Article Text

Abstract
BACKGROUND Butyrate oxidation within the colonocyte is selectively inhibited by hydrogen sulphide, reproducing the metabolic lesion observed in active ulcerative colitis.
AIMS To study generation of hydrogen sulphide by sulphate reducing bacteria (SRB) and the effects of 5-aminosalicylic acid (5-ASA) in patients with ulcerative colitis in order to identify a role of this noxious agent in pathogenesis.
PATIENTS Fresh faeces were obtained from 37 patients with ulcerative colitis (23 with active disease) and 16 healthy controls.
METHODS SRB were enumerated from fresh faecal slurries and measurements made of sulphate reducing activity, and sulphate and hydrogen sulphide concentrations. The effect of 5-ASA on hydrogen sulphide production was studied in vitro.
RESULTS All controls and patients with active ulcerative colitis carried SRB and total viable counts were significantly related to the clinical severity grade. SRB were of two distinct types: rapidly growing strains (desulfovibrios) which showed high sulphate reduction rates, present in 30% of patients with ulcerative colitis and 44% of controls; and slow growing strains which had little activity. In vitro, 5-ASA inhibited sulphide production in a dose dependent manner; in patients with ulcerative colitis not on these drugs faecal sulphide was significantly higher than in controls (0.55 versus 0.25 mM, p=0.027).
CONCLUSIONS Counts and carriage rates of SRB in faeces of patients with ulcerative colitis are not significantly different from those in controls. SRB metabolism is not uniform between strains and alternative sources of hydrogen sulphide production exist in the colonic lumen which may be similarly inhibited by 5-ASA. The evidence for hydrogen sulphide as a metabolic toxin in ulcerative colitis remains circumstantial.
- colitis
- sulphate
- sulphide
- bacteria
- fermentation
- salicylate
Statistics from Altmetric.com
The aetiology of ulcerative colitis is unknown but it is likely to depend on an interaction between genetic and environmental factors in a susceptible host.1 The commensal bacteria of the large intestine have become increasingly recognised as an essential component in the genesis of colonic epithelial inflammation in animal models of colitis.2-6 The normal flora are responsible for the fermentative breakdown of dietary and endogenous carbohydrate that has escaped digestion in the small intestine, the principal products being short chain fatty acids (SCFA) which are essential to the metabolism of the colonic epithelial cell and maintenance of normal mucosal function.7 Butyrate has a specific role as an energy source in healthy colonic epithelium,8-10 and evidence from in vitro11-14 and in vivo15 16 studies, albeit somewhat conflicting, indicates that oxidation of butyrate might be impaired in ulcerative colitis. This may account for the apparent benefit of SCFA enema therapy in refractory distal colitis in several small clinical trials,17 perhaps by overcoming mucosal “energy deficiency” in ulcerative colitis to induce disease remission. A recent sizeable controlled trial of SCFA enema therapy, however, only showed clinical efficacy in subsets of patients with distal ulcerative colitis18 and so their therapeutic place remains unsettled.
In vitro, butyrate oxidation in colonocytes can be selectively inhibited by hydrogen sulphide, especially in cells from the distal colon, reproducing the biochemical lesion reported in ulcerative colitis.19 20 In vivo, the application of hydrogen sulphide to rat colonic epithelium impairs metabolic performance,21 but the production of mucosal inflammation has been inconsistent between studies.21 22 Hydrogen sulphide has a number of other potentially adverse effects in the colon which indicate a possible role in disease pathogenesis,23and it is derived from the metabolism of gut bacteria. The sulphate reducing bacteria (SRB) are a diverse group of anaerobes which are uniquely capable of utilising the sulphate or sulphite ion as an electron acceptor in the dissimilation of organic substrates,24 principally SCFA or molecular hydrogen, produced during fermentation.25 SRB have been identified in human colonic contents26 and are an important source of hydrogen sulphide production within the lumen.27 Although SRB are not considered pathogenic,Desulfovibrio spp. have been isolated from pyogenic liver abscess material in humans. Moreover, the pig, ferret, and some other species, suffer an enteropathy that presents as watery diarrhoea with blood, anorexia, and weight loss and which can become chronic. In the pig it primarily affects the ileocaecal region and in the ferret the large bowel, extending proximally from the rectum. Histological changes include epithelial hyperplasia, goblet cell depletion, and crypt abscesses. The condition is caused by an intracellular Gram negative bacterium which is phylogenetically linked to SRB.28 29
We have therefore investigated the growth and activity of SRB in faeces in patients with ulcerative colitis and their influence on luminal hydrogen sulphide concentration in order to identify a possible relation with the diseased state. A secondary aim of the study was to determine whether drugs containing 5-aminosalicylic acid (5-ASA), a luminally active treatment in ulcerative colitis,30 31have an effect on faecal sulphide production in vitro and in vivo.
