Article Text

Abstract
BACKGROUND cagpathogenicity island (PAI) is reported to be a major virulence factor of Helicobacter pylori.
AIM To characterisecagA and the cagPAI in Japanese H pylori strains.
METHODS H pylori isolates from Japanese patients were evaluated for CagA by immunoblot, for cagA transcription by northern blot, and for cagA and 13 othercag PAI genes by Southern blot.cagA negative strains from Western countries were also studied. Induction of interleukin-8 secretion from gastric epithelial cells was also investigated.
RESULTS All Japanese strains retained cagA. Fifty nine of 63 (94%) strains had all the cag PAI genes. In the remaining four, cag PAI was partially deleted, lacking cagA transcripts and not producing CagA protein. Details of the PAI of these strains were checked; three lacked cagB tocagQ (cagI) and continuously cagS tocag13 (cagII), and the remaining one lacked cagB tocag8. WesterncagA negative strains completely lackedcag PAI includingcagA. Nucleotide sequence analysis in one strain in which the cag PAI was partially deleted showed that the partial deletion contained 25 kb ofcag PAI and thecagA promoter. Interleukin-8 induction was lower with the cag PAI partial deletion strains than with the intact ones. All Japanesecag PAI deleted strains were derived from patients with non-ulcer dyspepsia, whereas 41 of 59 (70%) CagA-producing strains were from patients with peptic ulcers or gastric cancer (p<0.05).
CONCLUSIONS Most Japanese H pylori strains had the intactcag PAI. However, some lacked most of thecag PAI in spite of the presence ofcagA. Thus the presence of thecagA gene is not an invariable marker ofcag PAI related virulence in Japanese strains.
- Helicobacter pylori
- pathogenicity island
- Japanese
Abbreviations
- IL
- interleukin
- IS605
- insertion sequence 605
- PAI
- pathogenicity island
- SDS-PAGE
- sodium dodecyl sulphate-polyacrylamide gel electrophoresis, SSC, standard saline citrate
Statistics from Altmetric.com
Helicobacter pylori infection is almost always associated with gastric mucosal inflammation, but more advanced diseases such as peptic ulcers and gastric cancer occur only in a subset of infected patients.1-6 Although the pathogenesis of H pylori infection is not well understood, several factors have been proposed as possible virulence determinants.
The CagA surface protein is one of the most investigated putative virulence factors, encoded by the cagAgene.7 ,8 The gene is found in about 50–70% ofH pylori isolates in Western countries, and the production of CagA protein is reported to be associated with advanced gastroduodenal diseases.9-16 CagA positiveH pylori strains are known to induce interleukin-8 (IL-8) secretion from gastric epithelial cells.17-19 However, recent studies have indicated that it is not CagA itself but the products of other genes in the cag pathogenicity island (PAI) that are responsible for IL-8 induction.20-23The cag PAI is an approximately 40 kb region in the H pylori genome, possibly of extraneous origin.20 ,21 ,24 It is divided into two parts: the upstream cagII region containing at least 14 genes, and the downstream cagI region containing 16 genes. The cagA gene is located in the most downstream portion ofcagI. In some strains, the two parts of thecag PAI are reported to be interposed by a segment called insertion sequence 605 (IS605).21
Previous reports indicated that most cagApositive strains, most of which are capable of producing CagA protein,7 ,8 retain all the other genes of thecag PAI, whereas mostcagA negative strains completely lack thecag PAI.21 This implies that the presence of the cagA gene may be used as a marker of enhanced virulence associated with thecag PAI. However, Censiniet al 21 reported partial deletion of the cag PAI. Thus the status of the cagA gene and the othercag PAI genes should be individually investigated.
By using recombinant CagA and antibodies raised against it, we have previously shown that the prevalence of CagA-non-producingH pylori strains is relatively low in Japan (about 10%).25 ,26 However, these strains had not been characterised, and we conducted the present study to elucidate the relation between cagA and the othercag PAI genes in Japanese isolates.
