Article Text

Abstract
BACKGROUND Oxidative and nitrosative stress have been implicated in the pathogenesis of inflammatory bowel diseases.
AIMS To study the role of nitric oxide (NO) derived from inducible NO synthase (iNOS) in an experimental model of murine enterocolitis.
METHODS Trinitrobenzene sulphonic acid (TNBS) was instilled per rectum to induce a lethal colitis in iNOS deficient mice and in wild type controls. The distal colon was evaluated for histological evidence of inflammation, iNOS expression and activity, tyrosine nitration and malondialdehyde formation (as indexes of nitrosative and oxidative stress), myeloperoxidase activity (as index of neutrophil infiltration), and tissue localisation of intercellular adhesion molecule 1 (ICAM-1).
RESULTS TNBS administration induced a high mortality and weight loss associated with a severe colonic mucosal erosion and ulceration, increased myeloperoxidase activity, increased concentrations of malondialdehyde, and an intense staining for nitrotyrosine and ICAM-1 in wild type mice. Genetic ablation of iNOS gene conferred to mice a significant resistance to TNBS induced lethality and colonic damage, and notably reduced nitrotyrosine formation and concentrations of malondialdehyde; it did not, however, affect neutrophil infiltration and intestinal ICAM-1 expression in the injured tissue.
CONCLUSION Data show that activation of iNOS is required for nitrosative and oxidative damage in experimental colitis.
- nitric oxide
- nitric oxide synthase
- inflammatory bowel disease
- intercellular adhesion molecule 1
- malondialdehyde
- nitrotyrosine
Abbreviations used in this paper
- IBD
- inflammatory bowel disease
- ICAM-1
- intercellular adhesion molecule 1
- iNOS
- inducible nitric oxide synthase
- NO
- nitric oxide
- TNBS
- trinitrobenzene sulphonic acid
Statistics from Altmetric.com
- nitric oxide
- nitric oxide synthase
- inflammatory bowel disease
- intercellular adhesion molecule 1
- malondialdehyde
- nitrotyrosine
Under physiological conditions in the gastrointestinal tract, nitric oxide (NO) from constitutive NO synthase plays an important homoeostatic role. The constitutive release of NO, presumably by intestinal epithelial or lamina propria cells and neuronal terminals, is critical to the preservation of normal intestinal permeability and regulation of neurotransmission and motility.1-3 NO also regulates neutrophil recruitment by inhibiting the expression of adhesion molecules, including intercellular adhesion molecule 1 (ICAM-1) in the vascular endothelium.4 Increased NO production by inducible NO synthase (iNOS) has been proposed to be responsible for tissue injury in various experimental models,5-13 and ulcerative colitis in humans, where iNOS activity and raised concentrations of luminal nitrite have been detected in rectal dialysates and in biopsy specimens.14-17 However, the critical contribution of iNOS to the pathogenesis of inflammatory bowel disease (IBD) has not been clearly delineated.
It has been recently proposed that the cytotoxic effects of NO are mediated in part by peroxynitrite, a potent oxidant produced by the reaction of NO and superoxide anion.18 As a highly toxic reactive species, peroxynitrite indiscriminately attacks biomolecules critical to function and viability of the cell.19 During IBD the simultaneous production of superoxide and NO is likely to produce peroxynitrite and to promote oxidative reactions. Biochemical evidence for the formation of peroxynitrite has been provided in trinitrobenzene sulphonic acid induced ileitis in guinea pigs by immunohistochemical staining of nitrotyrosine, a marker of peroxynitrite induced protein modification, in epithelial cells.20 Furthermore, intracolonic administration of exogenous peroxynitrite produced severe mucosal damage in rats.21 In support of these findings, experimental studies have shown that the inflammatory response may be reduced by administration of NOS inhibitors.6-13 In contrast to these findings, however, it has been reported that genetic ablation of iNOS activation may exacerbate intestinal inflammation induced by intraluminal administration of acetic acid in mice.22Other experimental studies have shown that only slight pharmacological inhibition of NO formation reduced colonic lesions, while a complete abolition of NO synthesis resulted in increased mucosal damage.23
To clarify further the biological role of activation of iNOS in intestinal inflammation, we induced colitis in mice lacking a functional gene for iNOS (iNOS−/−), while maintaining the capability to produce NO constitutively.24Specifically, we evaluated the relation between colonic histological alterations, nitrosative and oxidative damage, and iNOS activation using an experimental model of IBD induced by intracolonic administration of TNBS in 50% ethanol.25 We chose this model of IBD because it has macroscopic, histological, and biochemical alterations which are related to the production of free radicals, oxidants, and NO and are similar to the pathogenesis of colitis, in particular Crohn’s disease.26 ,27
Methods
ANIMALS
Male and female iNOS−/− mice and iNOS+/+ littermates (129/Sv × C57BL10, 20–22 g) were from breeding pairs generated as previously described.24The animals were a kind gift of Dr Laubach (Glaxo Wellcome Research Institute, Department of Molecular Biochemistry, Research Triangle Park, North Carolina). The experiments were carried out in accordance with National Institutes of Health guidelines and with the approval of the Institutional Review Board of the Children’s Hospital Research Foundation, Cincinnati, Ohio.
