Article Text

Abstract
BACKGROUND Transforming growth factor β (TGF-β) regulates hepatocyte proliferation and biosynthesis of the extracellular matrix.
AIMS This study investigated alternations in sensitivity to TGF-β1 and binding properties for ligand in hepatocytes and hepatic stellate cells (HSC) after CCl4 administration.
METHODS Plasma TGF-β1 levels in rats after CCl4 administration were determined using ELISA. Effects of TGF-β1 were examined by DNA synthesis in hepatocytes and by measurement of fibronectin production in HSC after CCl4 administration. Binding of125I TGF-β1 was tested in these cells.
RESULTS Plasma TGF-β1 levels were increased as early as 24 hours and were maximal by 48 hours . The antiproliferative response to TGF-β1 decreased in hepatocytes at 48 hours and normalised at 72 hours. Fibronectin production of both normal and injured HSC was affected by TGF-β1 treatment. Cross linked ligand/receptor complexes were detected in normal hepatocytes and HSC. However, these levels decreased specifically in hepatocytes at 48 hours and normalised by 72 hours.
CONCLUSIONS Downregulation of TGF-β receptor occurred in hepatocytes after chemical insult and TGF-β1 could not transduce its antiproliferative signal. Recovery of TGF-β receptor expression causes the signal to transduce to the nucleus at 72 hours. In HSC, whenever TGF-β1 is increased, TGF-β1 can transduce its signal for fibronectin production via its receptor because signalling receptors are expressed constantly.
- TGF-β receptor
- liver regeneration
- fibronectin
- hepatocyte
- hepatic stellate cell
Statistics from Altmetric.com
The transforming growth factor β (TGF-β) proteins are a family of multifunctional cytokines that regulate many aspects of cellular function and consequently have diverse effects on a variety of cell types and tissues. Although five highly homologous types of TGF-β have been characterised, only the first three isoforms are found in mammals.1 2
TGF-β elicit their effects by binding to cell surface receptors. Three major types of TGF-β binding proteins are known to be widely distributed in most TGF-β responsive cells. They are referred to as TGF-β type I (TβRI), type II (TβRII), and type III (or β-glycan) (TβRIII) receptors. TβRI and TβRII are glycoproteins of 53 and 75 kDa, respectively, whereas TβRIII is a proteoglycan of 280– 330 kDa.3-5 TβRI and TβRII possess an extracellular region, a single transmembrane portion, and a serine/threonine kinase domain in the cytoplasmic region. TβRII can bind free ligand whereas TβRI can only recognise ligand that is already bound with TβRII, and formation of a ligand induced heterotetramer involving both TβRI and TβRII is required for signalling. In contrast, TβRIII lacks a cytoplasmic protein kinase domain and appears to function mainly in the concentration and presentation of TGFβ to TβRI and TβRII.6 Signalling by these receptors is mediated by the recently identified Smad protein family.7
TGF-β1 appears to be an important regulator in both normal and pathological conditions in the liver.8 TGF-β1 is synthesised in non-parenchymal cells such as hepatic stellate cells (HSC)9 and inhibits hepatocellular DNA synthesis both in culture and in vivo. Picomolar concentrations of TGF-β1 suppress hepatocyte DNA synthesis in culture.10 Moreover, injection of TGF-β1 into partially hepatectomised rats significantly delays the onset of DNA synthesis.11 In addition, TGF-β1 increases the synthesis and deposition of extracellular matrix (ECM) proteins such as fibronectin by HSC and is closely associated with the progression of hepatic fibrosis.8 12
The level of DNA synthesis in parenchymal hepatocytes and mRNA expression of fibronectin and TGF-β1 in non-parenchymal cells are maximal approximately 48 hours after acute CCl4intoxication.9 13-15 However, these results raise the question of why maximal levels of DNA synthesis are seen when the level of TGF-β1, which inhibits hepatocyte proliferation, is already elevated and ECM is produced by non-parenchymal cells such as HSC. To answer this question, in a previous report we analysed expression of TGF-β and its receptor mRNAs in both hepatocytes and non-parenchymal cells after CCl4 administration.15 The results showed that there is downregulation of TGF-β receptor mRNAs in hepatocytes but that levels remain high in non-parenchymal cells after CCl4 administration. However, it is currently unclear if the cell type specific changes in mRNA expression reflect the abilities of TGF-β receptors to bind ligand and TGF-β signal. Therefore, in this study we focused our efforts on alternations in the sensitivity to TGF-β1 and binding properties for ligand in both hepatocytes and HSC after CCl4 administration.