Methods
SUBJECT SELECTION
Thirty three consecutive patients with ulcerative colitis (19 men, 14 women; median age 46 years) were recruited for study while undergoing treatment at Addenbrooke's Hospital, Cambridge. Four patients were studied sequentially while in disease relapse and subsequent remission. Thirty seven separate observations were made in 23 patients with clinically active disease of mild to moderate severity according to the Truelove and Witts criteria,32 and in 14 patients in clinical, sigmoidoscopic, and histological remission as defined by Ruddell et al.33Seventeen patients had extensive colitis and 16 had left sided disease. Histological examination of rectal biopsy specimens was consistent with ulcerative colitis in all patients studied. All stool cultures were negative for pathogens and no patient had received antimicrobial agents for at least four weeks prior to inclusion in the study. Current drug treatment with corticosteroids and/or 5-ASA preparations was documented and enema medication omitted for at least 24 hours prior to faecal sampling. Eight of the patients with ulcerative colitis were breath methane positive, as defined by Segal et al.34
The control group consisted of 16 healthy volunteers recruited from laboratory personnel at the MRC Dunn Clinical Nutrition Centre, Cambridge (10 men, six women; median age 32 years; four breath methane positive) who were not receiving any medication at the time of study and had not received antimicrobial agents for at least four weeks prior to recruitment. Seven of these controls were later selected on the basis of high sulphate reducing activity (SRA) in their faeces (median sulphate reduction rate 12.2 nmol/day/g wet weight) and positive culture for rapidly growing SRB (median total viable counts about 108/g wet weight faeces) for in vitro study of the effects of 5-ASA on dissimilatory sulphate reduction in faeces.
Approval for the study was granted by the Ethical Committee of the MRC Dunn Clinical Nutrition Centre.
BREATH METHANE
Breath methane reflects carriage of methanogenic archaea in the colon and is a major route for hydrogen disposal in anaerobic systems. Duplicate end expiratory breath samples were collected into 20 ml plastic syringes equipped with a three way tap. Methane concentration was measured within 24 hours by gas chromotography using a Pye 104 series gas chromatograph equipped with a molecular sieve column and ionisation detector. Peaks were calibrated with a 5 ppm standard (BOC) and measured on a chart recorder. A sample of room air was taken at the same time as each breath collection and the value for methane, usually between 1.7 and 2.2 ppm, subtracted from the mean of the duplicate breath samples. Methane excretors were defined as those with more than 1 ppm of methane in breath above ambient air.
PREPARATION OF FAECAL SAMPLES
Faecal slurries (10% wt/vol) were prepared by homogenising freshly voided faecal samples with oxygen free potassium phosphate buffer (0.1 M, pH 7.0 in deionised water (18.2 MΩ)) in a Colworth 3500 stomacher and then passing through a 500 μm stainless steel mesh sieve, diameter 210 mm (Endecotts Ltd) using only gravity for filtration.
ENUMERATION OF SULPHATE REDUCING BACTERIA IN FAECES
Viable populations of SRB were enumerated in triplicate using autoclaved Postgate's medium E24 adapted for faecal inocula using the anaerobic agar shake tube serial dilution technique as described by Macfarlane and Gibson.35 The substrates used were acetate, lactate, propionate, and butyrate in combination, as previous studies have shown that these are the major electron donors that support the growth of most SRB species in faeces.25The dilution tubes were incubated at 37°C until individual colonies of SRB could be counted. The detection limit for SRB was ⩾102 colony forming units (CFU) per g wet weight faeces. Viable counts of SRB are expressed as the mean of three values of log10CFU/g wet weight faeces.
DETERMINATION OF FAECAL SULPHIDE
Total acid volatile sulphide concentration was measured using a modification of the methylene blue method36 adapted for the analysis of faecal slurry samples in zinc acetate solution (final faecal dilution 1/200) as described by Strocchi et al.37 Concentrated HCl and a mixing time of one hour was used for sample blanks and 1 ml of sample was passed through 0.2 μm 13 mm diameter cellulose acetate filter (Whatman, catalogue no. 6771-1302) using a 1 ml syringe, after the full development of methylene blue, to remove residual turbidity. The optical density of the samples was determined after one hour at a wavelength of 670 nm in an ultraviolet scanning spectrophotometer (PU 8740, Philips). The concentration of sulphide in the faecal slurries was expressed as μmol/g wet weight faeces and corrected for recovery. The coefficient of variability of the assay was 8.6%, and mean recovery of sulphide 81.6%. The absorbance assay was linear over the physiological range for sulphide in undiluted faeces.
