Article Text

Abstract
BACKGROUND Hepatocellular carcinoma (HCC) arising in cirrhosis is frequently multifocal. Whether HCC develops monoclonally or multiclonally is an unresolved question. Of the multiple tumour nodules present in many patients, it has not been established whether the smaller lesions represent intrahepatic metastases or de novo cancers.
AIMS To assess the degree of genomic heterogeneity in synchronous HCCs in cirrhosis.
METHODS The arbitrarily primed polymerase chain reaction technique was utilised to compare the DNA fingerprint of HCCs and regenerative nodules (RNs) removed from cirrhotic explant livers.
RESULTS Polymorphic genomic heterogeneity was noted in 54 HCCs and 31 RNs microdissected. Even satellite nodules in close proximity within the same segment of the liver were found to have distinct genomic patterns.
CONCLUSION Such genomic heterogeneity in synchronous HCCs may explain poor patient survival after surgical resection. If the smaller tumours are de novo lesions rather than metastases (as these data suggest), then current concepts regarding liver resection as a curative treatment modality for HCC may require reassessment.
- hepatocellular carcinoma
- regenerative nodule
- cirrhosis
- DNA fingerprint
- arbitrarily primed polymerase chain reaction
- genomic heterogeneity
Abbreviations used in this paper
- AP-PCR
- arbitrarily primed polymerase chain reaction
- HBV
- hepatitis B virus
- HCC
- hepatocellular carcinoma
- MRN
- macroregenerative nodule
- RN
- regenerative nodule
Statistics from Altmetric.com
- hepatocellular carcinoma
- regenerative nodule
- cirrhosis
- DNA fingerprint
- arbitrarily primed polymerase chain reaction
- genomic heterogeneity
Hepatocellular carcinoma (HCC) is one of the most common malignancies worldwide, with an annual incidence of a quarter of a million cases.1 The profile of risk factors associated with HCC includes not only chronic infection with hepatitis B virus (HBV) and/or hepatitis C virus with subsequent cirrhosis, but also metabolic and alcoholic chronic liver diseases. In the cirrhotic livers that harbour HCC,2 the tumours are frequently multifocal. Most HCCs are large and symptomatic at presentation, and generally carry a poor prognosis. The number of patients presenting with surgically resectable tumours remains low at 10–15% of the total patient population with primary HCC worldwide. Recurrence is commonly observed in the cirrhotic liver, rates of up to 80% being reported within five years of resection.3 Screening of cirrhotic patients for the detection of small HCCs may improve prognosis of this disease.4 However, even if suspected lesions are detected and biopsied, small HCCs are still difficult to diagnose morphologically by conventional pathological methods, especially when they are less than 1.5 cm diameter. In the setting of cirrhosis, there are no agreed criteria for the diagnosis of these tumours. The smallest HCCs begin as well differentiated tumours, which generally dedifferentiate and become heterogeneous as they grow above 1.5 cm.5 Our inability to identify them at this size is likely to limit the effectiveness of treatment.
Whether HCC in human liver develops from a single clone or from multiple parallel clones is an unresolved question. Information on the clonal origin of tumours will influence management strategies for prevention of recurrence after operation. Various techniques have been developed to assess the clonality in tumours, such as the detection of alterations in oncogenes or tumour suppressor genes,6 ,7integration of HBV into the cellular genome,8 ,9 X linked restriction fragment length polymorphisms within the tumour,10 analysis of DNA ploidy pattern,11and the demonstration of a specific cytogenetic abnormality. However, only a small fraction of HCC exhibit point mutations of specific oncogenes or tumour suppressor genes, and integrated HBV DNA can only be used as a marker in those patients who have viral infection. X linked restriction fragment length polymorphisms are limited to tumours in women, while the analysis of DNA content does not detect the cell with a balanced gain or loss of chromosomes, or with genetic rearrangements.
