Article Text

Abstract
BACKGROUND Prostaglandin endoperoxide synthase/cyclooxygenase (COX) is the key enzyme in gastric mucosal protection and repair but its cellular localisation in the human stomach is still unclear.
AIMS To investigate immunohistochemically the cellular distribution of COX-1 and COX-2 proteins in the human stomach with or without gastritis or ulceration.
PATIENTS AND METHODS Tissues were obtained by surgical resection of gastric ulcers associated with perforation (n=9) or by biopsy from Helicobacter pylori positive patients with gastric ulcers (n=45) and H pylori negative healthy subjects (n=15). COX expression was detected by semiquantitative reverse transcription-polymerase chain reaction (RT-PCR), western blotting, and light and electron microscopic immunohistochemistry.
RESULTS COX-2 mRNA and protein were detected in gastric ulcer tissues but not in intact gastric mucosa. COX-1 mRNA and protein were detected in the intact mucosa. COX-2 immunostaining was exclusively localised in macrophages and fibroblasts between necrotic and granulation tissues of the ulcer bed. The percentage of COX-2 expressing cells was significantly higher in open than in closed ulcers, and in gastritis than in gastric mucosa without H pylori infection. COX-1 immunoreactivity localised in lamina propria mesenchymal cells was similar in various stages of ulcer disease and in intact gastric mucosa. Electron microscopic immunohistochemistry revealed both COX-1 and COX-2 on the luminal surfaces of the endoplasmic reticulum and nuclear envelope of macrophages and fibroblasts.
CONCLUSIONS Our results showed that COX-2 protein was induced in macrophages and fibroblasts in gastric ulcers and H pylorirelated gastritis, suggesting its involvement in the tissue repair process.
- cyclooxygenase
- macrophage
- fibroblast
- gastric ulcer
- gastritis
Statistics from Altmetric.com
Prostaglandins (PGs) are known to protect the mucosal lining of the stomach against injury caused by a variety of necrotising agents.1-3 PG endoperoxide synthase/cyclooxygenase (COX) is one of the rate limiting enzymes of PG synthesis from arachidonic acids. There are two forms of COX, a constitutively produced COX-1 and an inducible COX-2.4-6 COX-1 protein is present in a variety of tissues, including the stomach, whereas COX-2 mRNA and protein are induced in macrophages by lipopolysaccharide,7 8 in fibroblasts by platelet derived growth factor,9 and in epithelial cells by epidermal growth factor (EGF) family peptides.10 11
Recent studies have shown that COX gene expression and enzyme activity are regulated in the gastric mucosa bearing erosions or ulcers.12-15 Furthermore, COX-1, but not COX-2, is expressed in the intact stomach without lesions whereas expression of COX-2 mRNA, but not that of COX-1 mRNA, is increased in gastric mucosal lesions induced experimentally in animals by intragastric administration of acidified ethanol or the ischaemia-reperfusion technique.12 13 15 In this regard, high levels of COX-2 mRNA and protein during the acute stages of gastric lesions as well as during experimentally induced colitis are thought to be involved in the repair process of these lesions.16
To our knowledge localisation of COX-1 protein in human gastric mucosal lesions has not been reported previously. Furthermore, there have been conflicting reports as to the localisation of COX-2 protein in the human stomach.17-19 Previous studies used animal models in which the gastric lesions were induced experimentally but these data may not reflect the actual COX gene and protein expression in the human stomach with gastritis or ulceration. In the present study, we determined expression of COX mRNA and protein in the human stomach, the distribution of COX-1 and COX-2 proteins, and the level of expression of these proteins in human gastric tissues with eitherHelicobacter pylori related gastritis or gastric ulceration. Studies were conducted using either gastric tissues with ulceration or gastric biopsy samples obtained from ulcer margins. These tissues were examined immunohistochemically and by reverse transcription-polymerase chain reaction (RT-PCR) and western blot analysis.
Material and methods
PATIENTS
Gastric tissues from 54 patients with gastric ulcers and 15 healthy individuals were used. In nine patients (seven men and two women; age range 44–79 years; mean 60.0), gastric ulcer tissues with perforations were obtained by surgical resection. None had been treated with H2 receptor antagonists or proton pump inhibitors at the time of resection. In the other 45 patients (30 men and 15 women; age range 24–80 years, mean 61.0) and 15 healthy individuals (10 men and five women; age range 18–66 years, mean 42.3), gastric tissues were obtained by biopsy during endoscopy. Patients receiving non-steroidal anti-inflammatory drugs (NSAIDs) (including low dose aspirin) for medical indications were excluded. All subjects gave informed consent and the project was reviewed and approved by the ethics committee of Nippon Medical School, Tokyo, Japan.
ENDOSCOPIC APPEARANCE OF GASTRIC ULCERS AND DETECTION OFH PYLORI
Endoscopically, the acute stage of gastric ulceration was defined by the presence of marginal oedema with little or no regenerative mucosa, while the healing stage was characterised by the appearance of regenerative mucosa in the surrounding mucosa. The ulcer base of acute and healing stages was covered by an adherent, granular, grayish-white exudate. The presence of H pylori in the biopsy material was determined either by the rapid urease test or histologically using the Giemsa stain, or both. Patients were classified as H pylori positive if at least one of these examinations yielded a positive result.
