Article Text

Abstract
BACKGROUND In murine models of inflammatory bowel disease, colonic inflammation is considered to be caused by an aberrant Th1-type immune response.
AIM To investigate if systemic administration of interleukin (IL)-12 and IL-18 to wild-type BALB/c mice induces liver injury and intestinal inflammation, and if pathological changes are observed, what cytokines are involved.
METHODS Mice (BALB/c-wild-type (wt), MRL-lpr/lpr, BALB/c-interferon γ knock out (IFN-γ KO), C57BL/6-inducible nitric oxide synthase (iNOS) KO, and BALB/c tumour necrosis factor α (TNF-α) KO) were injected intraperitoneally each day with IL-12 (20 ng/g/mouse) and/or IL-18 (200 ng/g/mouse).
RESULTS Administration of IL-12 and IL-18 to BALB/c-wt mice induced prominent intestinal mucosal inflammation and fatty liver, leading to piloerection, bloody diarrhoea, and weight loss. IL-12 and IL-18 induced striking elevations in serum levels of IFN-γ that caused NO production, although increased NO had no exacerbating effect on mice. Moreover, iNOS KO mice, or MRL lpr/lpr mice lacking functional Fas were equally susceptible to IL-12 and IL-18. Administration of IL-12 and IL-18 did not induce TNF-α production in wild-type mice, and the same treatment to TNF-α KO mice induced intestinal mucosal inflammation. Furthermore, they had diffuse and dense infiltration of small fat droplets in their hepatocytes associated with an increase in serum levels of liver enzymes. In contrast, the same treatment in IFN-γ KO BALB/c mice and iNOS KO mice did not induce these changes.
CONCLUSIONS Our study strongly indicates that IL-18 together with IL-12 induces intestinal mucosal inflammation in an IFN-γ dependent but TNF-α, NO, and Fas ligand independent manner, and fatty liver is dependent on IFN-γ and NO.
- interleukins
- interferon γ
- intestinal inflammation
- fatty liver
- mice
Abbreviations used in this paper
- IFN-γ
- interferon γ
- IL
- interleukin
- LPS
- lipopolysaccharide
- L
- ligand
- wt
- wild type
- TNF-α KO
- tumour necrosis factor α knock out
- iNOS KO
- inducible nitric oxide synthase knock out
- NO
- nitric oxide
- PBS
- phosphate buffered saline
- ALT
- alanine transaminase
Statistics from Altmetric.com
CD4+ T helper cells can be divided into Th1 and Th2 cells on the basis of their cytokine production profile.1Their development depends on the mode of priming: interleukin (IL)-12 and IL-4 induce differentiation of naive T cells towards Th1 or Th2 cells, respectively.2 ,3 Macrophages are an essential immunological component of host defences and induce development of Th1 cells by production of IL-12,2 ,4-9 but also have the potential to induce host injury by production of proinflammatory cytokines.2-9 IL-1 and tumour necrosis factor α (TNF-α), products of activated macrophages, induce lethal shock and evoke serious multiple organ failures when injected intravenously or tissue injury when locally injected.7-9 IL-18, originally called interferon γ (IFN-γ) inducing factor, is a cytokine of 18 kDa synthesised by Kupffer cells and activated macrophages.4 ,10 ,11 Although IL-18 cannot induce Th1 cell development,12 ,13 this cytokine has a marked capacity to induce IFN-γ production by Th1 cells, especially in the presence of IL-12.4 ,11-16 In addition, IL-18 activates natural killer cells.4 ,10 Recently, we investigated the molecular mechanism of the synergistic effect of IL-12 and IL-18 on IFN-γ production.16 ,17 We revealed that IL-12 stimulation rendered T cells responsive to IL-18 by induction of IL-18 receptor.17 We also showed that Th1 or Th2 cells exhibit differential IL-18 responsiveness and that only the former cells express IL-18 receptor.17
Mice are relatively resistant to lipopolysaccharide (LPS) but in some circumstances they become highly susceptible to LPS.5 ,6Indeed, sequential administration ofPropionibacterium acnes and LPS induces severe liver injury.4 ,6 ,18 We showed previously that IL-18 initiates the development of LPS induced liver injury inP acnes primed mice by induction of Fas ligand (L) and TNF-α production in the liver.18 IL-18 directly induces Fas L expression19whereas TNF-α production is IFN-γ dependent.18 We and others showed that injection of anti-IL-12 or anti-IFN-γ antibody protectedP acnes primed mice from LPS induced liver injury.18-20 These results indicate that IL-12 and IL-18 play a critical role in the pathogenesis of LPS induced liver injury.
