Article Text

Abstract
Background: Coeliac disease is caused by an immune response to gluten. As gluten proteins are proline rich they are resistant to enzymatic digestion in the gastrointestinal tract, a property that probably contributes to the immunogenic nature of gluten.
Aims: This study determined the efficiency of gluten degradation by a post-proline cutting enzyme, Aspergillus niger prolyl endoprotease (AN-PEP), in a dynamic system that closely mimics the human gastrointestinal tract (TIM system).
Methods: Two experiments were performed. In the first, a slice of bread was processed in the TIM system with and without co-administration of AN-PEP. In the second, a standard fast food menu was used. Samples of the digesting meals were taken from the stomach, duodenum, jejunum and ileum compartments at time zero until 4 hours after the start of the experiment. In these samples the levels of immunogenic peptides from gliadins and glutenins were assessed by monoclonal antibody-based competition assays, Western blot analysis and proliferation T-cell assays.
Results: AN-PEP accelerated the degradation of gluten in the stomach compartment to such an extent that hardly any gluten reached the duodenum compartment.
Conclusion: AN-PEP is capable of accelerating the degradation of gluten in a gastrointestinal system that closely mimics in-vivo digestion. This implies that the co-administration of AN-PEP with a gluten-containing meal might eliminate gluten toxicity, thus offering patients the possibility of abandoning (occasionally) their strict gluten-free diet.
Statistics from Altmetric.com
Coeliac disease is a small intestinal disorder characterized by an abnormal immune response to gluten proteins. In patients with coeliac disease the ingestion of gluten evokes an immune response in the small intestine that eventually results in T-cell infiltration and flattening of the mucosa.1 Patients experience malabsorption, diarrhoea and failure to thrive, leading to fatigue, osteoporosis and neurological symptoms. Gluten proteins are the storage proteins of wheat and contain high percentages of proline (20%) and glutamine residues (38%). Because of their unusually high content of proline,2 gluten is poorly degraded by enzymes present in the gastrointestinal tract. After ingestion, partly degraded gluten proteins reach the small intestine. Such fragments are good substrates for the enzyme tissue transglutaminase, which can convert the amino acid glutamine in gluten into the negatively charged glutamic acid. These modified gluten fragments can bind with high affinity to the disease-associated HLA-DQ2 or HLA-DQ8 molecules and induce inflammatory T-cell responses.3–6
Gluten is composed of two different protein families, the gliadins and the glutenins. The gliadins can be further subdivided into α, β, γ, and ω-gliadins and the glutenins into low molecular weight (LMW) and high molecular weight (HMW) glutenins. T-cell stimulatory epitopes have been identified in all these proteins.7–15
For patients with coeliac disease the only cure is a lifelong strict gluten-free diet, which in practice excludes all foods that contain wheat, barley, rye and oats. Because of the widespread use of gluten and gluten-like proteins in food products, a gluten-free diet has a great impact on the lifestyle of coeliac disease patients. For this reason the search for new treatments, which are compatible with a normal social lifestyle, is of great importance. In this respect several lines of research have been proposed. Studies have been performed in which wheat varieties are screened for the level of T-cell stimulatory epitopes. Wheat varieties with a low toxicity might form the basis for future breeding programmes to generate wheat varieties suitable for the generation of food products that can be consumed by coeliac disease patients.16–18 Another option is the use of enzymes that degrade the proline-rich gluten molecules before they reach the small intestine. In this context prolyl oligopeptidases have been investigated. Such enzymes are not only effective in degrading gluten, the generation of smaller gluten fragments also improves the digestibility of gluten proteins by rendering them more accessible to brush border enzymes.19–25 To avoid T-cell recognition, however, gluten must be degraded before it reaches the small intestine, and the prolyl oligopeptidases investigated are not active under the conditions found in the stomach. These enzymes are thus not suitable for oral supplementation as an alternative treatment for coeliac disease.