RT-PCR ANALYSIS OF COX-1 AND COX-2 mRNAs
To compare levels of expression of COX mRNA in the stomach of those infected or not infected with H pylori, we used the semiquantitative RT-PCR method described previously.12 Gastric tissue samples obtained by biopsy were homogenised in a guanidine thiocyanate solution and the total cellular RNA was extracted using the standard guanidine thiocyanate method. In the next step, 2 μg of total RNA were dissolved in a total volume of 25 μl of 10 mmol/l Tris HCl buffer, pH 8.3, containing 500 mmol/l deoxynucleotide triphosphate (dNTP) mixture, 4 nmol random primer, and 8 U of murine leukaemia virus reverse transcriptase (Takara, Kyoto, Japan) and was then reverse transcribed at 42°C. For amplification, PCR was performed exactly as previously described.12 Briefly, 1.0 μl of the resulting cDNA product was incubated in a total volume of 10 μl of 10 mmol/l Tris HCl buffer, pH 8.3, containing 50 mmol/l KCl, 1.5 mmol/l MgCl2, 200 mmol/l dNTP mixture, a primer pair of either COX-1 and COX-2 or β-actin cDNA, and 2.5 U of Taq DNA polymerase (Takara). The thermal profile used in the Perkin-Elmer Cetus thermal cycler consisted of denaturation at 94°C for one minute, annealing at 55°C for both COX-1 and β-actin and at 61°C for COX-2 for 1.5 minutes, and extension at 72°C for two minutes. This was repeated for 30 cycles with a final extension process for three minutes to allow complete synthesis of the amplified product. PCR products were electrophoresed on 1% agarose gel and visualised by staining with ethidium bromide. PCR primers were as follows: COX-1, 5′-GCAACACTG GAACATGGCTA-3′ as a sense primer and 5′-ACGCCACCATTCTGTCTTTG-3′ as an antisense primer; COX-2, 5′-TCATTCACC AGGCAAATTGCTGGCAGGG-3′ as a sense primer and 5′-ACAGTTCAGTCGAACGTT CTTTTAGTAGTAC-3' as an antisense primer; and β-actin, 5′-CAAGAGATGGCCA CGGCTGCT-3′ as a sense primer and 5′-TCCTTCTGCATCCTGT CGGCA-3′ as an antisense primer. These primer pairs were found to yield PCR products of 517 base pairs (bp), 583 bp, and 275 bp for COX-1, COX-2, and β-actin, respectively. Quantitative image analysis of the PCR products on the gel was performed on a Macintosh computer using the public domain National Institutes of Health Image program version 1.61 (developed at the US National Institutes of Health) as described previously.20
CHARACTERISTICS OF ANTIBODIES
Antihuman rabbit polyclonal COX-1 or COX-2 antibody generated against 17 amino acids at position 251–267 was purchased from Immuno-Biological Laboratories Co. (Gunma, Japan). Characteristics and specificities of the anti-COX-2 antibody used in the present study have been described in previous studies.21-24
WESTERN BLOT ANALYSIS
Gastric and duodenal tissue samples obtained by biopsy were homogenised in 25 mmol/l Tris HCl (pH 8.0), 0.25 mol/l sucrose containing 1.0 mmol/l phenylmethylsulphonyl fluoride, 1.0 mmol/l pepstatin A, and 1.0 mmol/l ethylenediaminetetra-acetic acid. CHAPS was added to 1% (wt/vol) and the mixture was stirred for two hours at 4°C. After centrifugation at 15 000 g for 10 minutes, the supernatant was loaded onto an anion exchange column equilibrated with 25 mmol/l Tris HCl (pH 8.0) plus 0.4% CHAPS. The fraction eluted at 500 mmol/l NaCl was concentrated to 40% of the initial volume. Samples containing 0.1 mg of protein were separated on 10% acrylamide gel by sodium dodecyl sulphate-polyacrylamide gel electrophoresis. After electrophoresis, the separated proteins were transferred to a Hybond-P nitrocellulose membrane (Amersham Life Science, Buckinghamshire, UK) and probed with anti-COX-1 or anti-COX-2 antibody (diluted 1:100, IBL). Bound antibodies were detected with horseradish peroxidase conjugated antirabbit IgG (1:2000) using the enhanced chemiluminescence detection system.