INDUCTION OF EXPERIMENTAL COLITIS
Colitis was induced using the technique previously described.25 In fasted mice lightly anaesthetised with isoflurane, a 3.5F catheter was inserted into the colon via the anus until approximately the splenic flexure (4 cm from the anus). 2,4,6-Trinitrobenzene sulphonic acid (TNBS, 1 mg/mouse) was dissolved in 50% ethanol (vol/vol) and injected (0.1 ml) into the colon via the rubber cannula. Control animals received 50% ethanol alone. Animals were then kept in a vertical position for 30 seconds and returned to their cages. In the first experiment, animals were monitored for appearance of diarrhoea, loss of body weight, and survival for seven days. In a second set of experiments, groups of animals were sacrificed every 24 hours for seven days after TNBS administration; blood samples were collected by cardiac puncture, and a segment of the colon 4 cm long was excised for macroscopic damage evaluation. Tissue segments were then immediately frozen in liquid nitrogen and stored at −70°C for the histological and immunohistochemical studies and for determination of myeloperoxidase activity and malondialdehyde concentrations.
MEASUREMENT OF PLASMA NITRITE/NITRATE CONCENTRATION
Nitrite/nitrate production, an indicator of NO synthesis, was measured in plasma samples as previously described.28Nitrate in the plasma was reduced to nitrite by incubation with nitrate reductase (670 mU/ml) and NADPH (160 mM) at room temperature for three hours. After three hours, nitrite concentration in the samples was measured by the Griess reaction, by adding 100 μl of Griess reagent (0.1% naphthalethylenediamine dihydrochloride in H2O and 1% sulphanilamide in 5% concentrated H3PO4; vol 1/1) to 100 μl samples. The optical density at 550 nm (OD550) was measured using a Spectramax 250 microplate reader (Molecular Devices Sunnyvale, California). Nitrate concentrations were calculated by comparison with OD550 of standard solutions of sodium nitrate prepared in saline solution.
EVALUATION OF COLONIC DAMAGE
After removal, the colon was gently rinsed with saline solution, opened by a longitudinal incision, and immediately examined under a microscope. The visible colonic damage was assessed by a semiquantitative scoring system adapted to the murine model.27 ,29 The following morphological criteria were taken into consideration: score 0, no damage; score 1, localised hyperaemia without ulcers; score 2, linear ulcers, with no significant inflammation; score 3, linear ulcers with inflammation at one site; score 4, two or more major sites of ulceration and/or inflammation; score 5, two or more sites of inflammation and ulceration extending more than 1 cm along the length of the colon; score 6–8, one point is added for each cm of ulceration beyond an initial 2 cm. All measurements of damage were performed by two observers blinded to the experimental protocol.
HISTOPATHOLOGICAL ANALYSIS
For microscopic histological evaluation, formalin fixed tissues were embedded in paraffin; 5 μm sections were stained with haematoxylin and eosin and evaluated by light microscopy by a pathologist unaware of the experimental protocol.
IMMUNOHISTOCHEMISTRY FOR iNOS
Frozen sections (5 μm) were treated with 0.3% hydrogen peroxide for 15 minutes to block endogenous peroxidase activity and then rinsed briefly in phosphate buffered saline (PBS). Non-specific binding was blocked by incubating the slides with a blocking solution (0.1 M PBS containing 0.1% Triton X-100 and 2% normal goat serum) for two hours. To detect iNOS, rabbit polyclonal anti-iNOS antibody was applied in a dilution of 1/2000 at 4°C overnight. Control sections included buffer alone or non-specific purified rabbit IgG. Immunoreactivity was detected with a biotinylated goat antirabbit secondary antibody and the avidin-biotin-peroxidase complex (Vectastain Elite ABC kit, Vector Laboratories). Colour was developed using diaminobenzidine.20
IMMUNOHISTOCHEMISTRY FOR NITROTYROSINE
Tyrosine nitration, a marker of nitrosative damage, was measured in colonic sections by immunohistochemistry.20 Frozen sections (5 μm thick) were fixed in 4% paraformaldehyde and incubated for two hours with a blocking solution (0.1 M PBS containing 0.1% Triton X-100 and 2% normal goat serum) in order to minimise non-specific adsorption. Sections were then incubated overnight with 1/1000 dilution of primary antinitrotyrosine antibody or with control solutions. Controls included buffer alone or non-specific purified rabbit IgG. Specific labelling was detected by incubating for 30 minutes with a biotin conjugated goat antirabbit IgG and amplified with avidin-biotin peroxidase complex (Vectastain Elite ABC kit, Vector Laboratories) after quenching endogenous peroxidase with 0.3% H2O2 in 100% methanol for 15 minutes. Diaminobenzidine was used as a chromogen. To quantitate the degree of nitrotyrosine staining, a 0–4 grading system was used: 0, no staining; 1–3, increasing degrees of intermediate staining; 4, extensive staining. In each experimental group, five or six sections were evaluated by two independent observers blinded to the experimental protocol.