Materials and methods
ANIMALS
Eight to nine week old, 200–300 g male Wistar rats were obtained from Oriental Bio Service Co. (Kyoto, Japan) and were used in all experiments. All procedures were carried out in accordance with the Declaration of Helsinki and Guiding Principles on the Care and Use of Animals. For studies of liver regeneration after toxic injury, rats were given a single intragastric dose of 0.5 ml of a 1:1 (v/v) mixture of CCl4 in olive oil per 100 g body weight. These animals were sacrificed at 0, 24, 48, 72, and 96 hours after administration of the agent. Blood was collected into heparinised syringes at sacrifice, and plasma samples were stored at −80°C until use.
MEASUREMENT OF TGF-β1 BY ELISA
To determine the concentration of TGF-β1 in rat plasma after CCl4 administration, we used an enzyme-linked immunoassay kit for human TGF-β1 (R&D Systems Inc., Minneapolis, Minnesota, USA) that cross reacts with the rat protein. Values are expressed in ng/ml with a lower limit of detection of 0.005 ng/ml.
CELL SEPARATION PROCEDURE
Hepatocytes and HSC were isolated from normal rat liver or rat liver injured at 48 and 72 hours after CCl4 administration. HSC were obtained using collagenase and pronase-E digestion according to the modified method of Knook and colleagues.16 The liver was perfused in a non-circulating system for two minutes at 37°C (40 ml/min). The perfusion solution was calcium free Hanks buffer (137 mM NaCl, 5.4 mM KCl, 0.5 mM NaH2PO4, 0.42 mM Na2HPO4, 5 mM glucose, 25 mM NaHCO3, 10 mM HEPES, and 0.5 mM EGTA at pH 7.2–7.3). The livers were then circulated with the perfusion solution for five minutes. A solution (200 ml) consisting of 0.1% collagenase (Wako, Osaka, Japan) in 10 mM HEPES, pH 7.5, with 5 mM calcium without EGTA was recirculated for 5–10 minutes. The liver was cut into pieces and incubated in 100 ml of Gey's balanced salt solution with 0.1% pronase-E (Sigma, St Louis, Missouri, USA) on a magnetic stirrer at 37°C for 30 minutes. At the end of the incubation period, the suspension was filtered through Nylon gauze and the filtrate was centrifuged at 450 g for seven minutes. The collected cells were centrifuged on a two layer density cushion of Nycodenz (Nycomed, Oslo, Norway) (Gey's balanced salt solution/8% Nycodenz) at 1400 g for 17 minutes. The HSC fraction was collected from the upper layer. The purity of the HSC was greater than 90%, as assessed by typical morphological features and the immunological staining of desmin. Hepatocytes were prepared by perfusion of the liver in situ with collagenase and purified by centrifugal elutriation as previously described.15
LIVER CELL CULTURE
Purified cells were kept in culture at 37°C in a 5% CO2 atmosphere and 100% humidity. Hepatocytes were cultured in Williams' medium E supplemented with 5% fetal calf serum (FCS), 10 nM dexamethazone, 100 units/ml penicillin G, and 0.1 mg/ml streptomycin. HSC were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 5% FCS, 100 units/ml penicillin G, and 0.1 mg/ml streptomycin.
ASSAY OF DNA SYNTHESIS
The isolated hepatocytes were suspended in culture medium at 1×105 cells/ml and seeded into 24 well dishes (0.5 ml/dish). After two hours the medium was changed to serum free Williams' medium E. Various concentrations of porcine TGF-β1 (R&D Systems Inc.) with or without 10 ng/ml of recombinant human hepatocyte growth factor (HGF) (R&D Systems Inc.) were added to the medium 12 hours after seeding. After further incubation for 24 hours, 2 μCi/well of [methyl-3H]thymidine (25 Ci/mmol; Amersham, Buckinghamshire, UK) were added to the medium and culture was continued for three hours. At the end of the pulsing period, the medium was carefully aspirated, icecold 5% trichloroacetic acid was added, and dishes were kept on ice for 15 minutes. After two additional washes with 5% trichloroacetic acid, cells were solubilised by adding 500 μl of 0.5 N NaOH. The solubilised cell solution (250 μl) was then neutralised with 0.5 N HCl (250 μl) and radioactivity (cpm) measured in a liquid scintillation counter.