IMMUNOHISTOCHEMISTRY
Two biopsy samples were obtained, one from the ulcer edge and the other from the corpus of the stomach. Both specimens were fixed in 10% formalin and embedded in paraffin wax. Surgically resected specimens were treated in a similar fashion. Embedded gastric tissues were cut into 3 μm sections which were then immersed in 0.3% H2O2 in methanol for 30 minutes to block endogenous peroxidase activity. Sections were then microwaved in 0.01 mol/l citrate phosphate buffer (pH 6.0) for antigen retrieval, and incubated with 10% normal goat serum for 10 minutes at 37°C to block non-specific IgG binding. In the next step, sections were incubated for 18 hours at 4°C with rabbit antihuman COX-1 or COX-2 antibodies (diluted 1:25, IBL). They were then treated for one hour at 37°C with biotinylated antirabbit IgG (1:500; Dako a/s, Glostrup, Denmark) followed by treatment with avidin and biotinylated peroxidase complex (Dako) for one hour at room temperature. The reaction products were developed by immersing sections in 3,3′-diaminobenzidine tetrahydrochloride (DAB) solution containing 0.03% H2O2. Nuclei were counterstained with Mayer's haematoxylin. Non-immunised rabbit serum was used as a negative control.
SCORING METHODS
Immunohistochemical findings in biopsy specimens were analysed according to ulcer stage. We prepared two sections from each biopsy sample to examine COX-1 and COX-2 positivity. Almost all specimens contained the surface epithelium and muscularis mucosa. When COX-1 immunoreactivity was observed in both glandular and mesenchymal cells, patients were classified as positive for COX-1. In contrast, patients were classified as positive for COX-2 when any mesenchymal cells showed perinuclear and cytoplasmic COX-2 immunoreactivity. Negative cases consisted of sections lacking immunostaining in mesenchymal cells. All sections were examined independently by an investigator who was blind to the pathological and clinical data.
PRE-EMBEDDING ELECTRON MICROSCOPIC IMMUNOHISTOCHEMISTRY
In a few cases gastric biopsy samples were fixed in 4% paraformaldehyde for six hours for electron immunohistochemistry. After immersion of tissues in graded concentrations of sucrose (10% for four hours, 20% for four hours, 30% for four hours) at 4°C, they were embedded in OCT compound and frozen rapidly with dry ice/acetone. Sections (6 μm thick) were treated with 10% normal swine serum for 30 minutes at room temperature and incubated for 18 hours at 4°C with the COX antibodies described above (diluted 1:20). They were then incubated with peroxidase conjugated swine antirabbit IgG (diluted 1:50; Dako) for 18 hours at room temperature. After washing, tissue sections were treated with 1% glutaraldehyde for five minutes at room temperature, and sequentially treated with DAB solution for 30 minutes and DAB-H2O2 solution for five minutes. After fixation with 2% osmic acid for two hours sections were washed, dehydrated, and embedded in Epok 812 (Nagase Co., Tokyo). Ultrathin sections were lightly counterstained with lead citrate and examined under a Hitachi H-7100 electron microscope (Tokyo).
STATISTICAL ANALYSIS
Statistical analysis was performed using Statview software package (Abacus Concept Inc, California, USA). Significant differences between the two groups were analysed using the χ2 for 2×2 tables with Fisher's exact test when necessary. When multiple comparisons were performed, data were analysed by one way analysis of variance followed by Fisher's protected least significant difference (PLSD) test. A p value less than 0.05 indicated a statistically significant difference.
Results
COX mRNA AND PROTEIN LEVELS IN HUMAN STOMACH
We initially determined the level of expression of COX mRNA in human gastric mucosa with or without ulcers by semiquantitative RT-PCR. We prepared RNA and subsequent cDNA using biopsy samples from the gastric ulcer margin (n=4), gastritis (n=4), and intact gastric mucosa without H pylori infection (n=4). COX-1 and β-actin cDNA fragments were detected in all samples from gastric tissues (fig 1). In contrast, COX-2 cDNA fragment was detected in tissues from H pylori infected patients (n=4) but was absent in intact mucosa without H pylori infection (n=4). To further analyse levels of expression of COX mRNA against β-actin, we performed gel densitometry of COX-1 and COX-2 cDNA products standardised against constitutively expressed β-actin (fig 1B and C, respectively). There was no significant difference in COX-1 expression in gastric tissues with or withoutH pylori infection. In contrast, expression of COX-2 mRNA was significantly higher in both gastritis and gastric ulcer tissues with H pylori infection than in H pylori negative intact gastric mucosa (p<0.005; Fisher's PLSD test).

COX and β-actin mRNA levels in gastric biopsy samples. COX and β-actin mRNA levels were determined by semiquantitative RT-PCR in gastric biopsy samples as described in the Methods. PCR products were separated on 1.0% agarose gels. (A) A representative gel of samples from patients with intact mucosa without H pylori infection, in H pylori positive gastritis, and in H pylori positive gastric ulceration. (B, C) Densitometry of all samples from intact (n=4), H pylori positive gastritis (n=4) and H pylori positive gastric ulcer (n=4) tissues for COX-1 (B) and COX-2 (C), standardised against β-actin. *p<0.005 v intact mucosa (Fisher's PLSD test).