Materials and methods
PATIENTS
H pylori strains were isolated from gastric biopsy specimens of 63 H pyloriinfected patients who had undergone endoscopy in Tokyo University Hospital. The patient population consisted of 40 men and 23 women with a mean age of 52.5 (range 22–78). The endoscopic findings were as follows: gastric ulcer (13); duodenal ulcer (12); non-ulcer dyspepsia (22); gastric cancer (16).
H PYLORI CULTURE
Biopsy specimens were cultured on Columbia agar with 5% (v/v) horse blood and Dent antibiotic supplement (Oxoid, Basingstoke, Hants, UK) at 37°C for five days under microaerobic conditions (Campy-Pak Systems; BBL, Cockeysville, Maryland, USA). The organisms were identified as H pylori by Gram stain morphology, colony morphology, and positive urease, catalase, and oxidase activities. The isolates were kept at −80°C in Brucella broth with 5% (v/v) fetal bovine serum (FBS) containing 16% (v/v) glycerol.
DETECTION OF CagA PROTEINS IN H PYLORI ISOLATES
Western blot analysis was performed to detect CagA proteins produced by H pylori strains as previously described.25 CagA protein was investigated usingH pylori whole cell lysates. The whole cell lysates were washed twice in normal saline, centrifuged, and boiled at 100°C for five minutes in Laemmli sample buffer27 before use. The whole cell lysates were separated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) (10% gel) and transferred to a nitrocellulose membrane (Schleicher & Schuell, Dassel Germany). The filter was blocked with buffer containing 5% (v/v) skimmed milk in 25 mM Tris/HCl, pH 7.4, containing 150 mM NaCl and 5 mM KCl. The membrane was incubated with a 1:100 dilution of rabbit antiserum against the recombinant CagA. After being washed, the membrane was incubated with 125I-labelled goat anti-rabbit IgG, and then exposed to x ray film.
PREPARATION OF GENOMIC DNA
H pylori bacterial cells, suspended in 10 mM Tris/HCl, pH 7.5, containing 1 mM EDTA, were treated with 0.3% SDS and 0.6 mg/ml proteinase K at 60°C overnight. DNA was extracted with phenol, phenol/chloroform and chloroform, precipitated with ethanol, and suspended in TE. The DNA was stored at −20°C until use.
CLONING OF THE GENES OF THE CAG REGION
DNA was extracted from H pylori ATCC 43579 cells. Oligonucleotide primers were designed to amplify the genes of the cag region;cagA, cagB,cagC, cagE (picB), cagM,cagN, cagQ,cagT, cag15,cag13, cag12,cag10, cag6–7, IS605, and the left end of the cag region (LEC) which contained both inside and outside genes of the cag PAI according to the nucleotide sequences of the cag region previously reported (GenBank accession numbers AC000108 and U60176) (table 1, fig 1). PCR cycling conditions were 30 cycles of 94°C for 30 seconds, 50°C for 30 seconds, and 72°C for one to two minutes. The PCR products were cloned into pCRII using the original TA cloning kit (Invitrogen, San Diego, California, USA). The nucleotide sequence of the insert was determined by the dideoxy chain termination procedure.28 Cloned plasmids were digested withEcoRI and BstXI, which were able to digest the full length of the PCR products, and the digests electrophoresed on 1% agarose gel. The appropriate fragments were then excised from the gels and the DNA extracted with the gene clean kit to be used as probes (Bio 101, La Jolla, California, USA).
Oligonucleotide primers used in this study

Map of the cag pathogenicity island. The names of the genes are from GenBank accession number AC000108 and U60176. Cross bars indicate probes used in this study. Fifty nine of 63 strains isolated in Japan all had cagI and cagII. Large deletions were revealed in the remaining four. cagA negative strains isolated in Western countries lacked whole cagI and cagII.
