction by two mechanisms: (1) OX34 blocks the CD2/CD48 interaction directly by binding to the CD48 binding epitope on the CD2 molecule.24 (2) OX34 leads to longlasting downmodulation of the CD2 molecule on CD4+ T cells.10 As multiple immune functions such as macrophage activation, B cell differentiation, and T cell activation are mediated via the CD2/CD48 interaction25-28 blocking this interaction might well explain the anti-inflammatory effect of this mAb. To this end, treatment with OX34 not only suppresses AA but also induces tolerance in a rat heart transplant model,22prevents transfer of diabetes,29 and suppresses established experimental autoimmune encephalomyelitis.30
In summary, the anti-CD2 mAb OX34 prevents AA induction emphasising the importance of T cells, particularly CD4+ T cells, in the induction phase of AA. In addition, combined OX34 plus dexamethasone treatment suppresses established AA and, at least when used in early synovitis, prevents joint destruction. Therefore, OX34 plus dexamethasone interferes with the maintenance of arthritis showing that T cells and mainly CD4+ T cells perpetuate the arthritic process. Further studies are needed to investigate the mechanism of action of OX34, the long term efficacy of this treatment, and to evaluate whether a CD2 directed immunotherapy might be suitable for human RA.
Acknowledgments
This study was supported by SFB grant 244-A7 to JW, DFG-grant Ho 1561/1-2 to JCH, and by an award to JCH from the Kurt-Eberhard Bode foundation. The authors thank K Bankes-John, I Dressendörfer, and F Weidner for technical assistance with flow cytometry, differential blood counts, and immunohistology. Also, we are grateful to T Krüger for excellent animal care and C Falkenstein for help in performing radiographs. We kindly acknowledge the help of J König in performing statistical analysis.
Some of these results were presented at the Annual meeting of the Deutsche Gesellschaft für Immunologie in Hamburg, Germany, 1996.