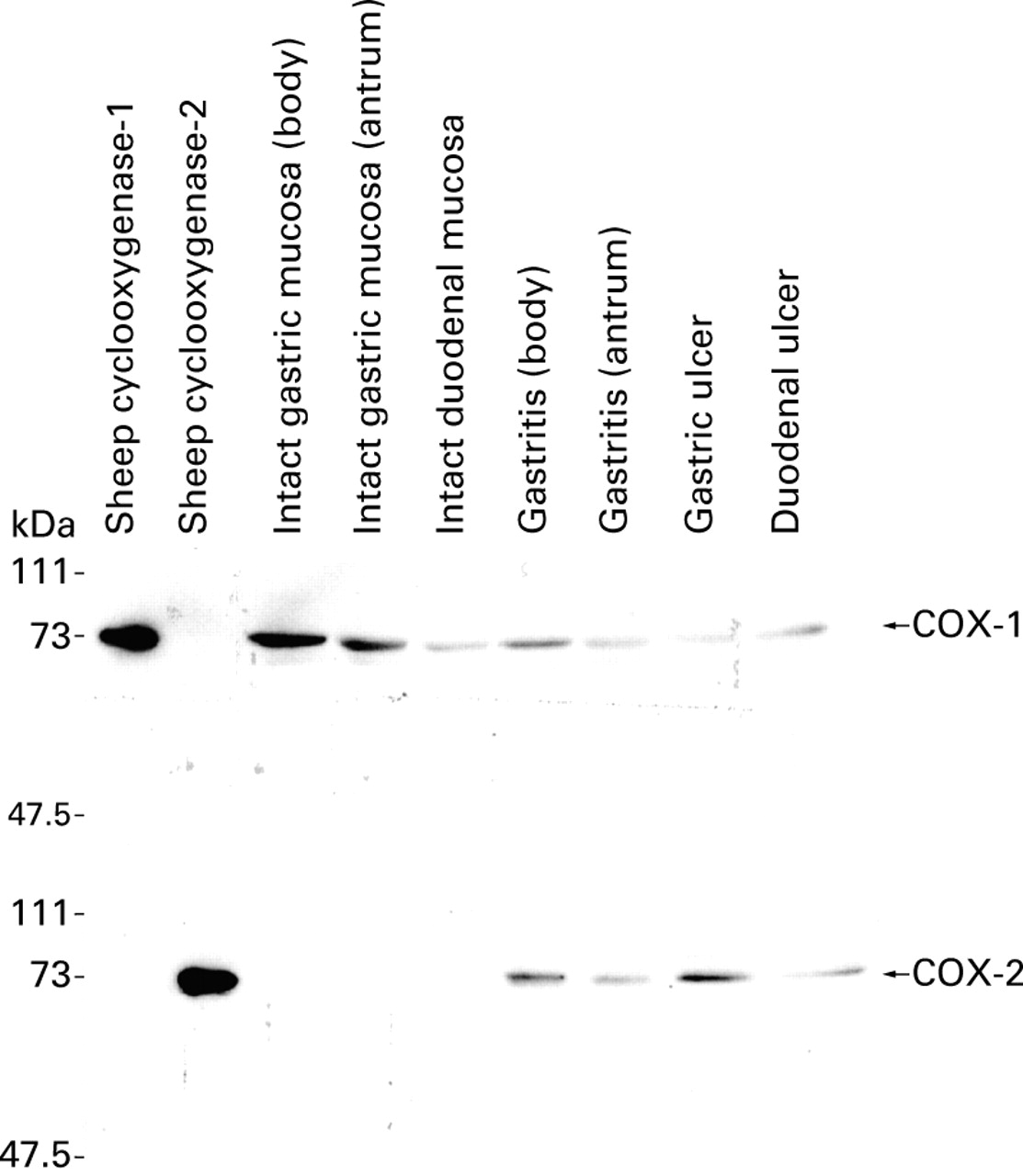
Elutes from anion exchange chromatography of gastric and duodenal tissue homogenates were immunoblotted with anti-COX-1 and anti-COX-2 antibodies. Anti-COX-1 antibody recognised sheep COX-1 but not COX-2. This antibody labelled a single band of 70 kDa molecular weight in samples from gastric and duodenal mucosa with or without ulceration. In contrast, anti-COX-2 antibody labelled one major band of 72 kDa in samples from gastric or duodenal ulcer margins. No protein band was detected in samples from intact gastric mucosa withoutH pylori infection or apparently intact duodenal mucosa without ulceration (fig 2).

Western blot analysis of COX-1 and COX-2 proteins. Purified sheep COX-1 and COX-2 proteins were included as controls. Anti-COX-1 antibody labelled a single band of 70 kDa molecular weight in samples from intact gastric mucosa (both body and antrum), intact duodenal mucosa, and gastric and duodenal ulcer edge. Anti-COX-2 antibody labelled a single band of 72 kDa molecular weight in samples from gastric and duodenal ulcer edge but not in any samples from intact gastric and duodenal mucosa.