SOUTHERN BLOT ANALYSIS
For each sample, 5 μg of the genomic DNA was digested withHindIII for 12 hours at 37°C, electrophoresed on 1% agarose gel, and transferred to a nylon membrane (Amersham International, Bucks, UK) as described by Sambrooket al.29 The DNA samples were hybridised to previously prepared DNA probes, which were digested from cloned plasmids and labelled with [32P]dCTP with the Ready To Go DNA labelling kit (Pharmacia Biotech, Uppsala, Sweden). The membrane was prehybridised with ExpressHyb solution (Clontech, Palo Alto, California, USA) at 60°C for one hour, and hybridisation was carried out at 60°C for two hours. The membrane was washed four times with 2 × standard saline citrate (SSC)/0.05% SDS for 10 minutes at room temperature, twice with 0.1 × SSC/0.1% SDS for 20 minutes at 50°C, and then autoradiography was performed using a BAS2000 image analyser (Fuji Photo Film Co, Ltd, Tokyo, Japan).
RNA EXTRACTION AND NORTHERN BLOT ANALYSIS OFcagA TRANSCRIPTS
H pylori bacterial cells were washed with normal saline and suspended in TE. Total RNA was extracted using Rneasy Total RNA kit (Qiagen, Chatsworth, California, USA). For each sample, 10 μg total RNA was electrophoresed on formamide-agarose gel and transferred to a nylon membrane (Amersham International). The hybridisation was performed using the cagAgene as described above, except that the membrane was kept at 68°C instead of 60°C, and washed four times with 2 × SSC/0.05% SDS for 10 minutes at room temperature, twice with 0.1 × SSC/0.1% SDS for 20 minutes at 50°C, and then exposed to x ray film.
SEQUENCE ANALYSIS OF cagANEGATIVE STRAIN
The nucleotide sequence of cagA and the upstream transcript of cagA genes were analysed in one of the cagA negative strains, T-94. Preliminary analysis using 15 primers designed inside the cagA gene revealed that nucleotides from position 910 to the terminal codon were retained. Subsequently, a primer named cagA-20R (5’-TTGGTCTTTATAACCAACGG-3’ corresponding tocagA position 1020–1001) was designed. PCR was performed using cagA-20R and 10 previously described random primers.30 The amplicons were cloned into the pCRII vector using the original TA cloning kit (Invitrogen). The cloned samples were screened by colony hybridisation. The Escherichia coli colonies were transferred to a nylon membrane (Amersham International). The hybridisation was performed using the full length of thecagA gene as probe. Hybridysation and washing were performed as for the Southern blot described above. Plasmid DNA was extracted from positive colonies and the nucleotide sequence of the insert determined by the dideoxy chain termination procedure.28
STIMULATION OF IL-8 SECRETION IN GASTRIC EPITHELIAL CELL LINES
Experiments were performed as previously described.17 ,18 H pyloristrains were harvested in Brucella broth containing 7.5% FBS for 24 hours at 37°C. After centrifugation, they were resuspended at 8 × 107cells/ml in RPMI 1640 containing 10% FBS and used immediately.
MKN-28 cells were routinely maintained in RPMI 1640 supplemented with 10% FBS. Confluent monolayers of MKN-28 cells in 24 well plates were co-cultured with H pylori strains for 16 hours in quadruplicate. Supernatants were then aspirated and stored at −70°C until assayed for IL-8 by enzyme immunoassay (PerSeptive Diagnostics, Framingham, Massachusetts, USA). Concentrations of IL-8 were determined from a standard curve of recombinant IL-8. Bacterial induced IL-8 secretion was expressed as ng/ml after subtraction of background unstimulated control culture values.
STATISTICAL ANALYSIS
Differences in categorised data were analysed with Fisher’s exact probability test, and differences in mean values by analysis of variance. p<0.05 was considered significant.
Results
DETECTION OF CagA PROTEIN
CagA protein was detected by western blot analysis as previously described. Overall, 57 of 63 (90%) strains were positive for CagA protein. The prevalence of CagA protein positive strains was 12/13 for patients with gastric ulcer, 11/12 for those with duodenal ulcer, 18/22 for patients with non-ulcer dyspepsia, and 16/16 for gastric cancer patients.