Recently the arbitrarily primed polymerase chain reaction (AP-PCR) method was developed as a technique which generates an information rich and unbiased fingerprint of genomic DNA without requiring prior knowledge of nucleotide sequence information.12 It is based on the selective amplification of genomic sequences that, by chance, are flanked by adequate matches to an arbitrarily chosen primer. This method utilises amplification with a single arbitrary primer and it has been applied to detect and isolate DNA fragments of somatic genomic changes that show losses or gains in cancer.13 Moreover, it is a highly specific technique that is able to generate species specific and individual diagnostic DNA fingerprints.14 Of the multiple tumour nodules present in many patients, it has not been established whether the smaller lesions represent intrahepatic metastases or de novo cancers. We therefore assessed the degree of genomic heterogeneity in synchronous HCCs using AP-PCR. If two template genomic DNA sequences are different, their arbitrarily primed PCR products display different fingerprinting patterns.
Materials and methods
TISSUE SAMPLES
Tissue samples used in this study were cirrhotic explant livers from patients undergoing elective orthotopic liver transplantation for end stage chronic liver disease at the Royal Free Hospital. The formalin fixed livers were cut into coronal slices 10 mm thick and closely examined for unusual nodules. If any unusual nodules were found, tissue samples were taken from these nodules, embedded in paraffin wax, and processed using standard techniques. The details of nodule selection and pathological categorisation have been described previously.15
One 5 μm section and ten 10 μm sections were serially cut from each archival block onto glass slides by microtome. The first 5 μm section was stained with haematoxylin and eosin (H&E) for histological confirmation. Normal gallbladder tissues from each patient were used to generate the fingerprint pattern of their normal constitutional DNA for comparison.
MICRODISSECTION AND DNA PREPARATION
The remaining 10 μm sections were stained with 0.1% toluidine blue solution for 10 minutes. Tissue samples were separately prepared by microdissection using sterile 27 gauge needles with the assistance of a dissection microscope, and referenced to a section stained with H&E. Deparaffinisation was achieved by incubating in xylene by 20 minutes at 55°C and then washing twice with absolute ethanol. DNA was prepared by incubating tissue in lysis buffer (10 mM Tris.Cl pH 8.0, 50 mM KCl, 2.5 mM MgCl2, 0.1 mg/ml gelatin, 0.45% NP-40, 0.45% Tween 20, proteinase K 500 μg/ml) overnight at 55°C. The lysate was subjected to three phenol/chloroform/isoamyl alcohol (25:24:1, by volume) extractions. After ethanol precipitating, washing, and drying the DNA, it was resuspended in sterile water and the concentration was determined by spectrophotometry.
OPTIMISATION AND VALIDATION OF AP-PCR ON PARAFFIN WAX EMBEDDED MATERIAL
We investigated the parameters that affected the production of reproducible AP-PCR fingerprints. The number, reproducibility, and intensity of bands in a fingerprint should be a function of several parameters, therefore various concentrations of template (50–400 ng), Mg2+ (1.5–7.5 mM), K+ (250–1000 mM), and pH (7.5–10.0) have been investigated. To compare the effect of tissue fixation and processing, normal non-cirrhotic liver tissue was divided into two pieces; the first was snap frozen in liquid nitrogen, while the other was fixed in formalin, embedded in paraffin wax, and processed using standard techniques. A 5 μm section was serially cut from each block onto glass slides and stained with H&E. Tissues from both slides were microdissected from different depths through the block (to test the effects of different lengths of exposure to formalin fixation) and separately collected in microfuge tubes. DNA extraction was performed as above.