ASSAY OF MYELOPEROXIDASE ACTIVITY
Myeloperoxidase activity was determined as the index of neutrophil accumulation.30 Colonic tissues were homogenised in a solution containing 0.5% hexadecyltrimethyl-ammonium bromide dissolved in 10 mM potassium phosphate buffer (pH 7) and centrifuged for 30 minutes at 20 000 g at 4°C. An aliquot of the supernatant was then allowed to react with a solution of tetramethylbenzidine (1.6 mM) and 0.1 mM H2O2. The rate of change in absorbance was measured by spectrophotometry at 650 nm. Myeloperoxidase activity was defined as the quantity of enzyme degrading 1 μmol/min of peroxide at 37°C and was expressed in milliunits per 100 mg weight of tissue.
IMMUNOHISTOCHEMISTRY FOR ICAM-1
ICAM-1 expression was evaluated in colon sections by immunohistochemistry.31 Frozen sections (5 μm thick) were fixed in 4% paraformaldehyde and incubated in 2% hamster serum for two hours in order to minimise non-specific adsorption. Sections were then incubated overnight at 4°C with monoclonal biotinylated antibodies directed at ICAM-1 (hamster antimouse CD54) at a dilution of 1/500. Controls included buffer alone or non-specific purified IgG. Antibody binding sites were visualised with an avidin-biotin peroxidase complex immunoperoxidase technique (Vector Laboratories) after quenching endogenous peroxidase with 0.3% H2O2in 100% methanol for 15 minutes. Diaminobenzidine was used as a chromogen. To quantitate the degree of ICAM-1 staining, a 0–4 grading system was used: 0, no staining; 1, constitutive presence of staining along endothelial wall; 2, increased staining along the endothelial wall; 3, increased staining along the endothelial wall, and presence of staining on infiltrated inflammatory cells; 4, increased staining along the endothelial wall, and presence of staining on infiltrated inflammatory cells and epithelial cells.
MEASUREMENT OF COLONIC MALONDIALDEHYDE
Tissue concentrations of malondialdehyde were determined as an index of lipid peroxidation.32 Tissue samples were homogenised in 1.15% KCl solution. An aliquot (100 μl) of the homogenate was added to a reaction mixture containing 200 μl of 8.1% sodium dodecyl sulphate (SDS), 1500 μl of 20% acetic acid (pH 3.5), 1500 μl of 0.8% thiobarbituric acid, and 700 μl distilled water. Samples were then boiled for one hour at 95°C and centrifuged at 3000g for 10 minutes. The absorbance of the supernatant was measured by spectrophotometry at 532 nm and compared with a standard curve obtained with 1,1,3,3-tetramethoxypropane. Data were expressed as μM per 100 mg weight of tissue.
MATERIALS
Primary iNOS and antinitrotyrosine antibodies were purchased from Upstate Biotech (Saranac Lake, New York). Primary monoclonal ICAM-1 (CD-54) antibody was purchased from Pharmingen (San Diego, California). Hamster serum was purchased from Jackson ImmunoResearch Laboratories (West Grove, Pennsylvania). Reagents, secondary, and non-specific IgG antibodies for immunohistochemical analysis were from Vector Laboratories Inc. (Burlingame, California). All other chemicals were from Sigma/Aldrich (St Louis, Missouri).
STATISTICAL ANALYSIS
All data are expressed as mean (SEM); n refers to the number of mice. Statistical differences between groups were calculated by one and two way analysis of variance (ANOVA) followed by a Bonferroni post hoc test. Survival differences were analysed with the χ2test. Results were considered significant at a p value of less than 0.05.
Results
SEVERITY OF TNBS INDUCED COLITIS IS REDUCED IN iNOS−/− MICE
Intracolonic administration of TNBS in wild type mice induced a severe illness characterised by bloody diarrhoea and a dramatic loss of body weight and resulted in a high mortality rate (only two mice out of 20 survived seven days after TNBS treatment). In contrast, iNOS−/− mice seemed notably less sensitive to the inflammatory effects of TNBS: animals experienced a mild diarrhoea and a less pronounced loss of body weight, and only six animals out of 16 died during the experimental observation period (fig 1A, B).