SOUTHERN BLOT ANALYSIS OFcagA GENE
Southern blot analysis of chromosomal DNA derived from 63H pylori strains cleaved withHindIII revealed that all 63 (100%) strains isolated in Japan were positive for the cagAgene. Six strains that were negative for CagA protein, as determined by western blot analysis, also had the cagAgene. Some restriction fragment length polymorphism patterns were observed, but no relation was found between gastroduodenal disease and these patterns. We also analysed two cagApositive and eight cagA negative strains isolated in the Western countries as controls. Negative strains did not contain cagA gene, as expected.
NORTHERN BLOT ANALYSIS OFcagA TRANSCRIPTS
To determine whether CagA protein expression is inhibited at the transcriptional level, we analysed 17 CagA protein positive strains and six CagA protein negative strains isolated in Japan. All 17 positive strains had cagA transcripts. In the negative strains, two of six had cagAtranscripts, but the remaining four did not. ATCC 43526 and 43579 were used as positive controls, and Tx30a was used as a negative control (fig 2).

Northern blot analysis of cagA transcripts from seven strains. cagA transcripts were present in T-57 (lane 2), T-1 (lane 5), and ATCC 43526 (lane 7), and absent from T-94 (lane 1), T-85, T-25 (lanes 3 and 4), and Tx30a (lane 6). The membrane was also hybridised to 23S rRNA to monitor the amount of RNA loaded.
SOUTHERN BLOT ANALYSIS OFcagI
The H pylori isolates were analysed for the cagB, cagC,cagE (picB),cagM, cagN, and cagQ genes of thecagI region. Overall, 59/63 (94%) strains had all the cagI genes, but the remaining four (6%) lacked all six cagI genes in spite of the presence of the cagA gene. Surprisingly, four strains that lacked cagI did not have cagA transcripts and CagA protein. In the strains derived from patients with non-ulcer dyspepsia, 4/22 (18%) strains did not have cagI, whereas all 41 strains from patients with peptic ulcer and gastric cancer had cagI. This difference was statistically significant (p<0.05; Fisher’s exact probability test). ATCC 43526 and 43579 had all the genes ofcagI, and eight CagA negative strains including Tx30a lacked cagI.
SOUTHERN BLOT ANALYSIS OFcagII
We analysed cagT,cag15, cag13,cag12, cag10, andcag6–7 of cagII, and LEC. All isolates with allcagI genes (59 strains) were positive for all six cagII genes. All clinical isolates (63 strains) hybridised with cag6–7irrespective of the presence of cagI. In the analysis of cagT andcag15, strains that had cagI all had these genes, but four strains without cagI did not have cagT andcag15 genes. In the analysis ofcag13, cag12, and cag10 genes, strains that hadcagI all hadcag13, cag12, andcag10 genes, in four strains withoutcagI, three strains hadcag13, cag12, andcag10 genes, and one strain did not have these genes. ATCC 43526 and 43579 had all thecagII genes and eight CagA negative Western strains including Tx30a lacked all the cagII genes (fig 1). All 73 strains used in this study hybridised with theLEC.
SOUTHERN BLOT ANALYSIS OF IS 605
Overall, 20/63 (31%) Japanese strains had IS605. The existence of IS605 was not related to the existence ofcag PAI nor to gastroduodenal status in the host. We also investigated IS605 in eight PAI negative strains isolated in Western countries, and three of the eight (38%) were positive (table 2).