We recently described Aspergillus niger prolyl endoprotease (AN-PEP).26 This enzyme was found to degrade gluten peptides and intact gluten proteins efficiently. Moreover, the pH optimum of the enzyme is compatible with that found in the stomach and the enzyme is resistant to degradation by pepsin. These results indicate that this enzyme might be suitable for oral supplementation to degrade gluten proteins in food before they reach the small intestine. To test this we determined the efficiency of gluten degradation under near in-vivo conditions. To predict the efficacy of enzymes and drugs for therapeutic use in the gastrointestinal tract of humans, a dynamic, multicompartimental in-vitro system was developed.27 Validation studies demonstrated that this system, called TIM (TNO gastro-intestinal model), allows a close simulation of in-vivo dynamic physiological processes that occur within the lumen of the stomach and small intestine of humans and reliably predicts in-vivo data.28–30 The system is fully computerized and is based on parameters obtained from data of healthy volunteers. The main parameters of digestion, such as pH, body temperature, peristaltic mixing and transit, salivary, gastric, biliary, and pancreatic secretions, as well as the absorption of small molecules (e.g. nutrients and drugs) and water are simulated. Gastrointestinal passage and successive conditions can be adjusted in order to mimic parameters in humans at different stages (infant, adult, and elderly), different food intakes, and physiological or pathological conditions (such as gastric hyperacidity or pancreatic failure).30 31
The results of the present study demonstrated that gluten degradation was strongly accelerated by the presence of AN-PEP in the stomach compartment. AN-PEP was capable of degrading all T-cell stimulatory epitopes of gluten tested to levels below the detection limit of the methods used. The co-administration of AN-PEP with a gluten-containing meal may thus be a feasible approach to detoxify gluten before it can do harm in the small intestine of coeliac disease patients. This may offer patients an alternative to the strict gluten-free diet and thereby improve their quality of life.
MATERIALS AND METHODS
Enzyme prolyl endoprotease from Aspergillus niger
AN-PEP was produced and purified by DSM Food Specialties (Delft, the Netherlands) according to established procedures.32 Activity of the protein is expressed in proline protease units (PPU). A PPU is defined as the quantity of enzyme that releases 1 μmol of p-nitroanilide per minute at 37°C in a citrate/disodium phosphate buffer pH 4.6 and at a substrate concentration of 0.37 mM Z-Gly-Pro-pNA (Bachem, Bubendorf, Switzerland) and represents 10 mg of pure protein. The reaction products were monitored spectrophotometrically at 405 nM.
TIM experiments
The TIM has been described in detail previously.27–31 This model has compartments for the stomach, duodenum, jejunum and ileum (fig 1). Each compartment has a flexible inner wall surrounded by water at 37°C. Changing water pressure squeezes the walls to simulate peristaltic mixing of the food with the “secreted” electrolytes and enzymes. The transport of the chyme is regulated by the peristaltic valves that connect the successive compartments. Using various sensors in the compartments, the pH values, temperature, volumes, and pressure, as well as the gastric emptying and small intestinal passage of the food are computer controlled according to pre-set curves. For the present experiments all parameters in TIM were adjusted to simulate the average physiological conditions in the gastrointestinal tract of young healthy adults after the intake of the type of meal as described below. During 2.5 hours the gastric content was gradually delivered into the small intestine via the “pyloric valve”. After 5 hours approximately 80% of the small-intestinal content was gradually delivered into the “large intestine” (sampling bottle) via the “ileo-caecal valve”.

Two experiments were performed. In the first experiment 70 g of white bread (containing 5 g gluten) and 110 ml drinking water was homogenized together with 110 ml artificial saliva in the absence or presence of AN-PEP (200 mg pure enzyme/g protein). After 40 seconds of homogenization, the mixture was added to the stomach compartment of the TIM system containing 10 ml simulated gastric juice and the experiment was started. In the second experiment a quarter of a commercial fast food menu consisting of a bread bun, a hamburger, ketchup, French fries, supplemented with additional bread (50 g in total), was homogenized with 110 ml soda and 110 ml artificial saliva in the presence or absence of AN-PEP (200 mg pure enzyme/g protein) and introduced into the TIM system.
Sampling and analysis
During the transit of the homogenized food products through the compartments of the TIM system, samples of 2 ml were taken at time points: 0, 15, 30, 45, 60, 90, 120, 150, 180 and 240 minutes from the stomach, duodenum, jejunum and ileum compartments. The samples were snap frozen in dry ice to stop enzymatic activity.