DETERMINATION OF SULPHATE REDUCTION RATE IN FAECES
The rates of dissimilatory sulphate reduction in faeces were measured by an adaptation of the method of Jørgensen.38Aliquots (8 ml) of faecal slurry from each stool were pipetted into five separate straight sided glass specimen tubes (Orme) and a trace quantity (5 μCi in 5 μl) of sodium 35S-sulphate (ICN Biomedicals) injected into each using a 10 μl capacity microsyringe (Terumo, Japan). Each tube was sealed from the air with a silicon rubber bung (Fisons). Two tubes were immediately frozen at −25°C to provide blanks and three sample tubes were incubated for 18 hours at 37°C prior to freezing at −25°C. For each subject, five frozen faecal samples were subsequently thawed after transfer under separate streams of oxygen free nitrogen in the reaction vessels of a custom made distillation apparatus (Soham Scientific, Ely), as described by Macfarlane and Gibson.35 Acid volatile hydrogen35S-sulphide gas was distilled from the samples over a one hour period and trapped in oxygen free 120 g/l zinc acetate solution. Duplicate 0.5 ml aliquots from the five distillates and slurry samples remaining in the reaction vessels were added to 3 ml scintillation fluid (Ultima-Gold AB, Canberra Packard) and counted in a β emission liquid scintillation analyser (1900TR, Canberra Packard) with internal quench correction, colour, and heterogeneity monitoring, and corrected for background. Sulphate reduction rate was calculated as described by Macfarlane and Gibson35 and expressed as nmol sulphate reduced/g wet weight faeces/day using free faecal sulphate values.
In order to study the effect of 5-ASA on sulphate reducing activity, eight 8 ml aliquots of faecal slurry from each of seven healthy subjects with high SRA and rapidly growing SRB were spiked in duplicate 5-ASA (MW 153.1, Sigma) to make final 5-ASA concentrations 0, 20, 40, and 100 mmol/l faecal slurry. Sample incubations were carried out as described above and two unspiked 8 ml aliquots were immediately frozen in duplicate to provide blanks. For each subject, Jørgensen distillations of sulphide were carried out in two batches on the same day using one control sample with each run. The experiment was repeated in three of the seven subjects using aliquots of faecal slurry spiked in duplicate with sulphasalazine (MW 398.4, Sigma) instead of 5-ASA at a final concentration of 20 mmol/l faecal slurry.
DETERMINATION OF FAECAL SULPHATE
Frozen slurry samples (−25°C) were thawed for measurement of faecal sulphate concentrations as described by Florinet al.39 To measure free sulphate, duplicate samples were treated with propan-2-ol to precipitate protein and diluted in eluant buffer (final faeces dilution 1/200) prior to analysis by anion exchange chromatography. Isocratic separation of sulphate was obtained using an ion chromatograph (QIC, Dionex UK) with a 50 μl injection loop and an Ionpac AG9-SC guard column in series with an Ionpac AS9-SC analytical column. The apparatus was equipped with an anionic micromembrane suppressor (AMMS-II, Dionex), continuously regenerated by 12.5 mmol/l sulphuric acid, and a conductivity detector (Dionex). The eluent was a mixed solution of 1.8 mmol/l sodium carbonate and 1.7 mmol/l sodium bicarbonate in deionised (18.2 MΩ) water of high purity (Elga) at a flow rate of 1.8 ml/min. Chromatogram peaks were measured by a SpectraPhysics 4290 integrator calibrated with external standard sodium sulphate solutions. For measurement of total faecal sulphate, samples were hydrolysed by 1.6 M HCl at 90°C prior to dilution. Bound sulphate was calculated by subtraction of free from total sulphate.
STATISTICAL ANALYSIS
Results are expressed as median (interquartile range). Statistical analysis was performed using the statistical package Systat, version 5.2.1 (Mann-Whitney U test). A p value of less than 0.05 was considered significant.
Results
CULTIVATION AND GROWTH OF FAECAL SRB
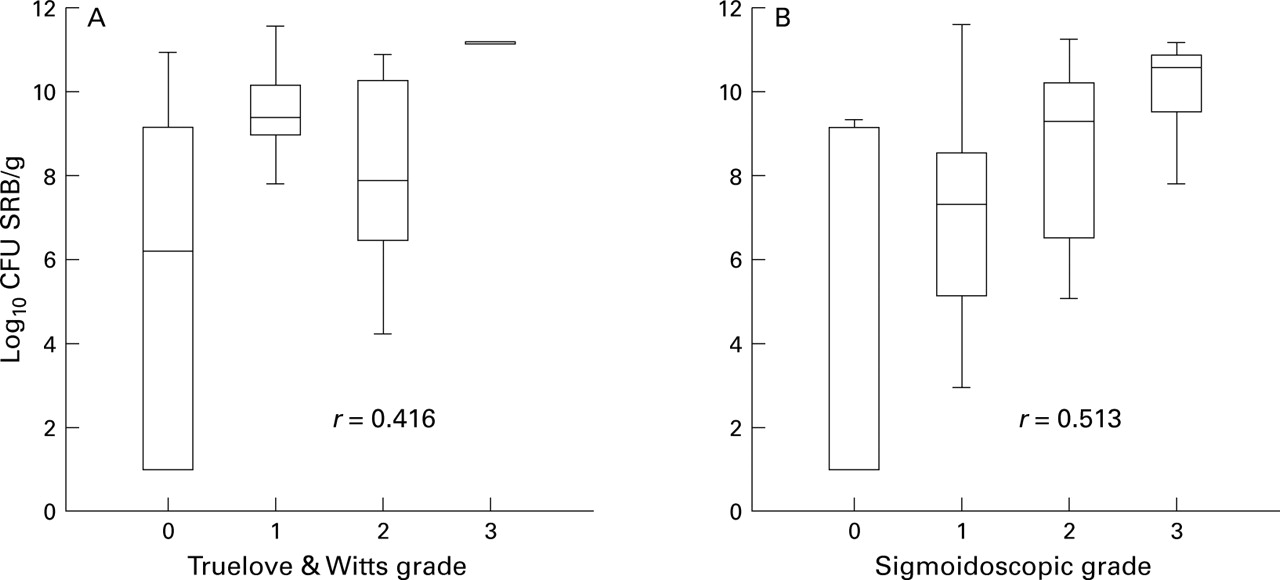
Growth of SRB was detectable in 23/23 patients with active ulcerative colitis, 10/14 patients with quiescent disease, and 16/16 controls. There was a significant relation between total viable counts of SRB and both the global clinical grade32 of patients with ulcerative colitis (r=0.42, Spearman rank correlation test; p<0.0001) and the sigmoidoscopic grade (r=0.51, p<0.01; fig 1). Although SRB counts were not significantly different between controls and all patients with ulcerative colitis studied, counts were significantly higher in patients with ulcerative colitis with clinically active disease compared with those in remission (p<0.005; table1).