AP-PCR
Two different arbitrarily primers (AR316: 5′-GCGAATTCATGTACGTCAGG-3′; ZF317: 5′-CCCCACCGGAGAGAAACC-3′) were utilised to amplify the genomic DNA of each sample. Five μmol of an arbitrary primer were labelled with γ-33P (Amersham, UK) using T4 polynucleotide kinase (Bioline, UK) at 37°C for one hour. AP-PCR amplifications were carried out using 200 ng template DNA in a reaction mixture containing 5 μM of 33P-labelled arbitrary primer, 0.2 mM each dNTP (Bioline, UK), 10 mM Tris.Cl pH 9.2, 3.5 mM MgCl2, 75 mM KCl, and 0.5 units of Taq DNA polymerase (Bioline, UK) in a final volume of 25 μl. The reactions were overlaid with mineral oil and cycled through the following temperature profile: 94°C for one minute to denature, 45°C for five minutes for low stringency annealing of primer, and 72°C for five minutes for extension for four cycles. This temperature profile was followed by high stringency cycles: 94°C for one minute, 60°C for one minute, and 72°C for two minutes for 35 cycles, with a final extension at 72°C for 10 minutes. To ensure reproducibility, all amplifications were performed in duplicate on each sample with a Hybaid Thermoreactor (Hybaid). The PCR product was added to 5 μl of dye mixture and subsequently run on 8% polyacrylamide gel electrophoresis in a Model S2 Sequencing Gel Electrophoresis Apparatus (GibcoBRL Life Technologies, UK). After electrophoresis, the gel was transferred to a piece of Whatman 3MM paper, dried using a gel dryer (Flowgen, UK), and exposed to x ray film (BioMax MR, Kodak, USA) for 24–48 hours at room temperature.
Results
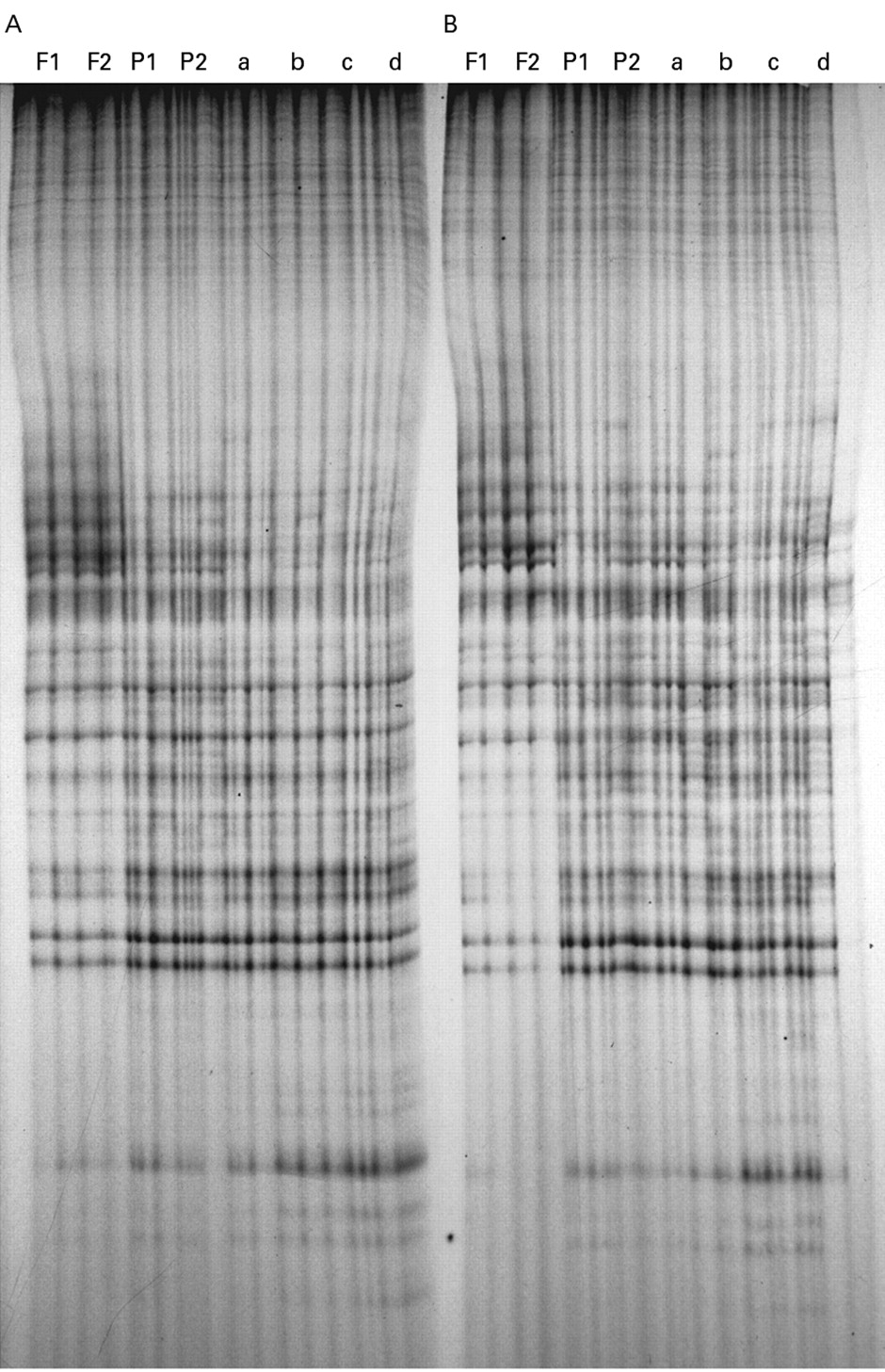
When run on a polyacrylamide gel, the fingerprint patterns generated by AP-PCR from a particular template with either the AR3 or ZF3 primer were consistent over a wide range of template concentrations. Moreover, they were consistent for Mg2+, K+, and pH at 3.5–7.5 mM, 750–1000 mM, and 7.5–9.2 respectively (data not shown). We found identical fingerprinting patterns in AP-PCR reactions of DNA extracted from paired frozen and formalin fixed tissues from an individual sample (fig 1). Similar results were observed in the tissues dissected from different depths.