Before analysis, the samples were thawed and AN-PEP activity was stopped by increasing the pH to 11–12 using 1 M NaOH, followed by neutralization with 1 M HCl. Hereafter the samples were kept at 85°C for 10 minutes to inactivate any residual enzymatic activity. Identical volumes from each sample were centrifuged for 10 minutes at 14 000 rpm to separate the water-soluble and water-insoluble components. The water-insoluble fractions were solubilized in the same volume of 6× protein sample buffer (60% glycerol, 300 mM Tris, pH 6.8, 12 mM EDTA, pH 8.0, 12% sodium dodecylsulphate, 864 mM 2-mercaptoethanol, 0.05% bromophenol blue). From these solutions 2 μl from each sample was used for the protein and Western blot analysis.
Synthetic peptides
Peptides were synthesized as described previously.33
Monoclonal antibodies against the Glia-α20, Glia-α9, Glia-γ1, LMW and HMW glutenin T-cell stimulatory epitopes
The specificity and the IgG subclass of the antibodies used in this study are presented in Table 1.
For the generation of a monoclonal antibody (mAb) specific for the α-gliadin-derived T-cell stimulatory epitope Glia-α20, mice were immunized with peptides chemically cross-linked to tetanus toxoid (TTd-DDDXPFRPQQPYPQP-amide). The fusion and screening of the hybridomas was performed as described.33 The minimal epitope of the anti-Glia-α20 mAb (FRPQQPYP) was determined using a set of partly overlapping 17-mer synthetic peptides.
Competition assays for the quantitative detection of T-cell stimulatory epitopes
Competition assays were performed as described earlier.16 33 Microtitre plates (Nunc Maxisorb, Immunoplate; Nunc, Copenhagen, Denmark) were incubated overnight with 2–5 μg/ml mAb in 0.1 M sodium carbonate/bicarbonate buffer, pH 9.2, at room temperature. Plates were washed in phosphate-buffered saline (PBS)/0.02% Tween-20 and residual binding sites were blocked with PBS/1% skim milk powder (Fluka, Zwijndrecht, the Netherlands). Of the gluten-containing samples, different dilutions were made in a buffer containing 50 mM Na2HPO4/NaH2PO4 pH 7.0, 150 mM NaCl, 0.1% Tween-20/0.1% skim milk and a protease inhibitor cocktail (Complete, Roche Diagnostics GmbH, Penzberg, Germany). For the detection of gliadins these were mixed with either a biotinylated Glia-α9 or Glia-γ1 gliadin T-cell epitope encoding peptides33 (table 2). For the detection of the Glia-α20 T-cell stimulatory epitope a biotinylated peptide encoding the Glia-α20 epitope was used as the indicator peptide (Bio-XKAKAKAKAXPFRPQQPYPQP-amide). For quantification, a standard curve was made using the prolamine working group gliadin reference material34 in the concentration range 10 μg/ml–10 ng/ml mixed with biotinylated indicator peptides. The assays, specific for the detection of T-cell stimulatory epitopes of LMW-glutenin, were calibrated using a 25-mer synthetic peptide as standard that contains the Glt-156 epitope.16 The HMW-glutenin-specific assay was calibrated using a chymotrypsin digest of six purified HMW-glutenin proteins (kindly provided by P. Shewry, Rothamsted Research, Harpenden, UK). Both standards were used in a concentration range from 1 μg/ml to 2 ng/ml.
The mixtures were incubated on plates for 1.5 hours at room temperature. Next, plates were washed and incubated for 30 minutes with streptavidin-conjugated horseradish peroxidase in PBS/0.1% skim milk.
Thereafter, bound peroxidase was visualised by incubation with a solution of 3′, 3′, 5, 5′-tetramethylbenzidine (Sigma-Aldrich, Zwijndrecht, the Netherlands). Finally, absorbance at 450 nm was read on a Multiscan plate reader (Wallac, Turku, Finland).