Survival (A), weight loss (B), and damage score (C) in iNOS+/+ and iNOS−/− mice after TNBS intracolonic administration. Each data point is the mean (SEM) of 8–20 animals for each group. *p<0.05, **p<0.01.
TNBS INDUCED COLONIC DAMAGE IS ATTENUATED IN iNOS−/− MICE
Macroscopic evaluation of the distal colon and rectum up to 72 hours after TNBS treatment revealed the presence of mucosal oedema and haemorrhagic ulcerations in both wild type and iNOS−/−mice. In wild type mice mucosal damage was still severe up to six and seven days after TNBS administration. However, at four days after colitis induction, in iNOS deficient mice, the mucosal surface of the colon and rectum showed only localised erythema or no damage, indicative of a significant resolution of colitis in comparison to the wild type mice (p<0.05; see fig 1C for damage score). Histological examination confirmed the above observations (fig 2). In colonic sections from iNOS+/+ mice, haemorrhagic necrosis of the mucosa was associated with infiltration of inflammatory cells into the lamina propria and submucosa. Histopathological evaluation of the colon and rectum of TNBS treated iNOS−/− mice showed a remarkable reduction in mucosal damage with recovering process of the epithelium starting at four days after induction of colitis. However, a massive infiltration of inflammatory cells was still present in the lamina propria and submucosa of colonic sections of TNBS treated iNOS−/− mice (fig 2).

Time course of changes of colonic epithelial architecture after TNBS administration. Representative colonic sections from non-treated iNOS+/+ (A) or iNOS−/− (E) mice show normal tissue structure at day 0. (B) and (C) show a notable disruption of the epithelial structure with extensive haemorrhagic necrosis and infiltration of neutrophils in a colonic section from an iNOS+/+ mouse at two and four days after TNBS administration, respectively. At day 7 (D) oedema was still present in the healing epithelium of a TNBS treated iNOS+/+ mouse. (F) shows alteration of epithelial architecture with a massive infiltration of inflammatory cells in a colonic section of an iNOS−/− mouse two days after TNBS administration. At days 4 (G) and 7 (H) inflammatory cells were still present, mainly in the submucosa, while a healing process started in the epithelium. Original magnification × 400.
TNBS ADMINISTRATION INDUCES iNOS EXPRESSION IN WILD TYPE MICE