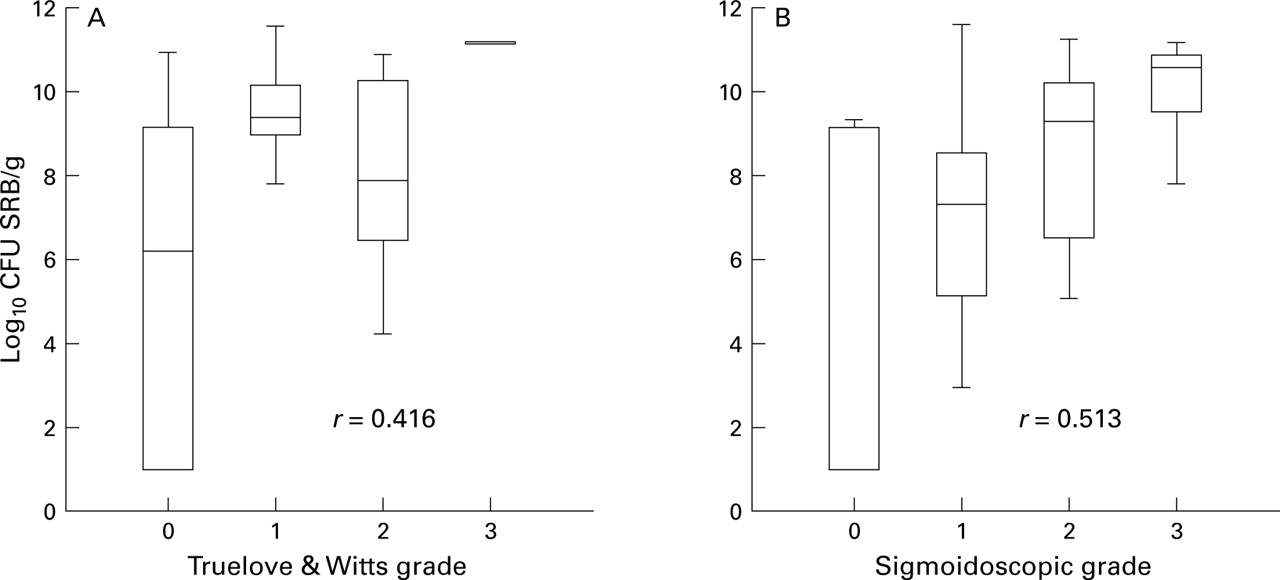
Box and whisker plots of total viable counts of sulphate reducing bacteria (SRB) related to (A) Truelove and Witts global clinical grade, and (B) sigmoidoscopic grade of mucosal inflammation in patients with ulcerative colitis (Spearman rank correlation test, p<0.0001).
Total viable counts of sulphate reducing bacteria (SRB) in faecal slurries from patients with ulcerative colitis (UC) and healthy controls
SRB could be differentiated into two phenotypic groups on the basis of colonial growth characteristics in agar shake tubes. Fast growing strains turned the agar shake medium uniformly densely black within 14 days at low dilutions, and individual colonies measuring 1–3 mm in diameter were readily identified at higher dilutions by the generation of ferrous sulphide. These SRB could be perpetuated by subculturing a representative colony into fresh Postgate's E liquid medium. By contrast, slow growing strains of SRB did not cause early blackening of the medium but could be identified as tiny black colonies, less than 1 mm in diameter, after 4–6 weeks of incubation. Perpetuation of these SRB strains was unsuccessful. Rapidly growing strains were present in 10/33 (30.3%) patients with ulcerative colitis and 7/16 (43.8%) healthy controls. SRB cultivated from faeces of four patients with ulcerative colitis who were studied sequentially possessed identical growth characteristics during active disease and remission.