Several rodent models of chronic intestinal inflammation share features of immunopathology with human inflammatory bowel disease, most of which show evidence of aberrant Th1 responses.21-26 In these models, colonic inflammation is considered to be caused by a dysregulated Th1 response but some are caused by alteration of the balance between the effects of IFN-γ and transforming growth factor β.26-28 Th-2 mediated colitis has also been reported.29 ,30 More recently, it has been demonstrated that the development of pathogenic T cells is influenced predominantly by IL-12 via the action of signal transducer and activator of transcription-4 proteins, and IFN-γ is required to establish colitis.31 But it has also been demonstrated that other inflammatory mediators are sufficient to mediate disease.31 To determine the pathological role of IL-12 and IL-18 in vivo, we investigated if administration of IL-12 and IL-18 induces IFN-γ production that may promote mucosal inflammation and liver injury with hepatitis. We also investigated if other inflammatory factors, such as TNF-α and nitric oxide (NO), are involved in mucosal inflammation and if IFN-γ knockout (KO) mice are resistant to this treatment.
Materials and methods
MICE
Specific pathogen free BALB/c mice, designated BALB/c-wild-type (wt), and MRL lpr/lpr mice (female, 7–8 weeks of age) were obtained from Shizuoka Laboratory Animal Centre (Shizuoka, Japan). IFN-γ KO mice32 were backcrossed to BALB/c mice for F8 generations, and the resultant female BALB/c-IFN-γ KO mice at 6–8 weeks old were used in this study. Inducible nitric oxide synthase knock out (iNOS KO) mice with C57BL/6 background (female, 7–8 weeks old) were obtained from Jackson Laboratory (Bar Harbor, Maine, USA). TNF-α KO mice were produced as described previously.32
REAGENTS AND ANTIBODY
Recombinant mouse IL-12 and IL-18 were produced and prepared as described previously.4 ,33
IN VIVO TREATMENT OF MICE
Mice (BALB/c-wt, MRL-lpr/lpr, BALB/c-IFN-γ KO, C57BL/6-iNOS KO, BALB/c-TNF-α KO, each group n=5) were injected intraperitoneally each day with 20 ng/g/mouse of IL-12 and/or 200 ng/g/mouse of IL-18 in 200 μl of phosphate buffered saline (PBS). Control mice were injected intraperitoneally each day with 200 μl of PBS alone. Mice were weighed daily and monitored for appearance and signs of illness, such as loose stools, diarrhoea, and physical activity. Mice were sacrificed on day 5 after injection and their organs were examined after fixation. Before sacrifice, sera were sampled, aliquoted, and frozen at −20°C until measurement of alanine transaminase (ALT), cytokines, and NO levels. Serum levels of liver enzyme, cytokines, and NO are presented as mean (SEM) values of five mice.
HISTOLOGICAL ANALYSIS
Liver specimens were sampled on day 5 at the indicated times after the last injection of IL-12 and/or IL-18 and fixed in 3.5% formaldehyde in PBS. Sections (2 μm) were cut and placed on slides for staining with haematoxylin and eosin using a standard technique. Hepatocytes were also examined for the presence of lipid by staining with oil red O. Specimens were fixed in 10% neutral buffered formalin. After fixation, they were frozen at −20°C, sliced (2–4 μm), and stained with oil red O. Colon tissues were simultaneously sampled and fixed in Zamboni's solution and embedded in paraffin. Sections (4 μm) were cut and placed on slides and stained with haematoxylin and eosin. For histological scoring of liver injury, 10 fields were evaluated at ×40 magnification.
For electron microscopy, the liver was perfused via the portal vein with saline for 20 seconds, and then with the prefixative containing 1.5% glutaraldehyde, 0.067 M cacodylate buffer (pH 7.4), and 1% sucrose (310 mosmol) for 1–2 minutes at a flow rate of 10 ml/min at room temperature. Tissue blocks from the liver were postfixed in 2% OsO4 in phosphate buffer (pH 7.4) at 4°C for two hours, further processed with 2% uranyl acetate for 20 minutes at room temperature, dehydrated in ethanol series, and embedded in Epon. Ultrathin sections were stained with saturated uranyl acetate and lead citrate, and observed under a JEOL 100C× electron microscope at 100 KV.
ELISA
Serum concentration of IFN-γ and TNF-α were determined using ELISA kits (Genzyme, Cambridge, Massachusetts, USA).