Protein analysis by 1D SDS–PAGE and Western blotting
To determine the level of T-cell stimulatory epitopes present in the water-insoluble fractions, these were solubilized in 6× protein sample buffer (60% glycerol, 300 mM Tris (pH 6.8), 12 mM EDTA pH 8.0, 12% SDS, 864 mM 2-mercaptoethanol, 0.05% bromophenol blue). The samples were separated on a 12.5% sodium dodecylsulphate–polyacrylamide gel electrophoresis (SDS–PAGE) gels for the detection of gliadins and LMW-glutenins and on 10% SDS–PAGE gels for detection of HMW-glutenins. The proteins were visualized either directly using Imperial Protein Stain (Pierce, Rockford, Illinois, USA), or after transfer to polyvinylidene fluoride (PVDF) membranes with the mAb specific for stimulatory T-cell epitopes from α and γ-gliadin33 (and this study) and HMW and LMW-glutenins.16
T-cell proliferation assay
For the T-cell proliferation assays the samples were treated with amylglucosidase and trypsin. Next the samples were incubated overnight at 37°C with guinea pig tissue transglutaminase (200 μg/ml) in the presence of 10 mM CaCl2. Proliferation assays were performed in triplicate in 150 μl Iscove’s Dulbecco’s medium (BioWhittaker, Vervier, Belgium) supplemented with 10% human serum in 96-well flat bottom plates (Costar, Corning Inc., Corning, New York, USA) using 104 gluten-specific T cells stimulated with 105 irradiated HLA-DQ2-matched allogenic peripheral blood mononuclear cells (3000 RAD). Of the samples three different amounts of the samples were tested: 0.5 μl, 0.16 μl and 0.05 μl, respectively. These amounts were shown to be non-toxic for the T cells. After 48 hours incubation at 37°C, cultures were pulsed with 0.5 μCi of 3H-thymidine, harvested 18 hours later, and the thymidine incorporation was quantified with a liquid scintillation counter (1205 Betaplate Liquid Scintillation Counter; LKB Instruments, Gaithersburg, Maryland, USA).
RESULTS
Digestion of bread and a fast food menu in the TIM system
To study the effect of AN-PEP on the degradation of gluten proteins in the gastrointestinal tract, two experiments were performed. These experiments were designed to represent a small meal (e.g. breakfast) and a more complex meal (e.g. dinner). In the first experiment 70 g of white bread was homogenized together with artificial saliva in the presence or absence of AN-PEP. In the second experiment a quarter of a fast food menu consisting of a sandwich, a hamburger, ketchup, French fries and a glass of soda was homogenized with artificial saliva in the presence or absence of AN-PEP. As the quantity of bread in this menu was relatively low (16 g), extra bread was added to a total amount of 50 g. After homogenization the mixtures were introduced into the TIM system, mimicking the human gastrointestinal tract (fig 1). Samples were collected at time points 0, 15, 30, 45, 60, 90, 120, 150, 180 and 240 minutes after the start of the experiment from the stomach, duodenum, jejunum and ileum compartments.
In order to study the degradation of gluten in the TIM experiments, three types of assays were performed. The presence of gluten peptides in the water-soluble fractions was measured by mAb-based competition assays specific for T-cell stimulatory peptides present in gliadin (Glia-α9, Glia-α20, Glia-γ1) and LMW and HMW-glutenin16 33 (this study). The presence of T-cell stimulatory epitopes from gluten in the water-insoluble fractions was determined by Western blot analysis. Moreover, samples of the fast food menu taken at 60 minutes after the onset of the experiment were tested by T-cell proliferation assays.
AN-PEP accelerates the degradation of gluten present in white bread
After the introduction of the white bread to the TIM system, the levels of all gluten peptides tested steadily decreased in the water-soluble compartment in the absence of AN-PEP. Even after 120 minutes, however, significant levels of gluten peptides were still detectable in the stomach compartment (fig 2). In contrast, when the white bread was introduced together with AN-PEP the digestion of gliadins (fig 2A and B) and glutenins (fig 2C and D) was accelerated at all time points.

Similarly, Western blot analysis indicated that the degradation of gluten caused by the addition of AN-PEP was accelerated. In the water-insoluble fractions of the stomach compartment α-gliadin, γ-gliadin and HMW-glutenin proteins were detectable (fig 3). Although a significant degradation of gluten was seen in the absence of AN-PEP, both gliadins (fig 3A and B) and glutenins (fig 3C) were still present after 120 minutes in the stomach compartment. In the presence of AN-PEP, however, faster degradation of both the gliadins (fig 3A and B) and the glutenins (fig 3C) occurred. This difference in degradation is already apparent after 45 minutes, and after 90 minutes gluten proteins could no longer be detected in the AN-PEP-treated fractions (fig 3).

The duodenum fractions contained only a small amount of water-insoluble material. Western blot analysis failed to detect intact gluten proteins in these fractions, neither in the absence nor the presence of AN-PEP (results not shown).