Phase contrast microscopy of SRB colonies identified rapid growing strains as motile bacteria of curved morphology. Subsequent DNA extraction and 16S rRNA sequence analysis identified a representative strain as Desulfovibrio desulfuricans. Attempts to characterise slow growing strains in a similar manner were unsuccessful. Sensitivity testing of Desulfovibrio desulfuricans in pure culture with 100 mg/l sulphasalazine or 5-ASA showed no effect on bacterial growth.
SULPHATE REDUCING ACTIVITY
There was no overall difference in sulphate reduction rate between patients with ulcerative colitis and controls. Median SRA (nmol sulphate reduced/day/g wet weight faeces) in all subjects harbouring rapidly growing SRB of the desulfovibrio type was significantly higher than in those harbouring slow growing SRB strains (28.5 versus 0.5, p<0.0001; fig 2).

Box and whisker plot showing sulphate reduction rates in faeces of all subjects (patients and healthy controls combined) according to colonial growth characteristics of sulphate reducing bacteria (SRB). Rapidly growing SRB are desulfovibrio-like.
EFFECT OF 5-ASA DRUGS ON SULPHATE REDUCTION IN VITRO
5-ASA inhibited dissimilatory sulphate reduction in faecal slurries in a dose dependent manner in all seven selected healthy subjects harbouring rapidly growing SRB of the desulfovibrio type. The percentage reduction of the basal rate was calculated for each subject at four concentrations of 5-ASA in faeces (fig 3). The inhibition by 5-ASA of sulphate reducing activity in the faecal slurries was highly significant for 20 mM (p=0.0005), but higher concentrations effected no further significant inhibition (p=0.22). By extrapolation from the dose response curve, the IC50 (5-ASA concentration producing half maximal inhibition) for sulphate reduction in vitro is 18.75 mM. Sulphasalazine also inhibited dissimilatory sulphate reduction in faecal slurries from all three healthy volunteers tested. At a final concentration of 20 mM, sulphasalazine reduced the mean sulphate reduction rate to 4.4% of basal values (p=0.00001).

Dose response effect of 5-aminosalicylic acid (5-ASA) on dissimilatory sulphate reduction by sulphate reducing bacteria in faecal slurries. Values are mean (SE) for seven healthy subjects selected with high basal sulphate reducing activity.
FAECAL TOTAL SULPHIDE CONCENTRATION
There was no overall difference in faecal sulphide concentration between patients with ulcerative colitis and controls. In light of the effect of 5-ASA drugs on sulphate reduction in vitro, subgroup analysis of the data was performed to identify any effect of treatment with oral 5-ASA drugs on faecal sulphide concentration in vivo (table 2). Faecal sulphide in patients with ulcerative colitis not receiving 5-ASA drugs (n=8) was significantly higher than in healthy controls (n=16) (0.53 mM versus 0.25 mM, p=0.027; fig 4). 5-ASA treated patients (n=29) had a significantly lower median faecal sulphide concentration than patients with ulcerative colitis not receiving treatment with oral 5-ASA drugs (n=8) (0.18 mM versus 0.55 mM, p=0.003; fig 5A). There was no significant difference in median faecal sulphide between patients receiving (n=11) and those not receiving (n=26) prednisolone (0.23 mM versus 0.21 mM, p>0.05; fig 5B). Faecal sulphide concentrations were also analysed in patients with low sulphate reducing activity (n=24) to determine whether there was an effect of 5-ASA drugs on bacterial sulphide production by a mechanism other than dissimilatory sulphate reduction. 5-ASA treated patients (n=17) had a significantly lower median faecal sulphide than those not receiving 5-ASA drugs (n=7) (0.16 mM versus 0.45 mM, p=0.01) but no significant differences were detected with respect to treatment with prednisolone.