NO ASSAY
Serum levels of NO were measured using Griess reagent (Dojin, Chm. Lab. Institute, Kumamoto, Japan).
STATISTICAL ANALYSIS
The paired two tailed Student's t test was performed for statistical analysis. Probabilities of 5% or less were considered statistically significant.
Results
IL-12 AND IL-18 INDUCE WEIGHT LOSS IN AN IFN-γ DEPENDENT MANNER
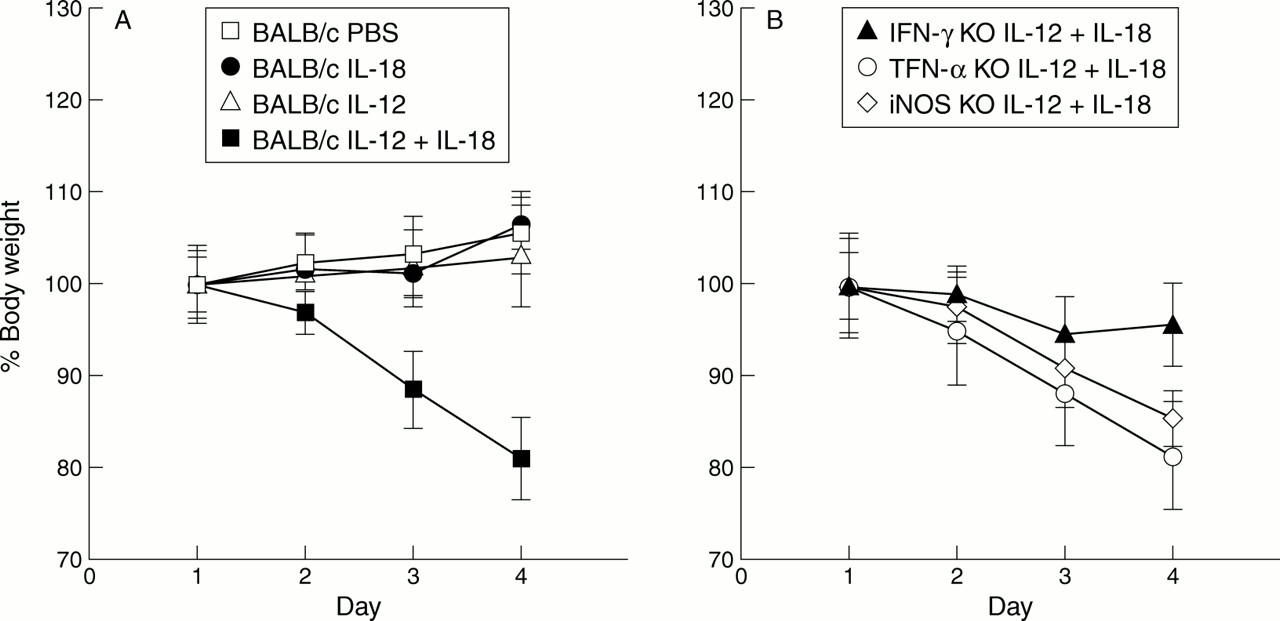
We first tested if injection of IL-12 and/or IL-18 induced weight loss in an IFN-γ dependent manner. For this purpose, we examined the effect of IL-12 and/or IL-18 on the gross clinical symptoms of BALB/c-wt, TNF-α KO, iNOS KO, and IFN-γ KO mice. BALB/c-wt mice injected daily with IL-12 (20 ng/g/mouse) or IL-18 (200 ng/g/mouse) retained a healthy appearance during monitoring. In contrast, BALB/c-wt, TNF-α KO, and iNOS KO mice injected daily with a mixture of IL-12 and IL-18 showed weight loss and overt clinical symptoms, such as piloerection and in most cases diarrhoea which was occasionally accompanied by visible blood. Furthermore, these symptoms advanced progressively, and eventually almost all mice died by day 6. Interestingly, the same treatment to BALB/c-IFN-γ KO mice caused only slight weight loss. Weight loss of wild-type mice injected with IL-12 and IL-18 was clearly illustrated (fig 1A), and TNF-α KO and iNOS KO mice given the same treatment also showed a marked decrease in weight (fig 1B). However, IFN-γ KO mice given the same treatment showed a moderate decrease in weight (fig 1B). TNF-α KO, iNOS KO, and IFN-γ KO mice injected with PBS showed no clinical symptoms or weight loss (data not shown). This decrease in weight in wild-type, TNF-α KO, and iNOS KO mice but not in IFN-γ KO mice after injection of IL-12 and IL-18 was significant (p<0.01).