AP-PCR fingerprint patterns of fresh frozen and formalin fixed, paraffin wax embedded blocks of two segments (A and B) of non-cirrhotic liver from a single patient. Identical profiles were observed on amplification of DNA from microdissected samples at different levels of the frozen blocks (F1 and F2) and paraffin wax blocks (P1 and P2). Similar results were obtained on amplification of various concentrations (a: 50 ng; b: 100 ng; c: 200 ng; d: 400 ng) of template DNA from each of the paraffin wax blocks.
Fifty four HCCs, 31 regenerative nodules (RNs), adjacent benign cirrhotic nodules from 13 cirrhotic explant livers removed at orthotopic liver transplantation, and normal gallbladder tissues were microdissected from the sections of each sample. The mean size of the HCCs was 7.69 (5.56) mm and of the RNs 7.87 (3.39) mm in diameter.
Using AP-PCR with each of the two arbitrary primers, we generated specific and reproducible DNA fingerprints, which were highly polymorphic between different nodules. Each PCR reaction was performed in duplicate with equal aliquots of sample template DNA, and the fingerprint of these paired reactions was identical, confirming the reproducibility of the technique. Genomic homogeneity was detected in the two different sectors (whether closely adjacent or widely separated) of normal tissues of each patient, confirming the specificity of the technique. Nevertheless, DNA fingerprinting of RNs and HCCs was different from that of normal gallbladder tissue in the same patient. Notable genomic heterogeneity was recorded among the nodules studied (fig 2). No two nodules (either RNs or HCCs) had identical electrophoretic patterns. Contrary to expectation, even “satellite” nodules in close proximity within the same segment of the liver were found to have distinct genomic patterns (fig 3).