IMMUNOLOCALISATION OF COX-1 AND COX-2 IN NORMAL GASTRIC MUCOSA AND GASTRIC ULCERS
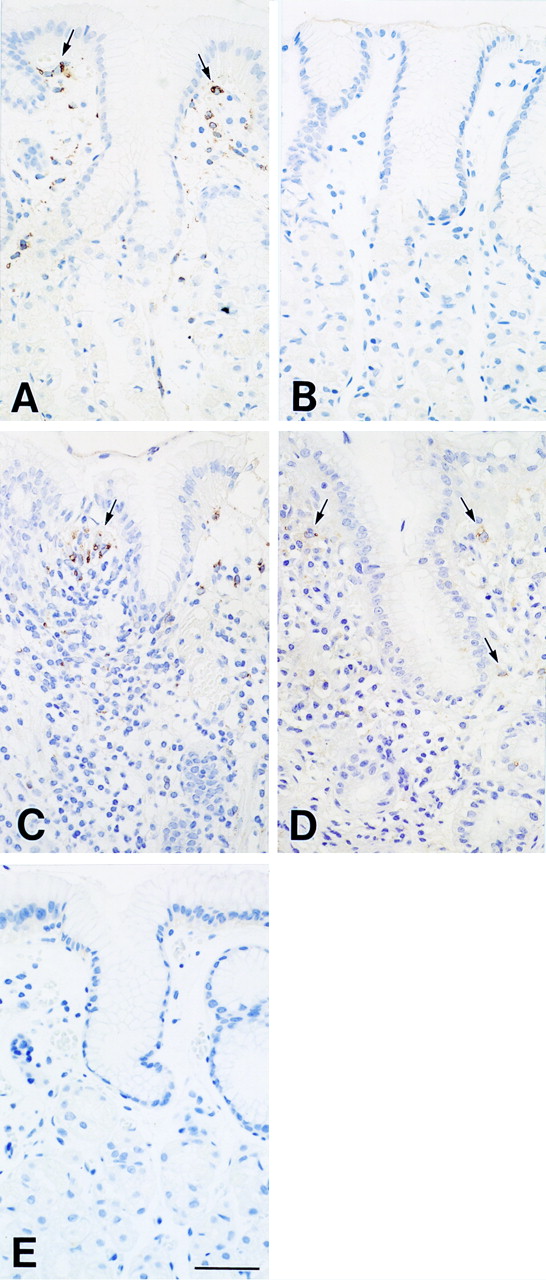
In gastric mucosa samples with or without H pylori related gastritis, COX-1 was clearly detected in scattered lamina propria mesenchymal cells corresponding to fibroblasts and macrophages (fig 3A,C). Less intense staining was observed in glandular cells such as parietal cells and in mucous cells in pyloric glands but not in surface epithelial cells. In contrast with COX-1 immunoreactivity, COX-2 immunoreactivity in gastric mucosa withH pylori related gastritis was localised only in mesenchymal cells (fig 3D). In the intact mucosa withoutH pylori related gastritis, no COX-2 immunoreactivity was noted in lamina propria mesenchymal cells (fig3B). Furthermore, COX-2 was strongly expressed in stomachs with ulceration. COX-2 immunoreactivity was exclusively localised in macrophages and fibroblasts between necrotic and granulation tissues of the ulcer bed in surgically resected gastric tissues (fig 4C, D), while COX-1 was negligible in all types of cells adjacent to the ulcer bed (fig 4A). COX-2 was detected preferentially in mesenchymal cells around arteries and capillaries but very weakly in endothelial cells (fig 4E). The mucosa immediately surrounding the ulcer showed chronic active inflammation, epithelial cells with various degrees of regenerative changes, and intestinal metaplasia. In contrast with the intact gastric mucosa and gastritis mucosa, COX-2 was observed in epithelial cells with regenerative changes and intestinal metaplasia (fig 4F). COX-2 immunoreactivity was also detected in lamina propria mesenchymal cells localised in ulcer margins but the density of positive cells in this area was markedly lower than in the ulcer bed. There was no obvious qualitative difference in COX-1 immunoreactivity between mucosa with gastritis and ulcer margin (fig 3C, fig 4B, b). No reaction was observed in any of the controls without anti-COX antibodies (fig 3E and fig 4G, H).

Immunohistochemical localisation of COX-1 (A, C) and COX-2 (B, D) in the intact gastric mucosa (A, B) and in H pylori related gastritis mucosa (C, D). COX-1 immunoreactivity is present in scattered lamina propria mesenchymal cells (arrows) in gastric mucosa with or without gastritis (A, C). COX-2 immunoreactivity is not present in the intact gastric mucosa (B) but in scattered lamina propria mesenchymal cells (arrows) in H pylori related gastritis mucosa (D). The control specimen, treated with non-immunised rabbit serum instead of the primary antibody, shows no reaction (E). Bar=50 μm

Immunohistochemical localisation of COX-1 (A, B, b) and COX-2 (C, D, E, F) in gastric ulcer tissues. (A) Low magnification view of the ulcer bed. Necrotic tissue (N) covers the surface of the ulcerated area. COX-1 was not stained in any type of cell in the ulcer bed. (B) COX-1 was stained in scattered lamina propria mesenchymal cells of the ulcer edge (arrows) as well as of H pylori related gastritis mucosa (see fig 3C) ((b) higher magnification view of (B)). (C) The same area as (A). COX-2 was strongly stained in many mesenchymal cells between necrotic (N) and granulation tissue of the ulcer bed. (D) Higher magnification view of (C). The immunoreactivity for COX-2 of both macrophages and fibroblasts varies from weak to strong. (E) Higher magnification view of (C). COX-2 was stained in mesenchymal cells around capillaries (arrow heads) in the ulcer bed. (F) COX-2 showed a fair amount of staining in epithelial cells of the ulcer edge. Arrows indicate regenerating epithelial cells and arrow heads intestinal metaplasia. (G) The same area as (A). The control specimen, treated with non-immunised rabbit serum instead of the primary antibody, shows no reaction. (H) The same area as (F). The control specimen, treated with non-immunised rabbit serum instead of the primary antibody, shows no reaction. N, necrotic tissue, ub, ulcer bed. Bars: A, B, C, F, G, H=100 μm; b, D, E=50 μm.