(A) Relative body weight of BALB/c and IFN-γ KO mice injected with each reagent. Weights of BALB/c mice injected with PBS (200 μl), IL-18 (200 ng/g/mouse), IL-12 (20 ng/g/mouse), IL-12 (20 ng/g/mouse) plus IL-18 (200 ng/g/mouse) are shown. Each group consists of five mice. The weight of BALB/c mice injected with IL-12 (20 ng/g/mouse) together with IL-18 (200 ng/g/mouse) on day 4 was 82.0% of that of the first day and this weight loss was significant (p<0.01). (B) Relative body weight of IFN-γ KO, TNF-α KO, and iNOS KO mice injected with IL-12 (20 ng/g/mouse) plus IL-18 (200 ng/g/mouse). Each group consists of five mice. The weight of IFN-γ KO mice given the same treatment on day 4 was 95.2% of that of the first day and weight loss was not significant. Weight loss in the other two groups was significant. Each group of mice injected with PBS (200 μl) daily showed no pathological findings and no weight loss (data not shown).
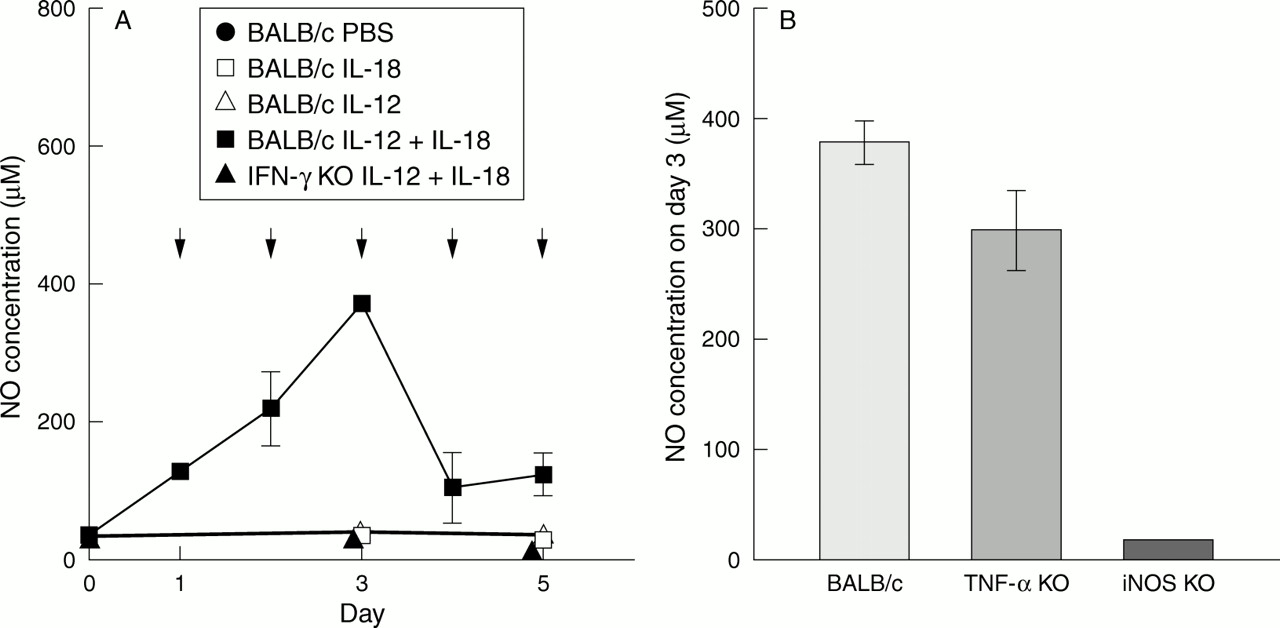
We next examined the capacity of IL-12 and IL-18 to induce IFN-γ production in wild-type, TNF-αKO, and iNOS KO mice. As shown in fig 2A, wild-type mice showed markedly increased levels of IFN-γ in sera. Kinetic studies revealed that levels of IFN-γ were highest on day 3 (35.10 (0.05) ng/ml) after the start of daily injections of a mixture of IL-12 and IL-18. But these levels declined thereafter and became very low on days 4 and 5. TNF-α KO and iNOS KO mice also showed increased levels of IFN-γ on day 3 (fig 2B). Levels of NO correlated well with those of IFN-γ in wild-type mice injected with IL-12 and IL-18 (fig 3A). In TNF-α KO mice, IFN-γ produced by stimulation with IL-12 and IL-18 also showed high levels of NO on day 3 (fig 3B). Furthermore, IFN-γ KO mice showed no such increase in serum levels of NO after IL-12 and IL-18 treatment (fig 3A), suggesting that IL-12 and IL-18 synergistically induce IFN-γ production that stimulates macrophages and other types of cells to produce NO. However, IFN-γ induced NO production was not pathogenic because reagents such as aminoguanidine that prevent iNOS dependent NO production failed to protect BALB/c-wt mice from IL-12 and IL-18 induced weight loss (data not shown). Moreover, iNOS KO mice were equally susceptible to IL-12 and IL-18 in the intestines (fig 5C, F). However, in the liver of iNOS KO mice injected with IL-12 and IL-18, the pathological changes and elevation of ALT were less than those of wild-type and TNF-α KO mice (figs 6E, 7). TNF-α was not elevated in the sera of mice injected daily with IL-12 and IL-18 (data not shown). TNF-α KO mice given the same treatment manifested weight loss (fig 1B) accompanied by intestinal inflammation (fig 5B, E). MRL lpr/lpr mice were also susceptible to treatment with IL-12 and IL-18, with weight loss accompanied by intestinal inflammation (fig 5A, D). Thus IL-12 and IL-18 induced weight loss and intestinal inflammation in an IFN-γ dependent but TNF-α, NO, and Fas L/Fas independent manner.