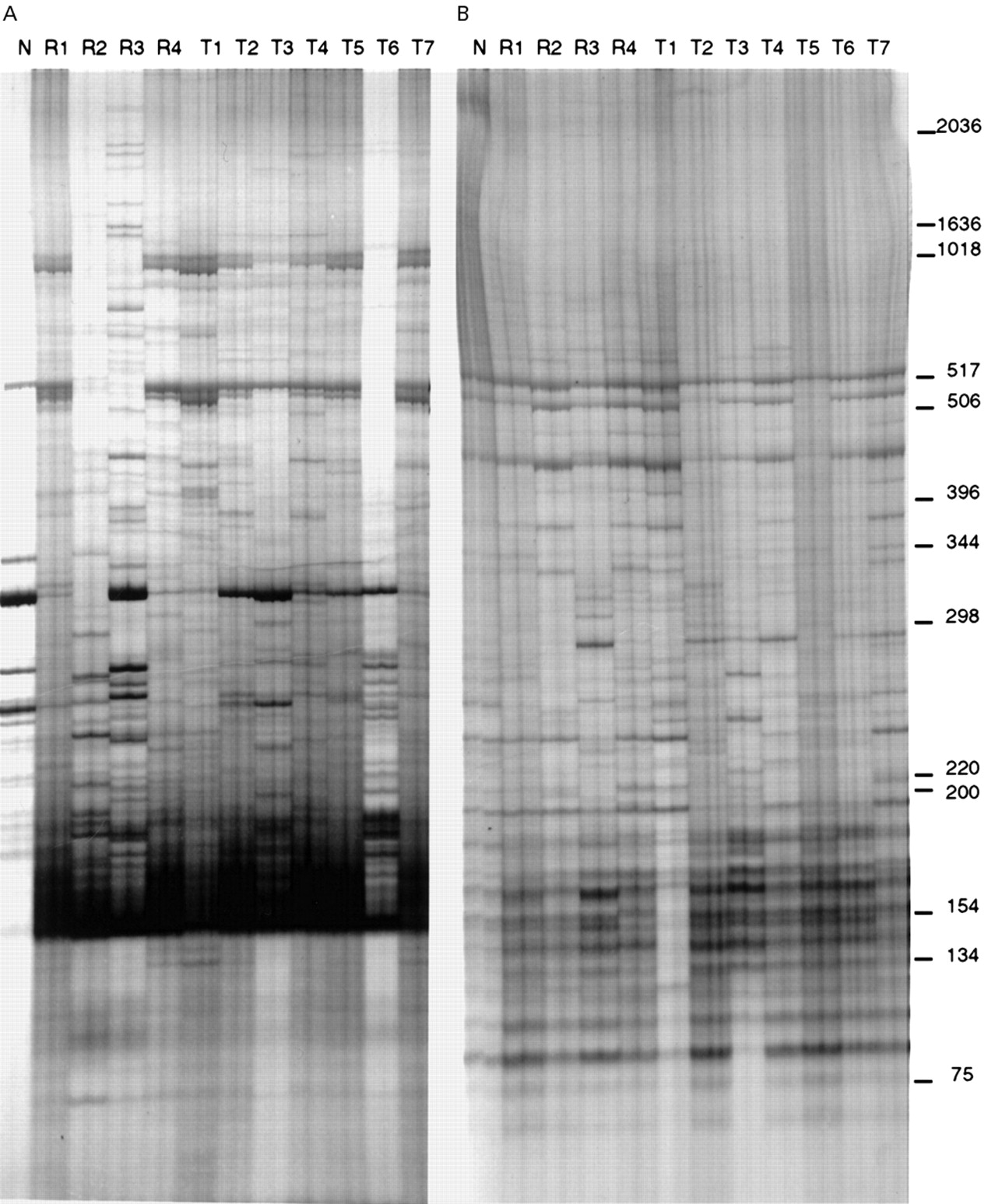
Typical genomic AP-PCR fingerprinting of synchronous hepatocellular carcinoma (HCC) and regenerative nodules (RNs) in the cirrhotic liver of one patient. Autoradiograms of polyacrylamide gel electrophoresis of 33P-labelled DNA fragments amplified by arbitrary primers, AR3 (A) and ZF3 (B), showed high polymorphism between different nodules. N, R1–R3, and T1–T7 represent paired PCR reaction samples from non-tumour tissue, three different RNs, and seven different HCC nodules, respectively. Size of bands (in base pairs) is indicated on the right.

{kind=link}
{kind=link}
{kind=link}
AP-PCR analysis of primary and satellite hepatocellular carcinoma (HCC) nodules. Autoradiograms of polyacrylamide gel electrophoresis of 33P-labelled DNA fragments amplified by AP-PCR. Genomic DNA from each sample was amplified with the AR3 (A) and ZF3 (B) arbitrary primers. S1, S2, and S3 represent the three different sectors in the same primary HCC nodule of patient Sc. N, 1–3, and A–E represent non-tumour tissue, three different sectors of primary, and five satellite HCC lesions of patient Go, respectively. Size of bands (in base pairs) is indicated on the right.