Relation between IS605 and cag region of H pylori
JUNCTIONAL SEQUENCES OF THEcag PAI DELETED REGION
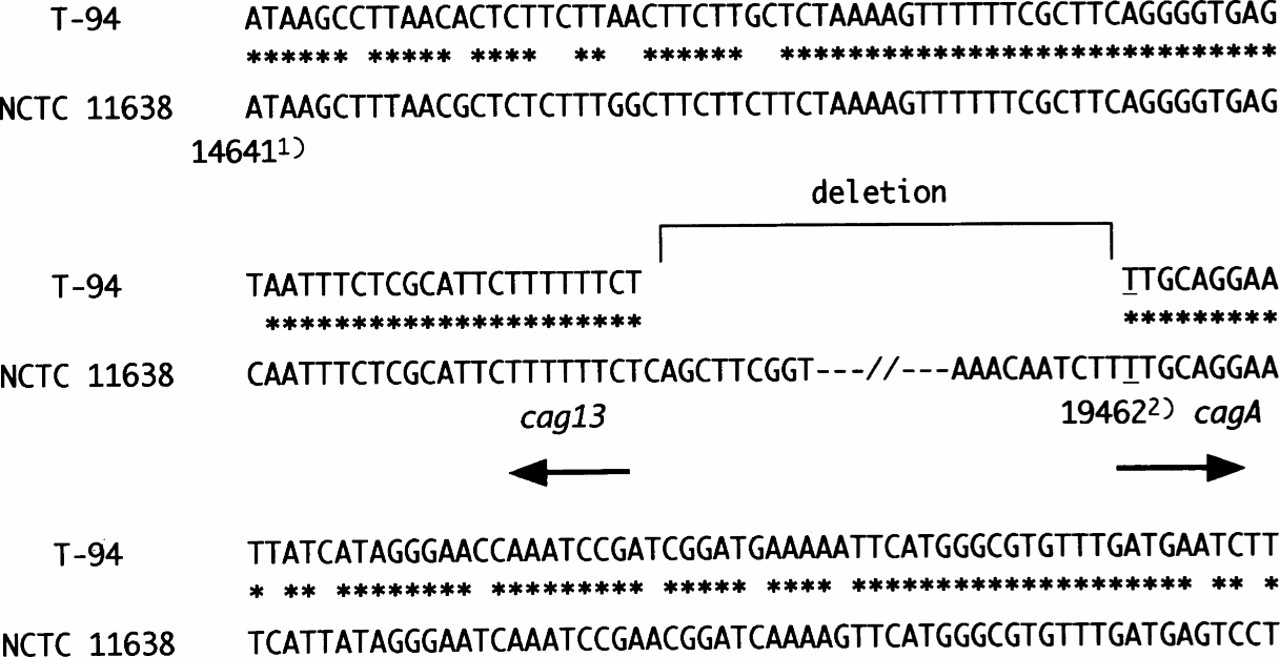
Sequence analysis of the T-94 strain revealed that thecagA gene was truncated at position 500, and the cag13 sequence was found directly upstream. About 25 kb of cag PAI including the cagA promoter region was deleted, and no insertional sequences were found in the junctional region (fig3).

Junctional sequences of T-94, which has cag PAI deleted. cagII was present down to position 14 723 (GenBank accession number AC000108) in the middle of cag13. The cagA gene was present up to position 19 462 (GenBank accession number U60176). This position is equal to position 500 of the cagA gene. An approximately 25 kb segment of cag PAI was deleted. 1) indicates the position of GenBank accession number AC000108. 2) indicates the cagA position of GenBank accession number U60176. Asterisks denote identity.
H PYLORI STIMULATION OF IL-8 SECRETION IN GASTRIC EPITHELIAL CELL LINES
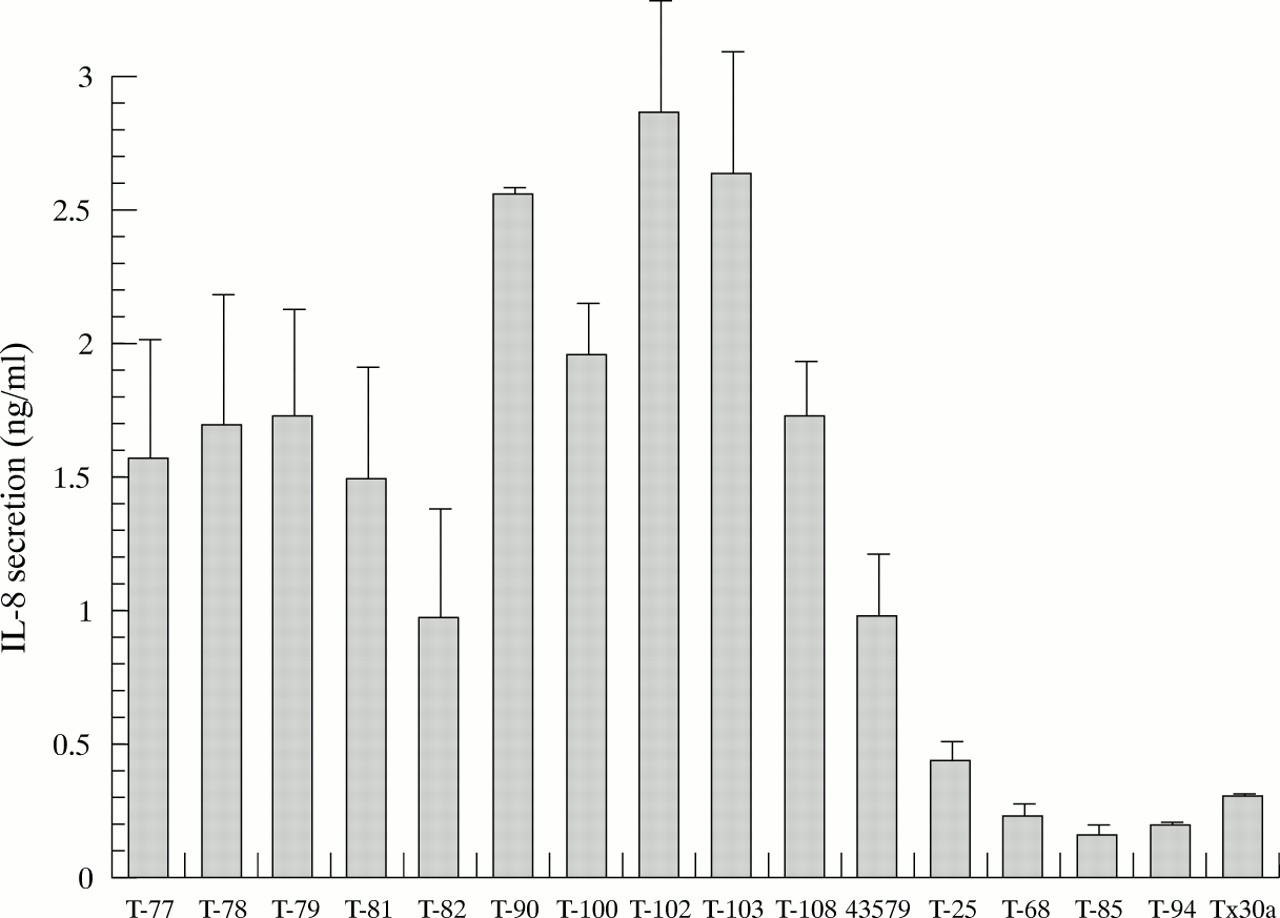
We analysed IL-8 stimulation by 16 strains: 10 clinical isolates and ATCC 43579, which were CagA protein and PAI positive; four clinical isolates (T-25, T-68, T-85, T-94), which were characterised as CagA protein negative with PAI deleted; and thecag PAI negative Tx30a. As shown in fig 4, IL-8 secretion induced by the PAI deleted strains and Tx30a was significantly lower than that induced by ATCC 43579 and the othercag PAI positive isolates (p<0.05; analysis of variance). These results indicate that intactcag PAI is necessary for the stimulation of IL-8 secretion, and the cagA gene is not an appropriate marker of IL-8 secretion.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Secretion of interleukin-8 (IL-8) from MKN-28 cells over the 16 hours after stimulation. T-77, T-78, T-79, T-81, T-82, T-90, T-100, T-102, T-103, T-108 strains were clinical isolates with an intact cag pathogenicity island (PAI). 43579 indicates ATCC 43579. Tx30a is a strain with cag PAI completely deleted. In T-25, T-68, T-85, and T-94, cag PAI was partially deleted. Results are expressed as the mean and SD from four to six experiments.
Discussion
Previous reports have used cagA status to divide H pylori strains into two main categories: CagA-producing strains which possess thecagA gene, and non-producers which lack the gene. However, a small number of strains possess the gene but do not produce the protein.31 This study investigated Japanese isolates belonging to the third category, but not onlycagA was examined but also wholecag PAI.