Induction of colitis in TNBS treated wild type mice was associated with expression of iNOS, which was localised mainly at the apical part of the epithelial cells, as determined by immunohistochemistry. Expression of iNOS was associated with a remarkable increase in plasma nitrite/nitrate concentration. In the iNOS knockout animals, low nitrite/nitrate concentrations were detected with no detectable iNOS expression (fig 3).

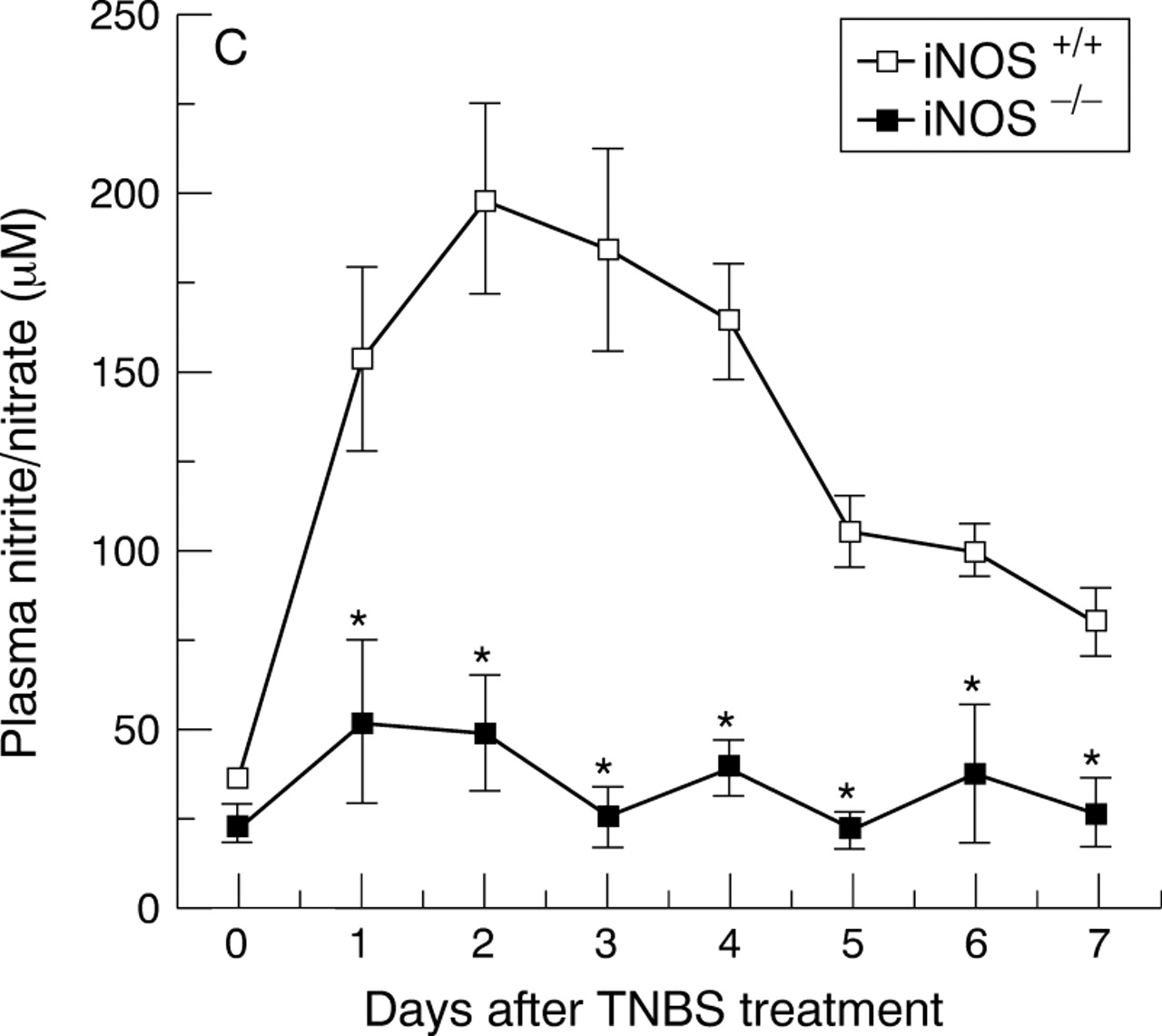
Immunohistochemical localisation of iNOS (A and B) and plasma nitrite/nitrate concentrations (C). (A) shows a diffuse dark staining localised in the apical epithelium (arrows) in the inflamed colon of an iNOS+/+ mouse four days after TNBS administration. (B) shows no staining in a colonic section of an iNOS−/− mouse four days after TNBS administration. Original magnification × 400. (C) shows the time course of plasma nitrite/nitrate concentrations after TNBS administration. Each data point is the mean (SEM) of 4–8 animals for each group. *p<0.05.
COLONIC INFILTRATION OF NEUTROPHILS AND ICAM-1 EXPRESSION WAS NOT AFFECTED IN iNOS−/− MICE AFTER TNBS ADMINISTRATION
As abrogation of colonic damage was not paralleled by a reduction in inflammatory cells, we next sought to estimate better the infiltration of neutrophils by quantification of tissue activity of myeloperoxidase, a specific enzyme of azurophilic granules and directly correlated to the number of neutrophils. In parallel with increased infiltration of inflammatory cells observed in the histological specimens, myeloperoxidase activity was significantly increased in TNBS treated iNOS+/+ mice after TNBS treatment. Similar high concentrations of myeloperoxidase activity were found in TNBS treated iNOS−/− mice (fig 4A).

Time course of myeloperoxidase activity (A) and score of immunostaining of ICAM-1 (B) in tissues from iNOS+/+ and iNOS−/− mice after induction of colitis. Each data point is the mean (SEM) of eight animals for each group.
We also evaluated the intestinal expression of ICAM-1. In colonic tissue from iNOS+/+ or iNOS−/− control mice at day 0, immunostaining for ICAM-1 was present along the endothelium of small vessels of the lamina propria and submucosa, consistent with a constitutive expression of ICAM-1 on endothelial cells (figs 4B and 5). After TNBS administration, the staining intensity for ICAM-1 increased along the endothelium and was found also on epithelial cells and neutrophil surface with similar distribution and extent in both iNOS+/+ and iNOS−/− mice (figs 4B and5).