Total faecal sulphide concentration in patients with ulcerative colitis (UC) according to treatment group
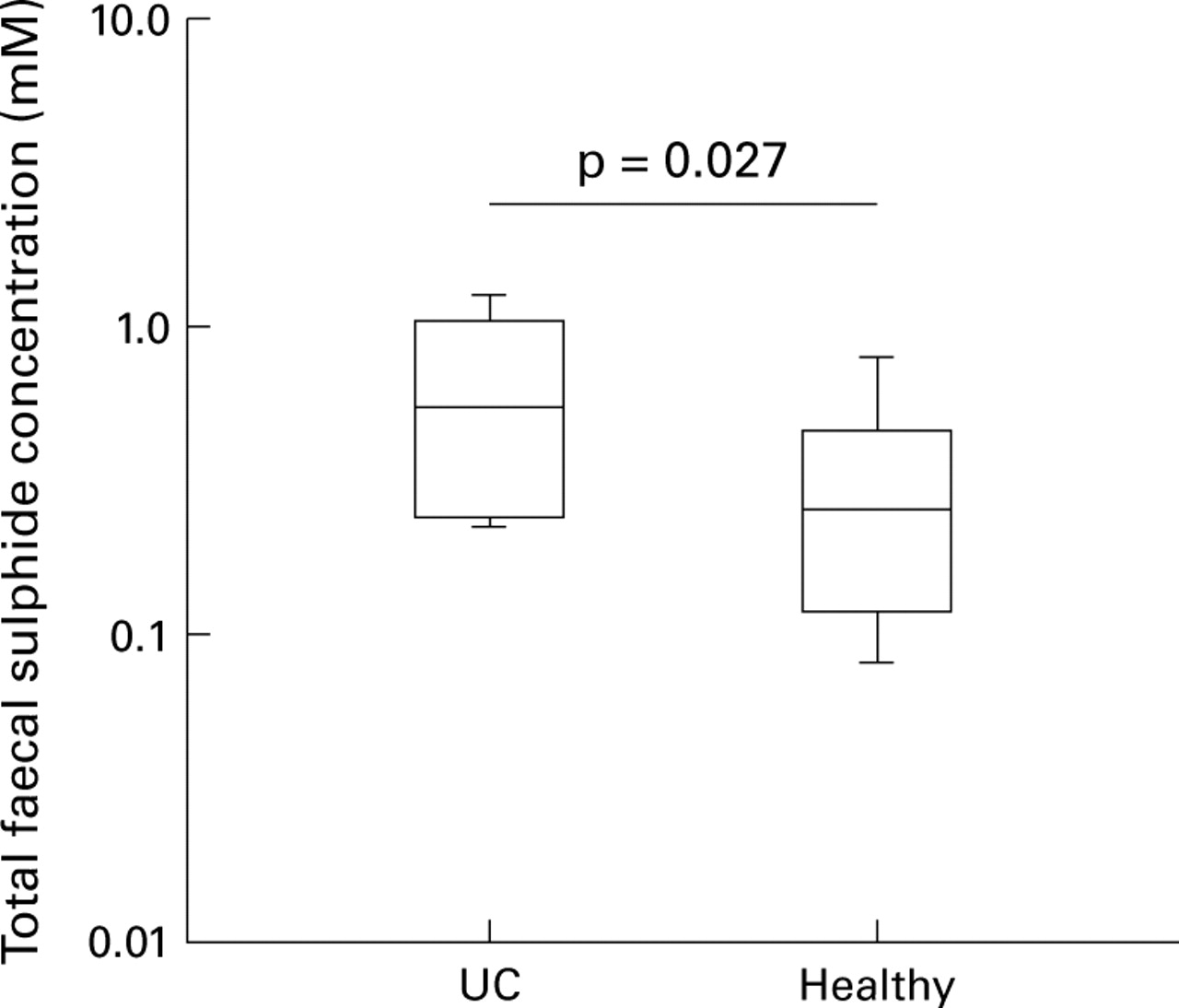
Box and whisker plot showing total faecal sulphide concentration in patients with ulcerative colitis (UC) (n=8) not receiving 5-aminosalicylic acid, and healthy controls (n=16).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Box and whisker plots showing total faecal sulphide concentration in patients with ulcerative colitis (n=37) grouped according to treatment with oral drug therapy. (A) Sulphide concentrations in patients treated with 5-aminosalicylic acid (5-ASA) drugs alone or in combination with prednisolone (n=29) versus no 5-ASA (n=8). (B) Sulphide concentrations in patients treated with prednisolone alone or in combination with 5-ASA (n=11) versus no prednisolone (n=26).
FAECAL SULPHATE CONCENTRATION
Table 3 shows free and total faecal sulphate concentrations. Levels were three orders of magnitude lower than sulphide concentrations. Free faecal sulphate was significantly higher in patients with ulcerative colitis and in those with extensive and left sided disease but there was no significant relation with disease activity. Free sulphate was significantly lower in faeces of all subjects with high sulphate reducing activity. Total faecal sulphate concentrations exhibited the same trends as for free concentrations, although the difference between ulcerative colitis and controls was not significant. The proportion of total faecal sulphate as bound sulphate (bound/total sulphate ratio) was lower in patients with ulcerative colitis, irrespective of disease activity, compared with controls (0.516 versus 0.718, p=0.04).
Free and total faecal sulphate concentration (nmol per g wet weight faeces) in patients with ulcerative colitis (UC) and healthy controls
BREATH METHANE STATUS
Breath methane was detectable (>1 ppm above background) in 7/34 (20.6%) patients with ulcerative colitis and 4/16 (25%) healthy controls. Median breath methane in patients with ulcerative colitis and controls was 9 and 24.5 ppm respectively. Eight of these 11 methanogenic subjects had low sulphate reducing activity in their faeces, as expected. However, the other three had rapidly growing SRB of desulfovibrio type in their faeces with concomitant high sulphate reducing activity.
Discussion
Hydrogen sulphide inhibits butyrate oxidation within the colonocyte and may be important in the pathogenesis of ulcerative colitis. This study has attempted to identify a relation between the growth and activity of sulphate reducing bacteria, effects of 5-ASA treatment, and hydrogen sulphide concentration in faecal slurries from healthy subjects and patients with ulcerative colitis.