AN-PEP accelerates the degradation of gluten present in a complex meal
In the second experiment the digestion of gluten in a fast food menu was monitored. In the absence of AN-PEP, gluten peptides were found to be degraded in the water-soluble fraction. Residual gluten peptides were, however, still detectable in the fractions collected from all compartments up to 150 minutes after the start of the experiment (fig 4). In the presence of AN-PEP, the degradation of gliadins (fig 4B) and glutenins (fig 4D) was accelerated. In the absence of AN-PEP, gluten peptides could be detected in the samples that were collected from the duodenum, jejunum and ileum compartments (fig 4A and C). In the presence of AN-PEP, much lower amounts of gluten peptides could be detected in these samples (fig 4B and D).
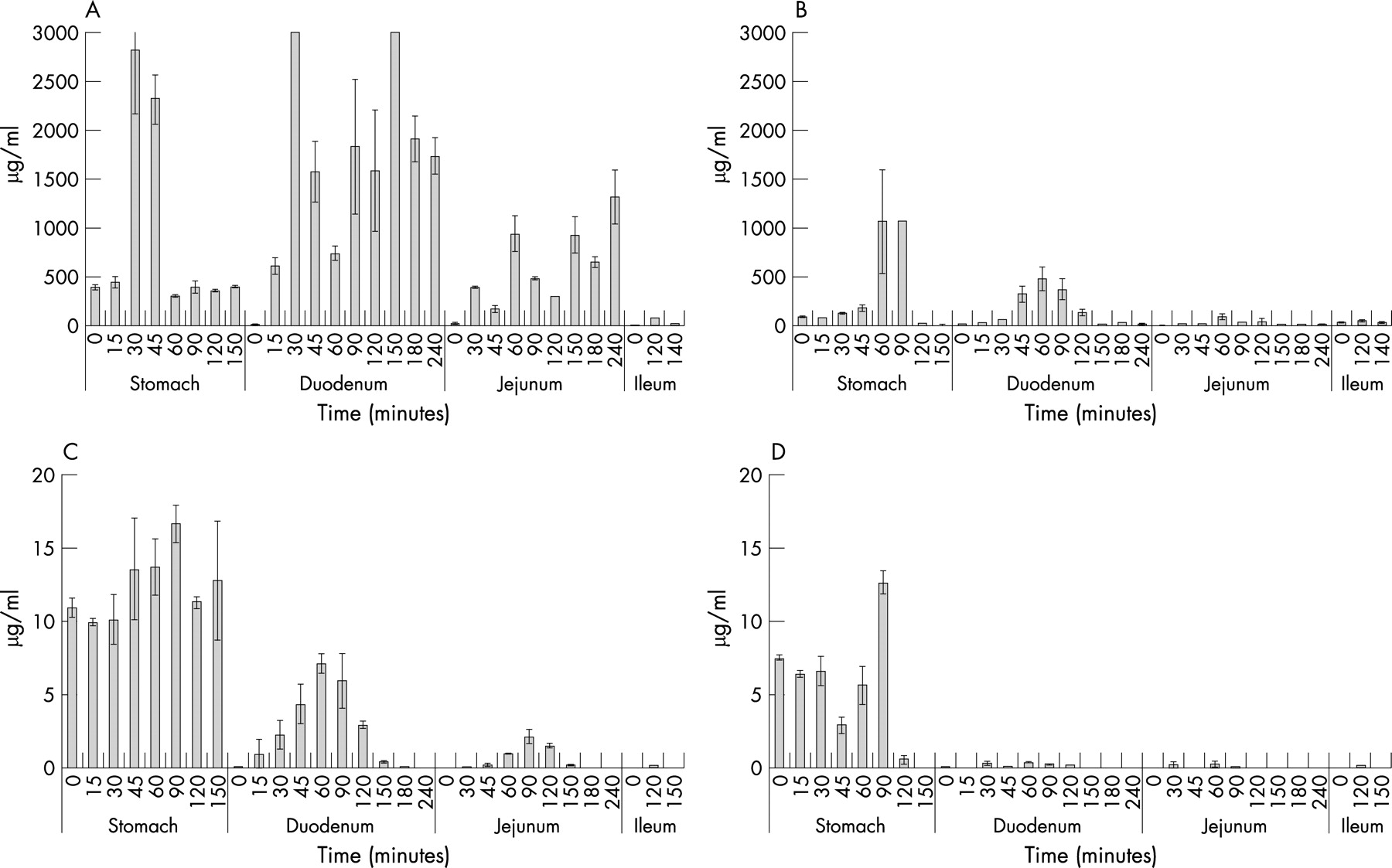
Again, the analysis of the water-insoluble material collected from the stomach and duodenum compartment indicated accelerated degradation of the gluten in the meal. In the presence of AN-PEP the amount of α-gliadin is clearly decreased in the stomach compared with digestion in the absence of AN-PEP (fig 5A). Directly after homogenization (t = 0 minutes) a clear difference is apparent and after 60 minutes no more gluten proteins could be detected in the AN-PEP samples, although such proteins were clearly still present in the control samples. Similarly, the HMW-glutenins were degraded faster in the presence of AN-PEP (fig 5C). After 60 minutes no more HMW-glutenins could be detected in the AN-PEP fractions, whereas partly degraded HMW-glutenins were still present in the control fractions (fig 5C). Finally, the degradation of the γ-gliadins and LMW-glutenins was similarly enhanced by the addition of AN-PEP (fig 5B and D).