MEASUREMENT OF FIBRONECTIN PRODUCTION
Fibronectin production was measured as described, with minor modifications.17 The isolated HSC were suspended in culture medium at 2×105 cells/ml and seeded into 12 well dishes. After two hours the medium was changed to fresh culture medium containing 0.2% FCS. After five hours the cells were incubated with or without various concentrations of TGF-β1 for 20 hours and labelled with 50 μCi of a [35S]methionine and [35S]cysteine mixture (Pro-mix cell labelling mix: Amersham) in 1 ml of methionine and cysteine free DMEM (ICN Pharmaceuticals, Costa Mesa, California, USA) for the final four hours. Labelled culture media (1 ml) were collected and labelled fibronectin was purified by absorption to 100 μl of gelatin-sepharose (Pharmacia, Uppsala, Sweden) in the presence of 0.5% Triton X-100. The beads were washed once in Tris buffered saline (50 mM Tris HCl, pH 7.4, 150 mM NaCl), once in 50 mM Tris HCl (pH 7.4), 0.5 M NaCl, and once in Tris buffered saline. Fibronectin was eluted by boiling in sodium dodecyl sulphate (SDS) sample buffer in the presence of 10 mM dithiothreitol. The samples were analysed by 7% SDS-polyacrylamide gel electrophoresis (PAGE) and autoradiography. Relative amounts of fibronectin were quantified by densitometric scanning.
BINDING AND COVALENT AFFINITY CROSS LINKING OF LIGAND
Carrier free TGF-β1 was iodinated to a specific activity of 1×105 cpm/ng (25 μCi/ng) by the chloramine T method as previously described.18 The isolated hepatocytes and HSC were suspended in culture medium at 3×106 cells/ml and seeded onto 10 cm dishes. After three hours, cell monolayers were incubated for one hour