Discussion
AP-PCR is a highly specific technique that is able to generate species specific and individual diagnostic DNA fingerprints.14 The low stringency of the early cycles ensures the generation of products by allowing priming with mismatches between primers and many sequences in the total genomic DNA. The subsequent PCR cycles are performed at a higher stringency after the generation of some initial products that now have ends complementary to the primers. Competition between the annealing events during the initial low stringency cycles results in the reproducible and quantitative amplification of many discrete bands during the subsequent high stringency cycles. Different template genomic DNA sequences display electrophoretic banding patterns in their AP-PCR fingerprints with a number of differences that is inversely proportional to the extent of similarity of the two genomes. This technique has not been previously used to assess the genomic relation between synchronous HCC nodules. We found that AP-PCR is a simple and effective approach for analysing the genetic profile in multiple HCC nodules. Polymorphisms in genomic fingerprints generated by AP-PCR can distinguish between each nodule. Moreover, we successfully showed reproducible and interpretable fingerprinting patterns of amplified genomic DNA isolated from paraffin wax embedded tissues.
The determination of clonality of human tumours has important implications for the mechanisms of carcinogenesis. Resected liver specimens sometimes show small nodular lesions without definite features of cancer in addition to the main HCC. Unfortunately, these lesions are not clearly defined, nor is their nomenclature established. They have been variously referred to as adenomatous hyperplasia, nodular hyperplasia, adenomatous hyperplastic nodules, and macroregenerative nodules (MRNs). In view of such a confused definition of these nodules, we used the term regenerative/macroregenerative nodules (RNs/MRNs) according to the proposals of Ferrellet al.18 Previous studies showed that the presence of MRNs was significantly associated with an increased incidence of liver cell dysplasia and HCC compared with cirrhotic livers without MRNs.19 ,20 It has been suggested that MRNs are possible precancerous lesions. Our findings showed a distinctly different molecular profile among all RNs examined, whereas previous reports of the incidence of monoclonal RNs (based on HBV or HCV analysis) varied from 0.5% to 43%.21-23 In this study, the DNA fingerprints of RNs were different from those of HCCs, even in the same patient, which suggests that HCC may develop independently of RN, by bypassing a rather persistent stage of precancerous nodules. Favouring this view, a previous study showed that no MRNs in 17 patients without treatment developed into HCC in 13–51 months of follow up; indeed, the nodules disappeared in four cases. However, HCCs developed apart from the MRNs in four of these patients during this period.24 There is also an experimental model of de novo occurrence of HCCs in animal models without any precancerous nodular formation.25
Although most HCCs were once believed to be monoclonal in origin,5 ,23 recent studies have shown that a certain proportion of HCCs clearly involve multiclonal development.8 ,9 ,21 ,26 A recent study also showed heterogeneity of DNA content in multiple synchronous HCCs.27 With the background of an anomalous hepatic parenchyma, multiple HCCs can arise independently from different foci in the liver. The genetic heterogeneity in synchronous HCCs may be the result of (in)activation of specific gene loci in the neoplasm, as alternative consequences to the continued presence of carcinogen. Such genomic heterogeneity in synchronous HCCs may explain their poor response to treatment. The high recurrence rate is still a major problem after curative resection of HCC, even for small tumours. If the smaller tumours are de novo lesions rather than metastases (as these data suggest), current concepts regarding liver resection as a curative treatment modality for HCC may require reassessment.
However, this information is based on the explant livers from patients with cirrhosis. Whether the same phenomenon is present in HCCs at an earlier stage of liver disease has not been established. The advent of novel techniques for laser microdissection of single cells may facilitate such analysis.28
Abbreviations used in this paper
- AP-PCR
- arbitrarily primed polymerase chain reaction
- HBV
- hepatitis B virus
- HCC
- hepatocellular carcinoma
- MRN
- macroregenerative nodule
- RN
- regenerative nodule