ELECTRON MICROSCOPIC IMMUNOHISTOCHEMICAL ANALYSIS FOR COX-1 AND COX-2
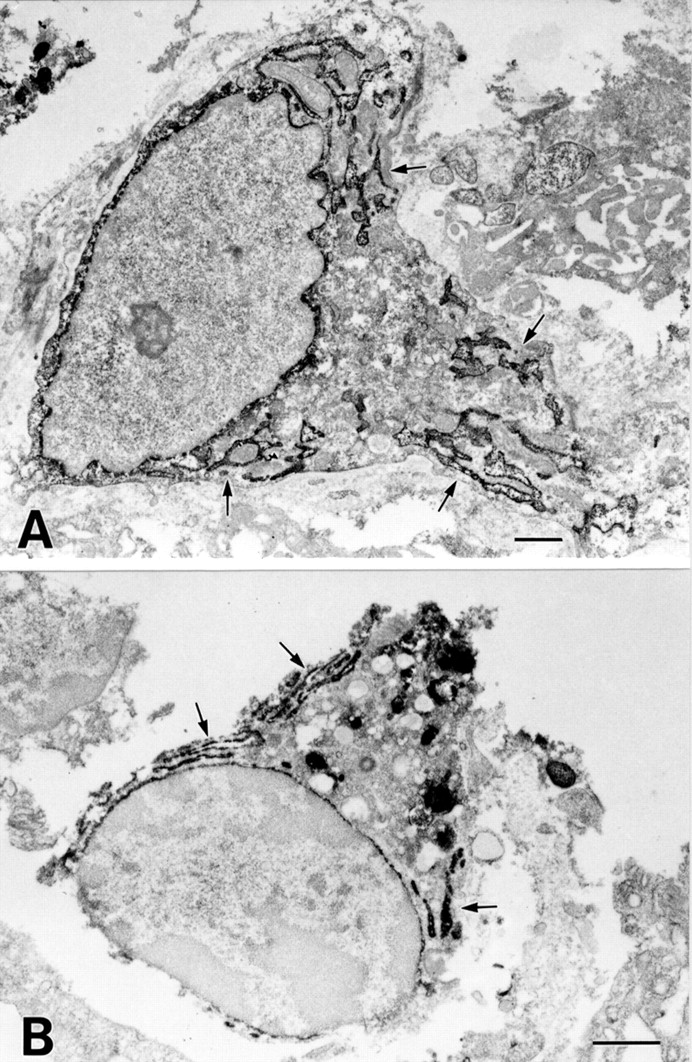
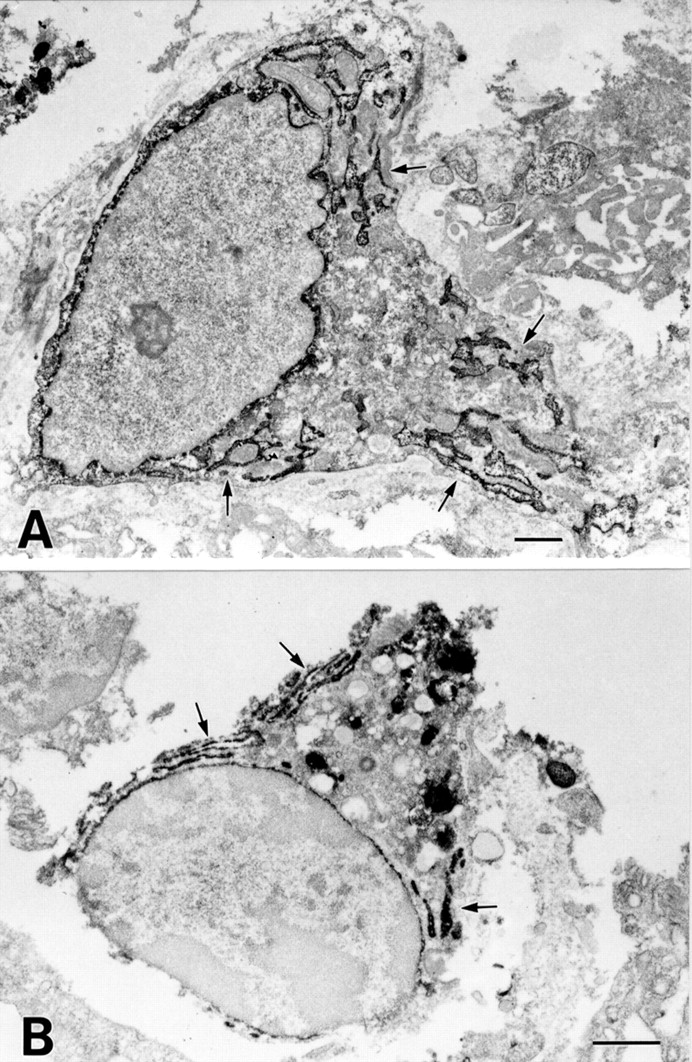
To confirm the results of light microscopic immunohistochemistry and detect differences in intracellular staining patterns between COX-1 and COX-2, we performed electron microscopic immunohistochemistry for the detection of COX-1 and COX-2. COX-1 immunoreactivity was observed on the nuclear membrane and in the endoplasmic reticulum (ER) of lamina propria mesenchymal cells, such as macrophages and fibroblasts (fig5A). The ER of other cells was also positive for COX-1 but the intensity was much less than that of mesenchymal cells. Similarly, COX-2 immunoreactivity was observed on the nuclear membrane and in the ER of lamina propria mesenchymal cells, such as macrophages and fibroblasts (fig 5B). These results indicate that COX-1 and COX-2 are membrane proteins of the ER and nuclear envelope, and reside in almost the same subcellular structures.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Electron microscopic immunohistochemical analysis of COX-1 and COX-2 in gastric ulcers. (A) COX-1 labelling is observed in the endoplasmic reticulum (arrows) and nuclear envelope in fibroblast-like cells. (B) COX-2 labelling is observed in the endoplasmic reticulum (arrows) and nuclear envelope in macrophage-like cells. There is no difference in the intracellular localisation of COX-1 and COX-2 in mononuclear cells. Bars=1 μm.
COMPARISON OF COX-1 AND COX-2 IMMUNOREACTIVITY IN GASTRITIS AND VARIOUS STAGES OF ULCER DISEASE
As COX-1 and COX-2 were mainly localised in mesenchymal cells, we examined COX-1 and COX-2 protein expression in biopsy specimens obtained from H pylori related gastritis and various stages of ulceration, focusing on immunoreactivity of mesenchymal cells. Almost all H pyloripositive patients with gastric ulcers at the acute stage showed a positive immunohistochemical reaction for COX-2. Increased COX-2 immunostaining in patients with gastric ulcers was still significant at the healing stage while immunoreactivity decreased at the scarring stage (table 1). The percentage of COX-2 expressing cells was significantly higher in open ulcers (acute and healing stages) than in closed ulcers (scarring stage) and in H pylori negative healthy individuals (tables 1, 2). In the intact mucosa without H pylori infection or ulceration, COX-2 immunoreactivity was not detected. In contrast, almost one-third of cases with H pylorirelated gastritis showed positive immunoreactivity for COX-2. The difference in COX-2 positivity between H pylori related gastritis and intact gastric mucosa was statistically significant. COX-1 immunoreactivity was not different among various stages of gastric ulcers and intact gastric mucosa (table1).
Positive rates of COX-1 and COX-2 immunoreactivities in gastric mucosa
Comparison of positive rates of COX-2 immunoreactivities between open and closed ulcers