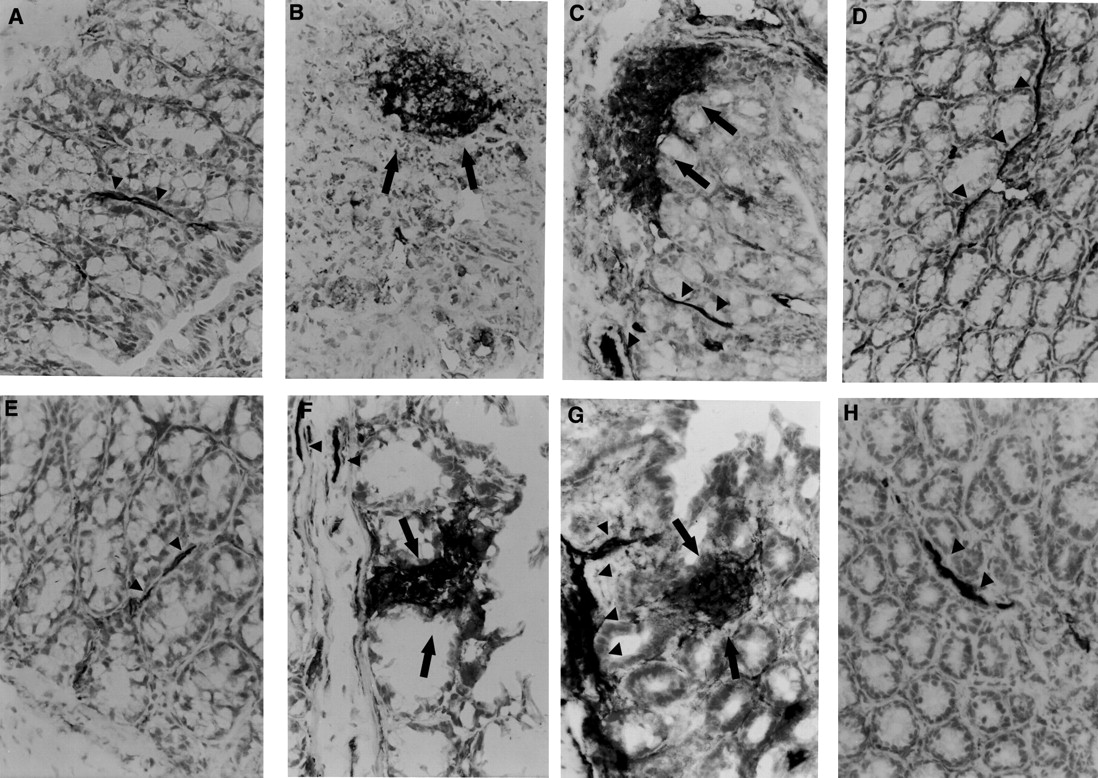
Time course of ICAM-1 expression after TNBS induced colitis. Control tissues at day 0 from non-treated animals iNOS+/+ (A) and iNOS-/- (E) showed a dark brown staining of endothelium of blood vessels (arrow heads) indicating the presence of constitutive ICAM-1 protein. TNBS administration induced an increase in the positive staining for ICAM-1 along the endothelial vascular wall (arrow heads) in the submucosa area and in infiltrated cells (arrows) at days 2 and 4 after TNBS administration in iNOS+/+ (B and C) and iNOS−/− mice (F and G). A representative microphotograph at day 7 shows the presence of constitutive ICAM-1 staining in vessels (arrow heads) of the healing mucosa in iNOS+/+ (D) and iNOS−/− mice (H). Original magnification × 400.
NITROSATIVE AND OXIDATIVE DAMAGE WAS REDUCED IN TNBS TREATED iNOS−/− MICE
It has been suggested that neutrophils contribute significantly to the mucosal damage of colitis by releasing radicals and oxidant molecules.33 ,34 In the present study, we found raised tissue concentrations of malondialdehyde, indicative of a notable lipid peroxidation (fig 6A). An intense immunostaining for nitrotyrosine was indicative of a nitrosative injury in TNBS treated iNOS+/+wild type mice (figs 6B and 7). Surprisingly, genetic deficiency of iNOS in TNBS treated mice prevented the increased formation of malondialdehyde and nitrotyrosine staining, even in the presence of a massive neutrophil infiltration (figs 6B and7).

Time course of colonic concentrations of malondialdehyde (A) and score of immunostaining of nitrotyrosine (B) in tissues from TNBS treated iNOS+/+ and iNOS−/− mice. Each data point is the mean (SEM) of eight animals for each group. *p<0.05.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immunohistochemistry for nitrotyrosine. In colonic sections from non-treated mice (day 0) no staining for nitrotyrosine was found in iNOS+/+ (A) and iNOS−/− mice (E). At days 2 (B) and 4 (C) after TNBS administration a diffuse dark staining was localised in infiltrated inflammatory cells and in the necrotic epithelium of an iNOS+/+mouse. At day 7 (D) nitrotyrosine staining was still present in the apical epithelium in the area of healing mucosa of an iNOS+/+mouse. Nitrotyrosine staining was absent at days 2 (F), 4 (G), and 7 (H) after TNBS administration in the mucosa of an iNOS−/− mouse. Original magnification × 400.
Discussion
Our data show that mice with a targeted deletion of the iNOS gene are significantly more resistant to death and pathological changes in the colon and rectum associated with TNBS induced colitis compared with wild type controls. Thus, these results suggest that the development and persistence of colonic injury require the presence of a functional iNOS gene and iNOS generated NO.