at 4°C with 200 pM 125I labelled TGF-β1 alone or with excess (5 nM) unlabelled TGF-β1. Receptors were cross linked to bound ligand with disuccinimidyl suberate and solubilised in the presence of Triton X-100 as previously described.19 Cell extracts were clarified by centrifugation and then subjected to 7% SDS-PAGE and autoradiography.
STATISTICAL ANALYSIS
Data are expressed as mean (SD). Experimental and control values were compared using the unpaired Student'st test and ANOVA; p<0.05 was considered significant.
Results
ELEVATION OF TGF-β1 LEVEL IN RAT PLASMA AFTER CCL4 ADMINISTRATION
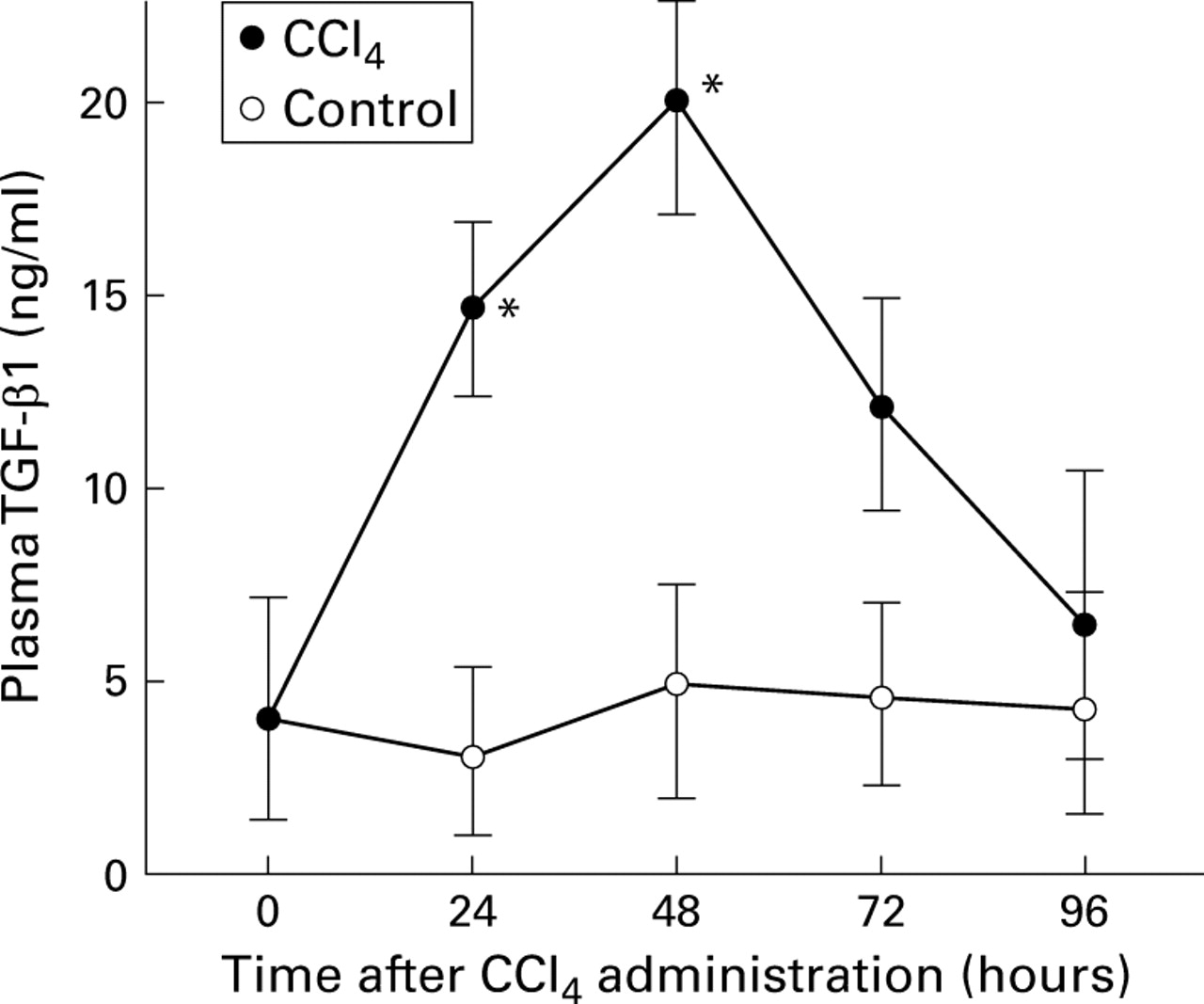
Using northern blot hybridisation we observed approximately fivefold induction of TGF-β1 mRNA in regenerating livers at 48 hours after CCl4 administration.9 15 To determine if protein levels also increased we measured TGF-β1 protein by ELISA in rat plasma after CCl4 administration. Figure 1illustrates the results of plasma TGF-β1 measurements after CCl4 administration. Plasma levels of TGF-β1 were increased as early as 24 hours and were maximal by 48 hours, reaching five times that in normal plasma and decreasing thereafter. Plasma levels of TGF-β1 were essentially unchanged in the rat treated only with olive oil at various times between 24 and 96 hours. Consistent with mRNA induction, TGF-β1 protein was induced in the injured liver. These results suggest that elevated levels of TGF-β1 have potential roles in liver regeneration and ECM production. Therefore, we studied the time dependent differential sensitivity of individual liver cell types to the growth inhibitory effect of TGF-β1 and its induction of ECM expression.