Total viable counts of SRB were significantly higher by almost three log values in faeces of patients with active as opposed to quiescent ulcerative colitis and correlated with clinical and sigmoidoscopic grades of disease severity. In in vitro studies certain colitic strains of SRB have adapted to fast turnover times40 and in healthy people the viable count of SRB in faeces is inversely related to gut transit time.41 A non-specific effect of diarrhoea is therefore a possible factor in the enumeration of SRB and other bacteria from faeces in ulcerative colitis, a phenomenon previously observed with respect to coliforms by Gorbach et al.42 Counts and carriage rates of SRB from faeces of patients with ulcerative colitis were not significantly different from healthy controls, a finding at odds with the only other comparative study in the literature.40 Methodological differences and longer incubation times may explain some of these discrepancies. Furthermore, the high carriage rates of SRB among healthy subjects in this study have been confirmed elsewhere.43
An important finding in the present study is the demonstration of significant physiological differences among strains of SRB in the human colon. The growth characteristics of SRB strains can be divided into rapid and slow growing phenotypes which show high and low sulphate reducing activity respectively. 16S rRNA sequence comparison techniques44 have identified a rapidly growing SRB strain from a patient with ulcerative colitis asDesulfovibrio desulfuricans. Phenotypically similar SRB strains are of similar prevalence, approximately 30–40%, in both the colitic and healthy colon and so their presence, per se, cannot be linked directly to the pathogenesis of ulcerative colitis. Faeces containing rapid growing desulfovibrios had significantly higher sulphate reduction rates than faeces containing slow growing SRB strains, suggesting that the latter are functionally unimportant in terms of dissimilatory sulphate reduction to hydrogen sulphide in the colon, possibly due to a low capacity for hydrogen utilisation. These findings have important implications for the fate of molecular hydrogen produced during fermentation in the human colon. For example, 50% of all subjects in this study (18/34 ulcerative colitis, 7/16 controls) were breath methane negative and their faeces showed low sulphate reducing activity (SRA); these data are consistent with acetogenesis (or some unknown hydrogen utilising process) as a quantitatively important route for hydrogen disposal in the colon.45-47Furthermore, three methanogenic subjects harboured rapidly growing SRB with high SRA in their faeces which shows that sulphate reduction and methanogenesis are not mutually exclusive metabolic processes in terms of hydrogen utilisation in the colon.
In this study, sulphate concentration was significantly higher in faeces with low SRA than in faeces with high SRA. This difference may be due to the greater reduction of much of the colonic sulphate pool to sulphide by rapidly growing strains of SRB, with consequent lower excretion of sulphate in faeces. These strains are likely to be sulphate limited in the distal colon. Although faecal constituents are not entirely representative of events occurring in the colon itself, these data suggest that slow growing SRB are poor utilisers of sulphate during oxidative metabolism, in spite of an abundance of hydrogen production in faeces.48 This may explain the continuing controversy concerning competition between SRB and methanogenic bacteria for hydrogen.47 49 50 Measurement of SRA in individual faecal samples is thus essential and cannot be predicted from a knowledge of breath methane status alone. Furthermore, reliance on hydrogen sulphide generation, as has been made by some centres,50-52 may not be wholly indicative of the metabolic activities of SRB, as hydrogen sulphide is also released during sulphur amino acid fermentation.
Although faecal total sulphate concentrations in patients with ulcerative colitis were not significantly different to controls, free sulphate was significantly higher in the ulcerative colitis group, particularly in those with extensive disease (table 3). These observations were independent of the sulphate reducing activity of SRB in faeces. Differences in dietary intake of sulphate between subject groups are unlikely to be responsible. Assuming that endogenously produced mucin contributes significantly to the total colonic sulphate pool,39 its desulphation by gut bacteria, includingBacteroides spp.,53 will contribute to the colonic pool of free sulphate. Tsaiet al have recently shown increased mucin sulphatase activity in faecal filtrates from patients with ulcerative colitis.54 This is likely to influence the activities of SRB since they do not possess sulphatase enzymes and are thus unable to metabolise sulphated carbohydrates directly. Assuming that increased desulphation will decrease the proportion of total faecal sulphate in a bound form and release more free sulphate into the lumen in ulcerative colitis, the observations in this study are consistent with the data of Tsai et al. This supports the theory that increased bacterial desulphation of secreted colonic mucin is contributory to the observed reduction in mucus gel thickness55 and possible loss of barrier function in ulcerative colitis.
The in vitro study has shown for the first time that 5-ASA containing drugs inhibit dissimilatory sulphate reduction by rapidly growing SRB. A dose response effect was observed from which the IC50 of 5-ASA for sulphate reduction was extrapolated as 18.75 mM. This is comparable with published colonic luminal concentrations of 5-ASA in patients taking maintenance doses of 5-ASA drugs (for example, oral mesalazine 2 g per day).56-58 Sulphasalazine 20 mM appeared to have a more potent effect on dissimilatory sulphate reduction in vitro than similar concentrations of 5-ASA, causing 95.6% inhibition of basal sulphate reduction rates compared with control conditions. Previous sensitivity testing of D desulfuricans in pure culture in our laboratory with 100 mg/l sulphasalazine or 5-ASA showed no effect on bacterial growth,59 however, and it is therefore proposed that 5-ASA inhibits the ability of SRB to reduce inorganic sulphate to sulphide. Possible mechanisms include competition with sulphate transport into the bacterial cell or alternatively destruction of intracellular ATP via the ATP sulphurylase reaction. These actions have both been described for molybdate, the classic metabolic inhibitor of dissimilatory sulphate reduction.60 61 Although 5-ASA clearly has an important and pluripotential mode of action at the level of the colonic epithelial cell which is clinically beneficial,62 63 these data show that 5-ASA, at physiological concentrations in faeces, significantly inhibits luminal production of hydrogen sulphide by a hitherto unrecognised action on colonic bacteria.