Discussion
In the present study we have demonstrated expression of COX-2 mRNA and protein by semiquantitative RT-PCR and western blot analysis and the distribution of the COX-2 enzyme in human gastric mucosa by immunohistochemistry using either gastric ulcer tissues that were resected surgically or gastric biopsy samples fromH pylori positive or H pylori negative subjects. Expression of COX-2 mRNA and protein was detected in gastric ulcer tissues, confirming our previous results obtained in experimentally induced gastric ulcer tissues in mice.12 Furthermore, in this study, COX-2 enzyme was found to be strongly expressed in macrophages and fibroblasts exclusively localised between granulation and necrotic tissues of and around ulcer beds in the human stomach. This COX-2 protein distribution in human gastric ulcer tissues is consistent with previous results obtained from experimentally induced gastric ulcers in rats.15
We next examined and compared COX-2 immunoreactivity in biopsy samples either from the ulcer margin of the stomach in H pylori positive subjects or from intact gastric mucosa inH pylori negative subjects. The percentage of COX-2 expressing cells was significantly higher in samples obtained from the ulcer margin than in samples from either the ulcer scar or gastritis mucosa away from the ulcer margin. These data are consistent with our previous results that COX-2 mRNA and protein levels are increased during the acute stages of experimentally induced gastric ulcer tissues in mice.12 Several lines of evidence suggest that COX-2 expressed in inflamed tissues may play a pivotal role in tissue repair processes. Firstly, administration of NS-398, a specific anti-inflammatory agent against COX-2 enzyme, to mice with ulcers at acute stages resulted in impairment of ulcer healing.12Secondly, interleukin 1, an inflammatory cytokine, induces not only COX-2 expression but also hepatocyte growth factor (HGF) production, and production of the latter in human gastric fibroblasts is inhibited by blockade of COX activity by NSAIDs.25 Thirdly, PGE2 causes marked acceleration of restitution which is completely inhibited by anti-HGF antibody, as shown in an in vitro cultured gastric mucosal model consisting of gastric epithelial cells and fibroblasts.26 Furthermore, the sites of HGF mRNA expression previously reported in stromal cells around regenerative glands and in arterial vessels of submucosal tissues are similar to those of COX-2.27 Fourthly, COX-2 is also involved in the production of angiogenic factors.28 Among such angiogenic factors, basic fibroblast growth factor (bFGF) is localised in fibroblasts and capillary endothelium in granulation tissue and epithelium at ulcer margins.29 Taking into consideration the above early findings and those of the present study, it is possible that COX-2 may promote angiogenesis and accelerate restitution in the ulcer bed and be involved in gastric ulcer healing.