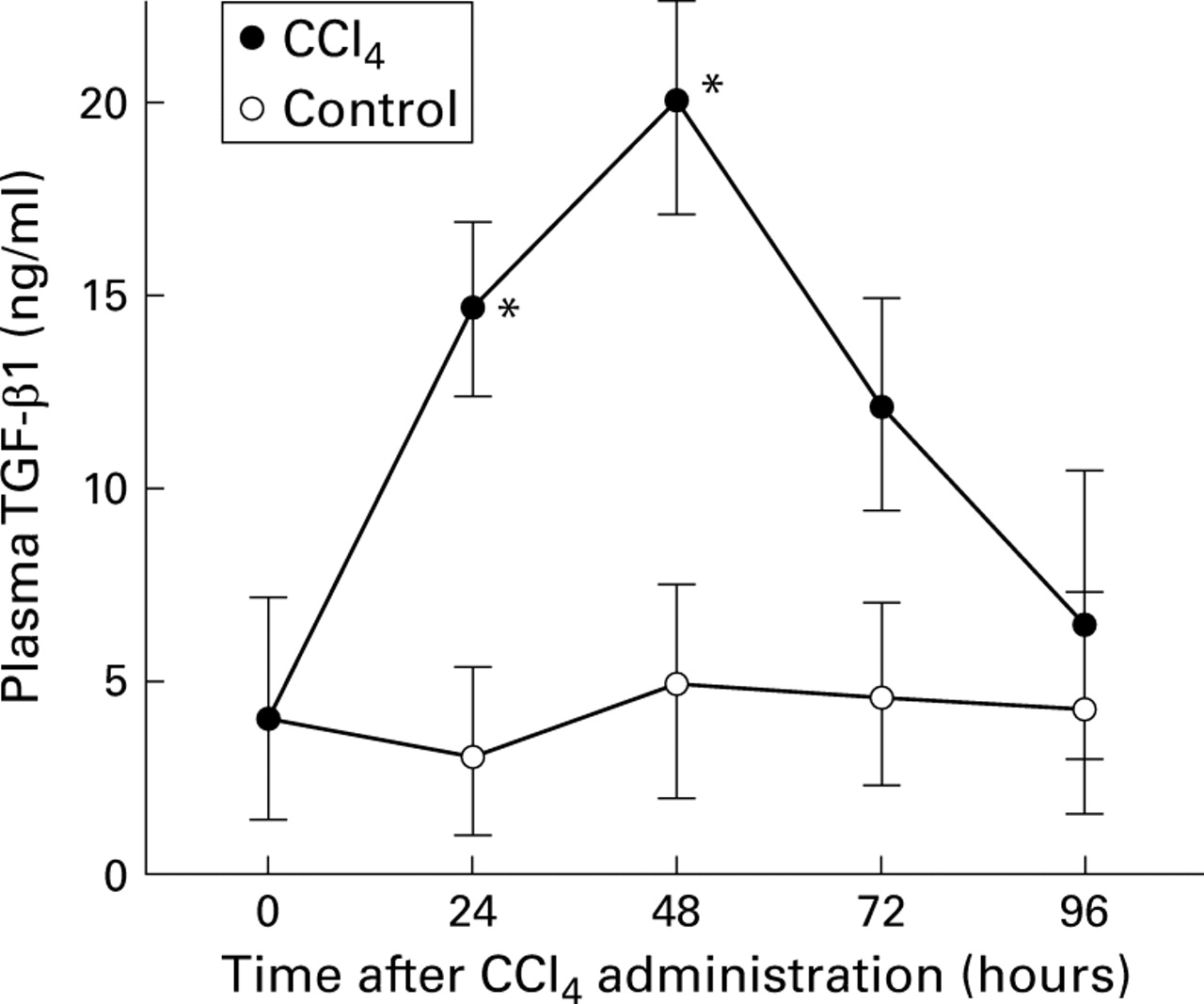
Quantitative analysis of plasma transforming growth factor β1 (TGF-β1) protein after CCl4 administration by enzyme linked immunoassay. Plasma samples were collected from normal and injured rats at the indicated times after CCl4administration and stored at −80°C. Data are expressed as mean (SD) for triplicate determinations in CCl4 treated and control rats. *p<0.05 v control.
DIFFERENTIAL ANTIPROLIFERATIVE RESPONSE OF TGF-β1 ON HEPATOCYTES AFTER CCL4 ADMINISTRATION
First we examined if TGF-β1 induced by liver injury affected hepatocyte growth. The effect of TGF-β1 on cellular proliferation was studied by measuring [methyl-3H] thymidine incorporation into the DNA of hepatocytes. Figure 2 shows the effects of TGF-β1 on DNA synthesis of normal hepatocytes in the presence or absence of exogenous HGF. HGF induced a fivefold increase in DNA synthesis of hepatocytes. TGF-β1 inhibited DNA synthesis of hepatocytes not only in the absence but also in the presence of exogenous HGF in a dose dependent manner in the range 1–104 pg/ml, consistent with previous reports.20 This effect was half maximal with approximately 10 pg/ml TGF-β1.