Median faecal sulphide concentration was significantly lower in those patients receiving treatment with oral 5-ASA drugs, whereas there was no such effect following treatment with oral prednisolone. Roediger's group have been unable to confirm these findings,64possibly due to the smaller number of patients studied, the majority of whom were in clinical remission. Treatment with 5-ASA was also associated with significantly lower faecal sulphide concentrations in patients with ulcerative colitis harbouring only slow growing strains of SRB with low sulphate reducing activity. It therefore appears that 5-ASA drugs also inhibit the luminal production of sulphide by acting on a mechanism other than dissimilatory sulphate reduction. Hydrogen sulphide may be generated as a result of fermentative degradation of the sulphur containing amino acids, cysteine and methionine, by other gut bacteria.65-68 Roediger and Duncan68have employed HPLC analysis with fluorescence detection to determine sulphide concentrations in faecal slurries and reported 5-ASA mediated inhibition of sulphide production in samples spiked with sulphur amino acids. This observation supports our proposal that 5-ASA can inhibit bacterial sulphide production from both sulphur amino acid fermentation and sulphate reduction.
In this study, fresh faeces were obtained from eight patients with ulcerative colitis who were not receiving treatment with 5-ASA drugs. Total sulphide measured per gram wet weight faeces was significantly higher in these patients compared with healthy controls. These findings are at variance with the study of Moore and colleagues64who found no differences in luminal sulphide concentration between patients not taking 5-ASA drugs and controls. This may be due to the fact that four of the six patients with ulcerative colitis studied were in clinical remission, in contrast to our study where patients had predominantly active disease. Our data suggest that there is a greater net production of hydrogen sulphide from gut bacteria in ulcerative colitis compared with healthy individuals, a factor which may be underestimated here on account of generally more dilute faeces in the ulcerative colitis group. Levitt's group have recently reported similar findings.69 SRB are probably important in the luminal production of hydrogen sulphide in the 30% of patients with ulcerative colitis which harbour rapidly growing desulfovibrios in their colons. The in vitro generation of sulphide by SRB (mean of 28.5 nmol/day/g for rapidly growing SRB) is modest compared with total faecal sulphide (0.25–0.55 mM) which suggests that hydrogen sulphide production from proteolysis may be contributory with respect to disease pathogenesis, particularly since amino acid fermentation is quantitatively greater in the distal rather than the proximal colon.70 This may be relevant to the findings of a prospective epidemiological dietary study which showed that patients with ulcerative colitis consumed significantly more protein immediately prior to the onset of their inflammatory bowel disease than subjects in a case control population.71 Furthermore, the recent report of clinical benefit of a low protein diet in ulcerative colitis72 sheds new light on mechanisms of disease pathogenesis as well as future possibilities for novel dietary therapy.
Faecal sulphide concentrations represent a balance between the production and removal of this toxic metabolite. A number of detoxification mechanisms are believed to occur. In the lumen, sulphide can be assimilated into sulphur amino acids during growth of gut bacteria which may constitute an important, and hitherto unexplored, luminal pathway for detoxification.73 Hydrogen sulphide is readily absorbed by the colonic epithelium66 where it is oxidised to inorganic sulphate and subsequently excreted in the urine.74 75 Alternatively, hydrogen sulphide is methylated by mucosal S-thiolmethyltransferase (TMT) to less harmful products such as methane thiol and dimethyl sulphide.76 A net luminal excess of hydrogen sulphide shown in patients with ulcerative colitis in this study may therefore be due to increased bacterial production and/or defects in one or more of these mechanisms of detoxification. TMT activity in erythrocyte membranes is significantly higher in patients with ulcerative colitis compared with matched controls, possibly in response to excess hydrogen sulphide exposure.77 The overall effect of a net excess of luminal hydrogen sulphide would be inhibition of butyrate oxidation within the colonocyte followed by aberrant cellular metabolism, the destruction of mucosal barrier function, and the genesis or perpetuation of mucosal inflammation.
Acknowledgments
Dr Pitcher was supported by research grants from the National Association for Colitis and Crohn's disease, Addenbrooke's NHS Trust Endowment Fund, and Peel Medical Research Trust.
References
Footnotes
- Abbreviations used in this paper:
- 5-ASA
- 5-aminosalicylic acid
- ANOVA
- analysis of variance
- CFU
- colony forming units
- IQR
- interquartile range
- SCFA
- short chain fatty acid
- SRA
- sulphate reducing activity
- SRB
- sulphate reducing bacteria