Japanese strains with the cag PAI partially deleted were further characterised by the absence of all the genes of the cagI region except forcagA itself and most of the genes in thecagII region. Whereas genes cag6 andcag7, located at the most upstream portion of the cagII, were retained in all of them,cag10, about 4 kb downstream of cag7, was deleted in one of four, andcag15, about 6 kb further downstream, was not detected in any. cagB, located next tocagA in the downstream portion ofcagI, and all the genes in thecag PAI fromcag15 to cagBwere deleted. Thus the nucleotide deletion in Japanesecag PAI deleted strains covers most but not all of the cag PAI, with a slight variation in the upstream border. Censini et al 21 reported that about 5–10% of strains had a partial deletion in the cag PAI. The prevalence of cag PAI deleted strains in Japan is the same as in Western countries, but it is not known whether the Western strains had a similar deletion to the Japanese strains.
Most Japanese CagA-non-producing strains lackedcagA transcripts in spite of the presence of thecagA gene, indicating that CagA expression was inhibited at the transcriptional level in these strains. This may be related to the deletion of the cagApromoter region, which is located betweencagB and cagA. We determined the sequences of the exact downstream border of the deletion in one strain and found that the promoter and the head of thecagA gene were deleted.
All strains with cag PAI partially deleted were derived from patients with non-ulcer dyspepsia (4/4), whereas only 18/59 strains with cag PAI intact were from such patients and the balance from patients with peptic ulcers or gastric cancer (p<0.05; Fisher’s exact probability test). Thus the presence of intact cag PAI appears to be a necessary condition for advanced gastroduodenal disease, and this is consistent with the in vitro finding that stimulation of IL-8 secretion from MKN-28 cells was significantly lower withcag PAI deleted strains than with intact ones. Indeed, previous studies using mutagenesis analysis have shown that many genes in the cagI and cagII regions, but not cagA itself, are essential for the induction of IL-8 secretion from epithelial cells.20 ,21 ,23
According to previous reports, the cagA gene appears to be of use as a marker of wholecag PAI genes and PAI related virulence.21 However, this study shows that the presence of the cagA gene does not necessarily indicate the presence of intact cag PAI in Japanese H pylori isolates. It remains to be investigated whether CagA-non-producing strains of this type are found outside of Japan.
Recent studies on the cagA gene in Japan, Korea, and China, where the incidence of gastric cancer is high, have shown that the prevalence of cagA positive strains is very high with no relation to gastroduodenal disease.32-34 For example, Pan et al 32 reported that 47 of 48 strains and 35 of 35 strains from Chinese patients with peptic ulcers and gastritis respectively were positive for the cagAgene. Infection with cagA positive strains is very common. However, our results indicate that genes of the PAI other than cagA should be also evaluated.
This study reveals another characteristic of JapaneseH pylori strains. IS605 is located betweencagI and cagII in the prototype H pylori, and reported to be closely associated with the presence of cagPAI.21 However, IS605 status in Japanese isolates was not related to cag PAI status: IS605 was detected in 18 of 59 (31%) strains with cagPAI intact and in two of four (50%) withcag PAI partially deleted.
In conclusion, this study shows genetic diversity between Western and Japanese H pylori strains. The non-virulent Japanese strains that do not produce CagA nevertheless retaincagA and a few other genes in thecag PAI. However, they do not induce IL-8 secretion from MKN-28 cells. Thus the presence of thecagA gene does not necessarily indicate the presence of intact cag PAI, and cannot be used invariably as a marker of cag PAI related virulence in Japanese strains.
Acknowledgments
We thank Professor Douglas E Berg for providing the information about cag pathogenicity island and his valuable suggestions. We also thank Dr John Atherton for his gift ofcagA negative strains.
Abbreviations
- IL
- interleukin
- IS605
- insertion sequence 605
- PAI
- pathogenicity island
- SDS-PAGE
- sodium dodecyl sulphate-polyacrylamide gel electrophoresis, SSC, standard saline citrate