Effect of transforming growth factor β1 (TGF-β1) on DNA synthesis of cultured hepatocytes. Experimental conditions were as described in the text. Various concentrations of TGF-β1 were added as indicated in the absence or presence of exogenous hepatocyte growth factor (HGF) (10 ng/ml). Data are expressed as mean (SD) of triplicate determinations. *p<0.05 v control.
It is not clear why maximal levels of DNA synthesis are observed at 48 hours in spite of the increased levels of TGF-β1 which inhibit hepatocyte proliferation. To determine this we monitored the sensitivity of hepatocytes to the growth inhibitory effect of TGF-β1 after CCl4 administration and compared DNA synthesis of normal liver in the presence of TGF-β1 with that of injured liver. In the presence of 10 pg/ml TGF-β1, HGF induced DNA synthesis in normal hepatocytes was reduced to approximately 50%. This inhibitory effect of TGF-β1 was decreased to approximately 20% in hepatocytes at 48 hours after CCl4 administration and returned to normal levels at 72 hours (fig 3). These results indicated that hepatocytes at 48 hours after chemical insults showed a reduced response to growth inhibition by TGF-β1.

Time dependent differential sensitivity of hepatocytes to the growth inhibitory effect of transforming growth factor β1 (TGF-β1) after CCl4 administration. Hepatocytes were isolated from normal rat liver or rat liver injured at 48 and 72 hours after CCl4 administration. The hepatocytes were subjected to a [3 H] thymidine assay in the presence of 10 pg/ml of TGF-β1 and 10 ng/ml of hepatocyte growth factor (HGF). The percentage of growth inhibition was calculated by measuring the decrease relative to[3H] thymidine incorporation into cells in the absence of TGF-β1. Data are expressed as mean (SD) for triplicate determinations. *p<0.05 v control.
STIMULATION OF FIBRONECTIN PRODUCTION BY TGF-β1 IN HSC AFTER CCl4ADMINISTRATION
At the regenerating stage of the liver, hepatocytes seemed to be resistant to TGF-β1 stimuli although the protein level of TGF-β1 was already increased. However, this leads to the question of whether the elevated TGF-β1 has other biological effects on non-parenchymal cells. In addition, TGF-β1 increased synthesis and deposition of ECM proteins in many mesenchymal cells.1 2 Therefore, we next examined ECM production from non-parenchymal cells. We used HSC because of the central importance for ECM production and liver fibrosis.21 Furthermore, we examined fibronectin production because fibronectins are major components in the liver and representative of the various matrix proteins induced by TGF-β1.22
To examine if TGF-β1 induced the production of fibronectin in hepatocytes and HSC, a fibronectin assay was performed by measuring incorporation of [35S]methionine and [35S]cysteine into fibronectin. The production of fibronectin in HSC, but not in hepatocytes, was induced dose dependently by TGF-β1 (fig 4A). The resulting changes in the expression of fibronectin shown in fig 4A are graphically represented in fig 4B. This effect in HSC was half maximal with approximately 100 pg/ml TGF-β1.

Fibronectin production induced by transforming growth factor β1 (TGF-β1) in hepatocytes (Hepa) and hepatic stellate cells (HSC). Cells were incubated with culture medium for five hours. Cells were then incubated with various concentrations of TGF-β1 for 20 hours and labelled with a [35S]methionine and [35S]cysteine mixture for the last four hours. Fibronectin secreted into media was purified by adsorption to gelatin-Sepharose, analysed by 7% SDS-PAGE, and visualised by autoradiography. This figure is representative of three separate experiments (A). Changes in expression of fibronectin were quantitated by densitometry (B). The values shown are the mean of results from three separate experiments. Fibronectin expression in untreated cells was assigned a value of 1 and other values expressed relative to this were plotted against the concentration of exogenous TGF-β1. The horizontal broken line indicates the control value.