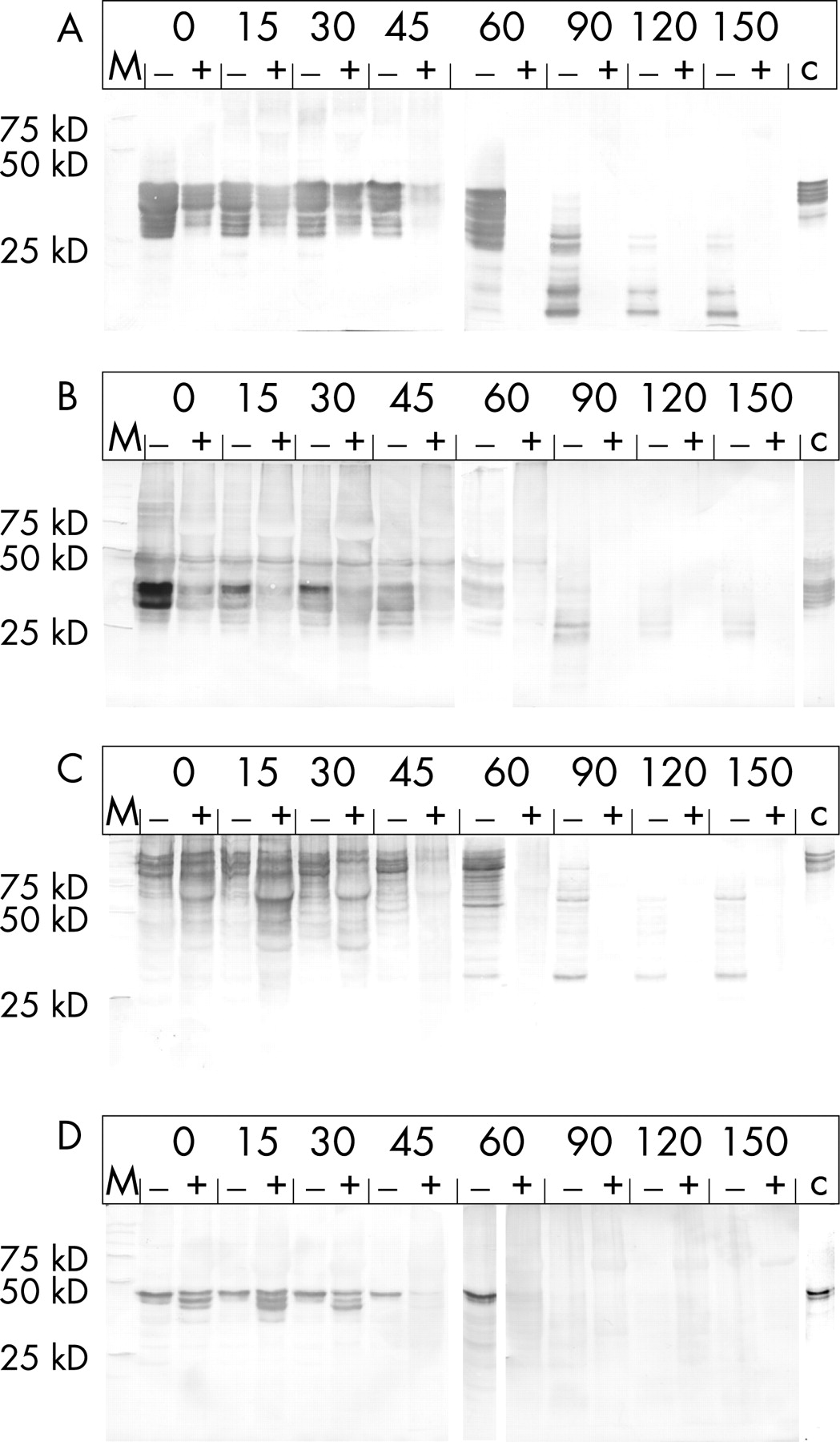
Similar to the white bread experiment, the amount of water-insoluble material in the duodenal fractions was too small to allow the detection of gluten proteins (results not shown).