We have previously shown that growth stimulation of primary cultured gastric surface epithelial cells by EGF induces COX activity.10 11 Recent studies have also shown thatH pylori induces COX-2 mRNA expression in gastric cancer epithelial cells in vitro30 or COX-2 protein expression in surface epithelial cells of gastric mucosa in vivo.18 19 In the present study, the percentage of COX-2 expressing cells in gastritis mucosa away from the ulcer margins inH pylori positive subjects was significantly higher than in H pylori negative subjects, suggesting that H pylori related gastritis alone induces COX-2 protein expression in the gastric mucosa. Although we identified COX-2 immunoreactivity in regenerating epithelial cells and intestinal metaplasia lining close to the ulcer base, its expression was rather weak compared with that in mesenchymal cells and we did not find COX-2 immunoreactivity in surface or foveolar epithelial cells infected with H pylori.This distribution of COX-2 immunoreactivity in gastritis is clearly different from that in colitis. COX-2 immunoreactivity is strongly induced not only in lamina propria mononuclear cells but also in apical epithelial cells of the colonic mucosa in Crohn's colitis and ulcerative colitis.31 Compared with H pylori related gastritis, Crohn's colitis is associated with inflammation extending through all layers of the gut wall. Ulcerative colitis is also known to induce diffuse erosions and ulcers in the colonic mucosa. Thus inflammatory reactions of the gastrointestinal mucosa seem generally to be stronger in active Crohn's enterocolitis and ulcerative colitis than in H pylorirelated gastritis. Therefore, the difference in the extent of inflammatory reaction of the mucosa may be the reason for differences in the intensity of COX-2 protein expression between colitis and gastritis epithelial cells. As overexpression of the COX-2 gene in intestinal epithelial cells is associated with resistance to apoptosis32 and COX-2 gene knockout is related to reduced size and number of intestinal polyps in mice,33 the strong and persistent COX-2 expression in colonic epithelium of inflammatory bowel diseases may be involved in the increased risk of carcinogenesis in the colonic epithelium.31 In contrast, COX-2 protein would not be directly related to the risk of gastric cancer noted in patients with H pylori related gastritis.
Our results also showed the presence of COX-1 immunoreactivity in lamina propria mononuclear cells. Immunoreactivity was also observed in glandular cells such as parietal cells, but the intensity of COX-1 immunoreactivity in these glandular cells was weak compared with that in lamina propria mononuclear cells. The intensity and number of positive cells in the lamina propria mesenchymal cells were not increased even in ulcer margins. COX-1 positive cells in lamina propria were still observed in the intact gastric mucosa withoutH pylori infection but COX-2 positive cells were not observed, suggesting that COX-1 plays a major role in protecting the mucosa against injury in the intact stomach.
In the present study, we examined subcellular localisation of COX-1 and COX-2 proteins by electron microscopic immunohistochemistry. However, despite the difference in the distribution of COX-1 and COX-2 protein in the gastric mucosa, both proteins were similarly localised on the luminal surface of the ER and nuclear envelope in mononuclear cells. Although earlier immunofluorescence studies suggested that COX-2 protein was more concentrated in the nuclear envelope than in the ER,34 our findings are consistent with a recent report showing similar subcellular localisation of COX-1 and COX-2 protein in human monocytes.35 We therefore conclude that the difference between the role of COX-1 and COX-2 proteins in human stomach may be related to the difference in their distribution in the gastric mucosa rather than subcellular localisation.
References
Footnotes
- Abbreviations used in this paper:
- COX
- cyclooxygenase
- NSAID
- non-steroidal anti-inflammatory drug
- ER
- endoplasmic reticulum
- PG
- prostaglandin
- HGF
- hepatocyte growth factor
- EGF
- epidermal growth factor
- bFGF
- basic fibroblast growth factor
- RT-PCR
- reverse transcription-polymerase chain reaction
- DAB
- 3,3′-diaminobenzidine tetrahydrochloride