ACTIVITY OF iNOS IS A CRUCIAL PREREQUISITE OF OXIDATIVE AND NITROSATIVE DAMAGE IN TNBS INDUCED COLITIS
It has been proposed that reactive oxygen and nitrogen species play a key role in IBD. These species are cytotoxic agents, inducing lipid peroxidation and other cellular oxidative stress by cross linking proteins, lipids, and nucleic acids, which then cause cellular dysfunction, damage, and eventually death. Evidence consistent with damage by reactive species is provided by the increase in lipid peroxides in rectal biopsy specimens from patients with ulcerative colitis.35-37 In our experiments we found that the mucosal damage induced by intracolonic administration of TNBS in wild type mice (that is, those with a normal iNOS gene), was associated with high concentrations of colonic thiobarbituric acid reactant malondialdehyde, which is considered a good indicator of lipid peroxidation.32
Recent evidence indicates also that nitration of tyrosine can result from a number of chemical reactions, and can be considered as a global marker of nitrosative stress.19 ,38 Nitrotyrosine can be formed from the reaction of nitrite with hypochlorous acid or the reaction of nitrite with myeloperoxidase and hydrogen peroxide.38-40 In our experiments, we found increased immunohistochemical expression of iNOS, mainly localised in the apical side of epithelial cells. This finding is supported by other reports showing that iNOS staining is always at the superficial part of the crypt and the surface of the mucosa, while no other positive cells or part other than apical were found in human colonic sections of ulcerative colitis.41 ,42 Furthermore, we found a positive immunohistochemical staining for nitrotyrosine mostly on epithelial cells and in the area of infiltrated inflammatory cells, suggesting that peroxynitrite or other nitrogen derivatives and oxidants are formed in vivo and may contribute to tissue injury. These data are consistent with previous findings that immunohistochemical staining for nitrotyrosine was colocalised on epithelial cells with iNOS immunoreactivity in a TNBS model of guinea pig ileitis20and in active Crohn’s lesions in humans.43 The pathogenic role of nitrogen derived species such as peroxynitrite in IBD is further supported by the fact that intracolonic administration of exogenous peroxynitrite induces a severe colonic inflammation which mimics the features of both ulcerative colitis and Crohn’s disease.21
We observed that epithelial disruption was similar in wild type and iNOS−/− mice at the earlier time (up to 72 hours after TNBS instillation). However, iNOS deficiency prevented the formation of tissue malondialdehyde and nitrotyrosine staining in TNBS treated animals. Furthermore, iNOS−/− mice seemed more resistant to TNBS induced lethal disease with a significant resolution of the macroscopic and histological signs of the inflammatory process at the later time. Therefore, although the lack of iNOS activity does not prevent the non-specific chemical induced colonic damage secondary to the caustic nature of TNBS, it prevents the persistence of damage due to the involvement of NO driven inflammatory pathways. Thus, these findings, coupled with the inhibitor effects on nitrotyrosine formation and lipid peroxidation, suggest that the protective effect of iNOS gene ablation is probably due to reduced production of toxic oxidants during the inflammatory process. It has been proposed, however, that peroxynitrite formation may also be produced from the reaction of superoxide anion with constitutive NOS derived NO.44 Our study, showing a notable reduction in malondialdehyde and nitrotyrosine in iNOS−/− mice, which still maintain the capability to produce NO from the constitutive enzyme,24 suggests that an overproduction of NO and peroxynitrite and/or other nitrogen derivatives from iNOS plays the major role in the oxidative and nitrosative stress of IBD.
THE BENEFICIAL EFFECTS OF GENETIC ABLATION OF iNOS IN TNBS INDUCED COLITIS ARE NOT RELATED TO AN ALTERATION IN NEUTROPHIL RECRUITMENT
Neutrophils have been considered to play a crucial role in the development and full manifestation of gastrointestinal inflammation, as they represent a major source of free radicals in the inflamed colonic mucosa.33-35 Neutrophil infiltration seems to be mediated, at least in part, by an increase in the expression of ICAM-1 on endothelial cells, neutrophils, and epithelial cells.45-49 In this regard, NO functions as an inhibitor of the neutrophil-endothelial cell interaction, most probably by inhibiting expression of adhesion molecules on the endothelial surface.4 A major finding of this study was that, although the iNOS−/− mice showed a remarkable recovery of the mucosal morphology associated with a reduction in oxidative and nitrosative damage after TNBS administration, infiltration of polymorphonuclear neutrophils was similar in tissue extent and distribution to that seen in TNBS treated iNOS+/+ mice. Furthermore, ICAM-1 was expressed in endothelial and epithelial cells, and neutrophils in the distal colon in both TNBS treated wild type and iNOS−/− mice. Thus, in contrast with previous findings showing that genetically iNOS deficient mice have enhanced leucocyte-endothelium interactions in endotoxaemia,50 our data suggest that activation of inducible NOS does not contribute to the regulation of neutrophil infiltration. Notably, Khanet al reported that iNOS−/−mice subjected to Toxoplasma gondiiinfection showed a notable reduction in hepatocellular dysfunction and necrosis of the small intestine but had no difference in neutrophil infiltration, compared with parenteral controls.51