Small amounts of gluten peptides could still be detected in the AN-PEP-treated samples that were measured with the antibody-based competition assays. As these assays detect gluten fragments that are smaller than those required for T-cell stimulation, we determined whether AN-PEP destroyed the T-cell stimulatory properties in the TIM system. T-cell clones specific for the Glia-α9, Glia-α20, LMW-glutenin and the HLA-DQ8-restricted HMW-glutenin-derived T-cell epitopes were incubated with the untreated and the AN-PEP-treated samples and T-cell proliferation was measured (fig 6). Whereas in the untreated samples T-cell stimulatory activity was clearly present, AN-PEP treatment completely abolished this activity.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
It is well established that coeliac disease patients are intolerant to gluten and that a strict gluten-free diet is an effective treatment for coeliac disease. The lifelong maintenance of such a diet, however, is hard to achieve. First of all, the western diet is heavily based on gluten-containing foods such as bread and pasta. In addition, because of its special properties, like its elasticity and capacity to bind water, gluten is often added to foods that would otherwise be gluten free. Finally, many foods that are rendered gluten free may still contain traces of gluten, while naturally gluten-free ingredients are often contaminated with gluten. Coeliac disease patients can, therefore, inadvertently be exposed to gluten. Moreover, a gluten-free diet can cause social constraints and this can lead to non-compliance with the diet. For these reasons, an alternative to a gluten-free diet would be useful to patients. Oral supplementation with enzymes to degrade gluten before it causes damage has been suggested for this purpose. Initially this proposition was based on the theory that coeliac disease is caused by an enzyme deficiency,35 and the use of non-human proteases for gluten detoxification had already been proposed in the late 1950s.36 A clinical trial was recently performed in which an extract from animal intestines was used. It was shown that the enzyme therapy offered better protection than placebo. The differences between enzyme therapy and placebo were, however, small and no complete protection was given by the extract. Furthermore, not all patients benefitted to the same extent from enzyme supplementation.37
As gluten is rich in the amino acid proline, other recent studies have used post-proline cutting enzymes. Promising enzymes tested are the prolyl oligopeptidases from Flavobacterium meningosepticum, Sphingomonas capsulate, and Myxococcus xanthus. These enzymes are capable of degrading proline-containing peptides that are otherwise resistant to degradation by proteases in the gastrointestinal tract in vitro.23 38 39 The suitability of these enzymes to degrade gluten in vivo is, however, questionable because the enzymes have a pH optimum between 7 and 8, and do not function at the acid pH of the stomach. Moreover, they are efficiently broken down by pepsin.23 Finally, as a result of their structure, in which a β-propeller domain restricts entry into the active site of the enzymes, they preferentially cleave short peptides.26 Encapsulation of these prolyl oligopeptidases was proposed to protect them against gastric juice.39 In a recent ex-vivo study, using biopsy-derived intestinal tissue mounted in Ussing chambers, it was observed that only high dosages of prolyl oligopeptidase tested were capable of eliminating the accumulation of immunogenic peptides in the serosal compartment.21 This indicates that, even if the enzyme is encapsulated, because of its relatively low efficiency, the prolyl oligopeptidase will not be able to degrade gluten before it reaches the proximal part of the duodenum, the site where gluten triggers inflammatory T-cell responses. To overcome these problems a combination therapy has been proposed. For this the prolyl oligopeptidase is combined with a cysteine endoprotease from barley, whose natural function is to degrade the gluten-like molecules in barley, to the benefit of the germinating seed. Whereas such a combination therapy may prove effective, a therapy based on one enzyme would have obvious advantages.40
We have recently investigated a prolyl endoprotease of Aspergillus niger, AN-PEP.32 We observed that AN-PEP has clear advantages over prolyl oligopeptidases as it is much more efficient in degrading gluten peptides, is active at low pH, and is resistant to pepsin degradation.32 41 Moreover, as AN-PEP is a prolyl endoprotease, it degrades gluten peptides as well as intact gluten proteins.32 41