(A) IFN-γ concentrations in the serum of BALB/c mice injected with IL-12 (20 ng/g/mouse) plus IL-18 (200 ng/g/mouse), IL-12 (20 ng/g/mouse), IL-18 (200 ng/g/mouse), or PBS (200 μl) in addition to IFN-γ KO mice given the same treatment. Each group consists of five mice. Samples were collected six hours after injection every day. IFN-γ production was observed only in the serum of BALB/c mice injected with IL-12 plus IL-18; it increased daily until day 4 and then decreased on day 5. Arrows show injection of IL-12 plus IL-18. (B) IFN-γ concentrations in the serum of BALB/c, TNF-α KO, and iNOS KO mice injected with IL-12 (20 ng/g/mouse) plus IL-18 (200 ng/g/mouse) daily. Each group consists of five mice. Samples were collected six hours after injection on day 3. High levels of IFN-γ production were observed in the serum of TNF-α KO and iNOS KO mice, and were higher than those of BALB/c mice.

(A) NO concentrations in the serum of BALB/c mice injected with PBS (200 μl), IL-18 (200 ng/g/mouse), IL-12 (20 ng/g/mouse), IL-12 (20 ng/g/mouse) plus IL-18 (200 ng/g/mouse), and IFN-γ KO mice injected with IL-12 (20 ng/g/mouse) plus IL-18 (200 ng/g/mouse). Each group consists of five mice. Samples were collected six hours after injection. Prominent production of NO is observed only in the serum of BALB/c mice injected with IL-12 plus IL-18. It appears that NO production was elevated and decreased in parallel with IFN-γ production in BALB/c mice treated with IL-12 plus IL-18. Arrows show injections of IL-12 plus IL-18. (B) NO concentrations in the serum of BALB/c, TNF-α KO, and iNOS KO mice injected with IL-12 (20 ng/g/mouse) plus IL-18 (200 ng/g/mouse) daily. Each group consists of five mice. Samples were collected six hours after injection on day 3. High levels of NO production were observed in the serum of BALB/c and TNF-α KO mice. Low levels of NO production were observed in iNOS KO mice.

Histology of the jejunum (A–C); haematoxylin-eosin ×350. (A) Jejunum of MRL lpr/lpr mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). (B) Jejunum of TNF-αKO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). (C) Jejunum of iNOS KO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). In all of these jejunums, a decrease in the height of the villi and infiltration of inflammatory cells were observed. Histology of the colon (D–F); haematoxylin-eosin ×350. (D) Colon of MRL lpr/lpr mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). (E) Colon of TNF-α KO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). (F) Colon of iNOS KO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). In all of these colons, a decrease in the number of goblet cells was observed. Prominent infiltration of the inflammatory cells was observed in the colon of the MRL lpr/lpr mouse.