Substantial evidence supports the view that the maintenance of the constitutive production of NO is a critical homoeostatic regulator for maintaining microvascular patency, permeability, tissue perfusion, and mucosal barrier function, and for inhibiting neutrophil infiltration and activation.1-3 In this regard, it is noteworthy that in the gastrointestinal tract, acute non-selective blockade of constitutive and induced NO synthesis augments leucocyte adhesion to the microvascular endothelium3 and exacerbates the damage associated with ethanol insult52 and intracolonic administration of TNBS.23
ACTIVATION OF iNOS AMPLIFIES THE OXIDANT DEPENDENT PATHWAY INJURY IN THE INFLAMED COLONIC MUCOSA
The findings presented here clearly illustrate that, in spite of a massive neutrophil infiltration, which is expected to maintain the production of oxygen derived free radicals, oxidative and nitrosative damage in IBD requires the presence of a functional iNOS gene. In accordance with our findings, previous studies have shown that infiltration of inflammatory cells was not essential for the development of the major pathological features of colitis induced by phorbol myristate acetate, acetic acid, or TNBS,53 and neutrophil depletion failed to attenuate colonic injury.53-55 Several hypotheses may explain this phenomenon. Although NO itself is a weak oxidant, biochemical studies have shown that NO rapidly interacts with superoxide anion to yield peroxynitrite and its conjugate acid, peroxynitrous acid, which then decomposes to form highly reactive oxidant species, such as hydroxyl and nitrogen dioxide. Both NO and peroxynitrite, directly or via reactive intermediates, may induce cytotoxicity via a number of mechanisms including tyrosine nitration, lipid peroxidation, DNA strand breaks, and the consequent activation of poly-ADP ribose synthase.19 ,28 ,56-60 Furthermore, nitrogen centred oxidants may also degrade the iron-sulphur clusters of mammalian mitochondrial NADH-coenzyme Q reductase, succinate dehydrogenase, aconitase, and ATPase.57 ,61 As a consequence, intracellular iron homoeostasis may be disrupted, providing a catalyst for further generation of deleterious oxygen species from neutrophils.62
Conclusions
Utilising a murine iNOS deficient model of chemically induced colitis, we have obtained strong evidence implicating iNOS as a critical element in the pathogenesis of inflammatory bowel injury. Our data are in general agreement with previous pharmacological studies of experimental colitis, in which NOS inhibition has had a favourable effect on mucosal injury.6-13 In contrast to these previous reports and our present study, McCaffertyet al reported a deleterious effect of iNOS deficiency on the ability to resolve a colitic injury in an experimental model of inflammatory bowel disease induced by rectal instillation of acetic acid.22 The discrepancy in these reports may relate to the difference in the stimuli used to induce the injury. Although the acetic acid and TNBS induced colitis models reproduce many of the characteristic features of human IBD, there are significant differences in the extension of intestinal damage, the time requirement to develop lesions, and the relative involvement of iNOS independent pathways.26 It is noteworthy that similar controversial roles have been ascribed to iNOS derived NO in a variety of pathophysiological conditions. While a number of studies have reported that genetic ablation of iNOS may exert beneficial effects in endotoxic shock, infection by Toxoplasma gondii, and autoimmune vasculitis,51 ,63-65 other studies have reported that genetic ablation of iNOS may exacerbate the inflammatory process in endotoxaemia, encephalomyelitis, and tuberculosis.24 ,66 ,67 Further experiments are needed in order to explain these differences, which have important implications for clinical therapeutic strategies based on inhibition.
Furthermore, TNBS induced colitis may not represent a suitable model for investigatation of the pathophysiology of human Crohn’s disease or ulcerative colitis, which are spontaneous and heterogeneous disorders with multiple aetiological and pathogenic mechanisms. Nevertheless, the TNBS induced colitis model reproduces many of the characteristic features of human IBD, including generation of various inflammatory mediators, generation of oxygen and nitrogen radicals and oxidants, and immunological alteration.26 ,27 Therefore, it is conceivable that the mechanisms responsible for IBD in animals and humans are notably different. Nevertheless, based on the current data, we propose that the oxidative and nitrosative changes and the lethality associated with TNBS induced murine colitis require iNOS activation. Together with previous reports, our study provides further evidence of the critical role of iNOS derived NO in the pathogenesis of colitis in experimental animals. However, whether iNOS activation has a role in the immunoregulatory and inflammatory processes in humans needs further investigation.
Abbreviations used in this paper
- IBD
- inflammatory bowel disease
- ICAM-1
- intercellular adhesion molecule 1
- iNOS
- inducible nitric oxide synthase
- NO
- nitric oxide
- TNBS
- trinitrobenzene sulphonic acid