The sensitivity of HSC to TGF-β1 at 48 hours after chemical insult needs to be studied because hepatocyte growth is resistant to TGF-β1 at this time. Therefore, we compared fibronectin production of normal HSC in the presence of TGF-β1 with that of HSC after CCl4administration (fig 5). The level of fibronectin was already elevated in activated HSC after CCl4 administration. However, similar to normal HSC, 100 pg/ml TGF-β1 stimulated fibronectin production in HSC at 48 hours when hepatocytes, after chemical insult, showed a reduced response to growth inhibition by TGF-β1. This indicated that fibronectin production of HSC after CCl4administration was also affected by TGF-β1 treatment. The resulting changes in the expression of fibronectin shown in fig 5A are graphically represented in fig 5B.

Time dependent differential sensitivity of hepatic stellate cells (HSC) to the effect produced by transforming growth factor β1 (TGF-β1) after CCl4 administration. HSC were isolated from normal rat liver or rat liver injured at 48 and 72 hours after CCl4 administration. HSC were subjected to fibronectin assay in the absence or presence of 100 pg/ml of TGF-β1. Fibronectin secreted into the media was purified by adsorption to gelatin-sepharose, analysed by 7% SDS-PAGE, and visualised by autoradiography. This figure is representative of three separate experiments (A). Changes in expression of fibronectin were quantitated by densitometry (B). For each culture condition the relative expression in untreated normal HSC was assigned a value of 1. Data are expressed as mean (SD) for triplicate determinations. *p<0.05 v control.
DIFFERNTIAL BINDING OF 125I TGF-β1 TO THE RECEPTORS ON HEPATOCYTES AND HSC AFTER CCl4ADMINISTRATION
In contrast with the small antiproliferative response to TGF-β1 on hepatocytes during liver regeneration, the sensitivity of HSC for fibronectin production by TGF-β1 was not affected after chemical insult. To pursue the possible mechanisms of these differences, we focused our efforts on the affinities of the ligand to the receptors in both hepatocytes and HSC after CCl4 administration. To test TGF-β receptor for the ability to bind ligand in hepatocytes (fig 6A) and HSC (fig 6B) after CCl4 administration,125I TGF-β1 was cross linked to these cells using the homobifunctional cross linker disuccinimidyl suberate. 125I TGF-β1 bound to three different proteins in normal HSC with molecular weights of approximately 65, 85, and 300 kDa, which disappeared on addition of an excess of unlabelled TGF-β1. From the molecular weights, the three proteins appear to represent 125I TGF-β1-TβRI, 125I TGF-β1-TβRII, and125I TGF-β1-TβRIII complexes, respectively.125I TGF-β1-TβRI and 125I TGF-β1-TβRII complexes were observed in normal hepatocytes as well as HSC. However,125 I TGF-β1-TβRIII complexes were scarcely detected. At 48 hours after CCl4 administration, TβRI and TβRII in hepatocytes interacted very weakly with ligand. Then, the ability of two TGF-β receptors to bind ligand recovered, reaching normal levels at 72 hours. Our results using a receptor cross linking assay suggested that expression of TβRI and TβRII was depressed specifically in hepatocytes at 48 hours after CCl4 administration. Unlike in hepatocytes, the ability of TβRI and TβRII to bind ligand did not decrease in HSC during treatment.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Time dependent differential binding of 125I transforming growth factor β1 (TGF-β1) for the receptors on hepatocytes (A) and hepatic stellate cells (HSC) (B) after CCl4 administration. Cells were isolated from normal rat liver or rat liver injured at 48 and 72 hours after CCl4administration. Cells were affinity labelled with 200 pM of125I TGF-β1 in the absence or presence of 5 nM unlabelled (cold) TGF-β1. Labelled proteins were separated by SDS-PAGE and visualized by autoradiography. This figure is representative of three separate experiments.
Discussion
In this present study we addressed the important question of the role of TGF-β in regulating cell proliferation and ECM synthesis following rat liver injury. We have reported previously that the levels of TβRI and TβRII mRNA expression in rat hepatocytes decreased from 12 to 48 hours and returned to normal by 72 hours after CCl4 administration while TβRI and TβRII mRNA were expressed constantly in non-parenchymal cells.15 These results were consistent with the change in binding of 125I TGF-β1 to the receptors on hepatocytes and HSC after CCl4administration. At 48 hours after CCl4 administration, TβRI and TβRII in hepatocytes interacted very weakly with ligand. Our results indicated that there is a transient downregulation of TGF-β receptor proteins in the hepatocytes following CCl4administration.