Histology of the liver (A–F); haematoxylin-eosin ×350. (A) Liver of BALB/c mouse injected with PBS. (B) Liver of BALB/c mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). Multiple small fat vacuoles occupied the cytoplasm of hepatocytes. The arrow indicates an apoptotic cell. (C) Liver of TNF-α KO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 ( 200 ng/g/mouse). Multiple small fat vacuoles also occupy the cytoplasm of hepatocytes. (D) Liver of IFN-γ KO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). (E) Liver of iNOS KO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). (F) Liver of MRL lpr/lpr mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). Multiple small fat vacuoles occupy the cytoplasm of hepatocytes. (G) Electron micrograph of an apoptotic hepatocyte. A shrunk apoptotic cell detached from neighbouring cells with condensed cytoplasm (bar=2 μm).
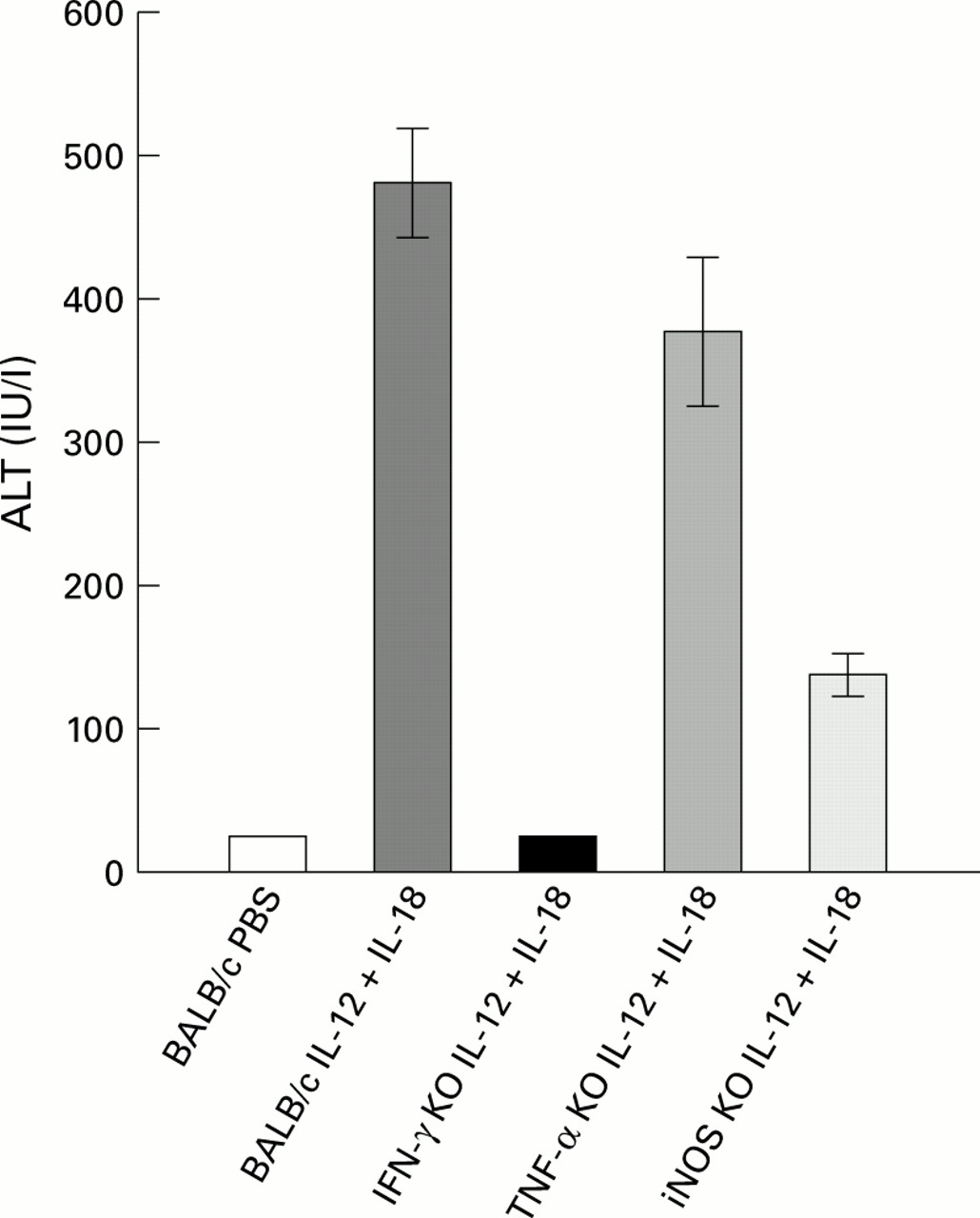
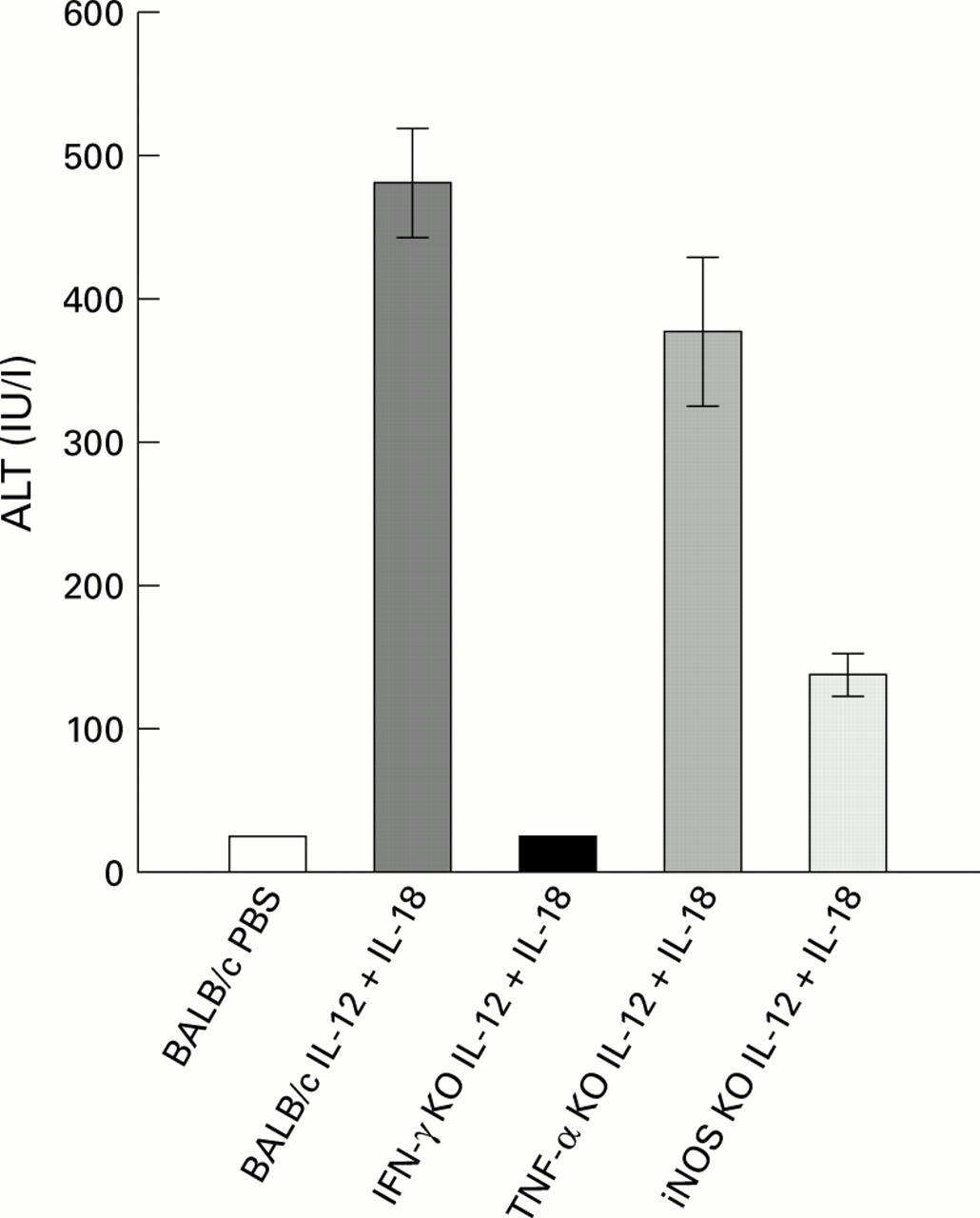
Serum alanine transaminase (ALT) levels (mean (SEM)). ALT levels in the sera of BALB/c-wt, IFN-γ KO, TNF-α KO, and iNOS KO mice (five mice), and in BALB/c wt injected with PBS (five mice). Samples were collected six hours after injection on day 5. Highly elevated levels of ALT were observed in BALB/c-wt and TNF-α KO mice. Moderately elevated levels of ALT were observed in iNOS KO mice and normal levels in IFN-γ KO mice treated with IL-12 and IL-18 and BALB/c wt mice treated with PBS.