These results indicated that AN-PEP may be suitable as an oral supplement for gluten degradation, but its efficacy in vivo remained to be established. Although this can be tested in animal models, as has been carried out for a prolyl oligopeptidase, the relevance of this for the human situation is debatable as the gastrointestinal tract of animals only partly simulates that of humans. We have therefore chosen to study the efficacy of AN-PEP using a dynamic gastrointestinal model that closely mimics the in-vivo conditions found in the human stomach and small intestine.27–29 Moreover, we have tested the capacity of AN-PEP to degrade gluten when present in a relatively simple food matrix, e.g. white bread, as well as when gluten is present in a complex food matrix, e.g. a complete fast food meal. The results of this study show that AN-PEP is indeed capable of degrading gluten under conditions found in the human gastrointestinal tract. We have carried out three types of analysis: competition assays that measure the presence of small gluten fragments in water-soluble fractions, Western blot to measure intact gluten proteins and relatively large fragments thereof (>5 kDa), and T-cell proliferation experiments that measure the presence of gluten-derived T-cell stimulatory peptides. Both the competition assays and the Western blot analysis gave comparable results: the disappearance of gluten proteins as indicated by Western blot analysis also led to a disappearance of smaller gluten peptides, as measured in the competition assay. Moreover T-cell proliferation assays on samples collected at 60 minutes from the digesting fast food menu showed that after the addition of AN-PEP, the stomach content is no longer capable of inducing T-cell proliferation. Taken together, these results indicate that AN-PEP completely degrades gluten into harmless fragments within 2 hours, the average passage time of food in the stomach. Besides, during the stay of the gluten in the stomach, the level of T-cell stimulatory epitopes of α and γ-gliadin HMW-glutenin and LMW-glutenin epitopes in the water-soluble fragments were significantly lower in the presence of AN-PEP than in its absence. This results in much lower amounts of T-cell stimulatory epitopes that are released from the stomach into the duodenum, thereby reducing the exposure of the proximal duodenum to T-cell-activating peptides. Moreover, in a previous study we have already demonstrated that AN-PEP will efficiently cut the alpha-gliadin peptide 31–43 and thus destroy the innate stimulatory properties of gluten.
It is important to note that our results may underestimate the rate of gluten degradation in the human body. Although the TIM system simulates the lumen of the human gastrointestinal tract, including peristaltic movements, secretion of juices from the salivary gland, the stomach, pancreas and liver, no brush border enzymes are present in the model. As brush border enzymes have an additive effect on proteolysis, which is also shown for the breakdown of gluten proteins,25 it is likely that the degradation of gluten is even more efficient than indicated by our results.
CONCLUSION
We tested the effect of the addition of AN-PEP on gluten degradation in a system that closely mimics the conditions in the upper human gastrointestinal tract. Our results demonstrate that within the timespan that food is normally present in the stomach, the co-administration of AN-PEP led to a complete disappearance of T-cell stimulatory peptides of gliadins and glutenins. Importantly, our results demonstrate that AN-PEP is capable of degrading gluten when this is present in a complex food matrix. Moreover, AN-PEP is derived from the food grade microorganism Aspergillus niger and is available on an industrial scale. Ultimately, clinical trials will be required to determine whether oral enzyme supplementation can remove all gluten toxicity. Our results indicate that AN-PEP is a very suitable candidate for testing in such trials.
REFERENCES
Footnotes
Funding: This study was supported by the Celiac Disease Consortium, an Innovative Cluster approved by the Netherlands Genomics Initiative and partly funded by the Dutch Government (BSIK03009)
Competing interests: Declared (the declaration can be viewed on the Gut website at http://www.gutjnl.com/content/vol57/issue1).
Linked Articles
- Digest