TGF-β is known to be the most potent growth inhibitor for a wide variety of cell types, including hepatocytes.1 10 Our results showed that the inhibitory effect of TGF-β1 decreased in hepatocytes at 48 hours after CCl4 administration and returned to normal level at 72 hours. Consistent with downregulation of the mitoinhibitory effect, DNA synthesis in hepatocytes reached a peak at 48 hours after CCl4 administration. This result raises the possibility that downregulation of the mitoinhibitory effect of TGF-β1 enhances the effect of mitogenic factor induced by chemical insult. During the proliferative phase, there is an inverse correlation between expression of the TGF-β receptor and DNA synthesis in hepatocytes. These observations suggest that by decreasing receptor expression, TGF-β1 cannot transduce its antiproliferative signal via TGF-β receptors on hepatocytes. Furthermore, return of receptor expression to a normal level in the later stages of regeneration may account for the return in hepatocyte sensitivity to TGF-β and may be involved in limiting hepatocyte proliferation once the liver has regenerated.
Although the mechanism of reduction of TGF-β receptor transcripts is currently unknown, several reports analysed the promoter region of TβRII to pursue the transcriptional mechanism of this gene. Baeet al identified two positive regulatory elements and at least one negative regulatory element in the promoter region of TβRII.23 Furthermore, Kimet al reported that an absolute reduction of TβRII mRNA followed by transformation of keratinocytes with E1A oncogene is the result of decreased expression of unidentified transcription factor complexes that interact with two positive regulatory elements.24 Therefore, we speculate that downregulation of TβRII in hepatocytes is caused by a decrease in these transcription factors.
Among the most striking effects of TGF-β on cellular function in vitro is its promotion of extracellular formation. TGF-β increases synthesis and deposition of extracellular components such as fibronectin and some forms of collagen.1 8 In the liver, HSC is an important effector of hepatic fibrosis.12 21Despite depression of TGF-β receptor levels in hepatocytes, these levels remained constant in HSC in the liver injured by CCl4 treatment. In addition, TGF-β1 stimulated the production of fibronectin even by injured HSC. These results suggest that TGF-β1 induces many components of the ECM by an autocrine mechanism whenever TGF-β1 is produced after CCl4administration.
TGF-β1 mRNA expression in non-parenchymal cells reaches a maximum at about 48 hours after CCl4 administration.9 15In addition, we report that plasma levels of TGF-β1 are also increased as early as 24 hours after CCl4 administration and are maximal by 48 hours, returning to control levels after four days. However, the question of why the regulation of hepatocyte proliferation and promotion of ECM deposition occurred at the same time remains unanswered. Our present results confirmed a hypothesis concerning the action of TGF-β1 in liver regeneration and ECM production. In HSC, TGF-β1 can transduce its signal for ECM production via its receptor stably by an autocrine mechanism because signalling receptors are expressed constantly after CCl4administration. In contrast, without the receptor, TGF-β1 cannot bind to hepatocytes and block the DNA replication signal induced by mitogenic factor at 48 hours after CCl4 administration. Recovery of TGF-β receptor expression in hepatocytes makes the antiproliferative signal transduce to the nucleus at 72 hours. Further analysis of TGF-β signal transduction including as the Smad protein family in liver damaged by viruses or chemical toxicity will contribute to our understanding of the mechanism of liver regeneration and ECM production.
Acknowledgments
This study was supported by a grant-in-aid for scientific research from the Ministry of Education, Science, and Culture of Japan.
References
Footnotes
- Abbreviations used in this paper:
- DMEM
- Dulbecco's modified Eagle medium
- ECM
- extracellular matrix
- FCS
- fetal calf serum
- HGF
- hepatocyte growth factor
- HSC
- hepatic stellate cells
- PAGE
- polyacrylamide gel electrophoresis
- SDS
- sodium dodecyl sulphate
- TGF-β
- transforming growth factor β
- TβRI
- TβRII, TβRIII, TGF-β type I, II, and III receptors