HISTOPATHOLOGY OF SMALL AND LARGE INTESTINES IN MICE INJECTED WITH IL-12 AND IL-18
Next, requirement for IFN-γ for induction of these clinical symptoms was tested by comparing pathological changes in wild-type and IFN-γ KO mice after treatment with IL-12 and IL-18 (figs 4, 6). As inferred from the severe diarrhoea seen in wild-type mice injected with IL-12 and IL-18, the small intestine, especially the jejunum, was severely damaged. There were erosions with shortened villi, elongated crypts, and diminished number of goblet cells and Paneth cells in crypts (fig 4B), while IFN-γ KO mice were free of these pathological changes (fig 4C). Additional common histological findings in the crypts of the small intestine obtained from wild-type mice treated with IL-12 and IL-18 were increases in the numbers of mitotic and apoptotic cells (data not shown). We also observed diffuse infiltration of inflammatory cells, including lymphocytes, plasma cells, and neutrophils in the mucosa. Similar changes were also observed in the small intestines of MRL lpr/lpr (fig 5A), TNF-α KO (fig 5B), and iNOS KO mice (fig 5C) when they were injected with IL-12 and IL-18. In contrast, IFN-γ KO mice given the same treatment did not show these changes (fig 4C) suggesting that IL-12 and IL-18 induce mucosal inflammation in an IFN-γ dependent but TNF-α, NO, and Fas L/Fas independent manner.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Histology of the jejunum (A–C); haematoxylin-eosin ×350. (A) Jejunum of BALB/c mouse injected with PBS. (B) Jejunum of BALB/c mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). (C) Jejunum of IFN-γ KO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). Histology of the colon (D–F); haematoxylin-eosin ×350. (D) Colon of BALB/c mouse injected with PBS. (E) Colon of BALB/c mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse). (F) Colon of IFN-γ KO mouse injected with both IL-12 (20 ng/g/mouse) and IL-18 (200 ng/g/mouse).
Treatment of wild-type mice with IL-12 and IL-18 also induced marked colonic mucosal inflammation whereas the same treatment of IFN-γ KO mice did not. Histological study revealed the presence of elongated crypts and multiple mucosal erosions (fig 4E). Again, induction of these pathological changes depended on the synergistic action of IL-12 and IL-18. Neither IL-12 nor IL-18 induced such changes (data not shown). The crypts were characterised by an increase in the number of mitotic as well as apoptotic cells. Furthermore, a marked decrease in the number of goblet cells was often observed, presumably leading to mucin depletion. The number of inflammatory cells, including lymphocytes, plasma cells, and neutrophils, increased in the mucosa, and crypt abscesses were also found (fig 4E). In contrast with wild-type mice treated with IL-12 and IL-18, IFN-γ KO mice were resistant to the action of IL-12 and IL-18 in inducing mucosal inflammation in the colon (fig 4F). Microscopic changes were also observed in the colons of TNF-α KO, MRL lpr/lpr, and iNOS KO mice treated with IL-12 and IL-18 (fig 5D–F).
HISTOPATHOLOGY OF THE LIVER IN MICE INJECTED WITH IL-12 AND IL-18
Treatment of P acnes primed mice with LPS induces severe liver injury.4 ,6 ,18 Anti-IL-12 antibody and/or anti-IL-18 antibody blocked liver injury induced byP acnes and LPS,18 ,20indicating that IL-12 and IL-18 play critical roles in the pathogenesis of LPS induced liver injury. Therefore, we investigated the capacity of IL-12 and IL-18 to induce liver injury in BALB/c-wt mice. We also tested the requirement for IFN-γ in induction of liver injury by using BALB/c IFN-γ KO mice. We administered IL-12 and/or IL-18 daily to BALB/c-wt and IFN-γ KO mice for four days and examined pathological changes in their livers.
Two hours after the last injection of IL-12 and/or IL-18, mice were sacrificed for histological analysis. In the liver of BALB/c-wt mice injected with IL-12 and IL-18 for four days, hepatocytes were enlarged, and most dramatically, their cytoplasm was occupied by multiple small vacuoles which surrounded a central nucleus (fig 6B). In addition, apoptotic cells containing condensed and unevenly distributed chromatins were sporadically found among these hepatocytes (fig 6B). In contrast with the histology of the liver of P acnes primed and LPS challenged mice,18 ,20infiltration of mononuclear cells was limited and only a small number of mononuclear cells were observed in the lobes. Staining frozen sections with oil red O showed positive stain of these vacuoles (data not shown), indicating that these droplets were lipid. Hepatocytes were also examined with an electron microscope. Figure 6G shows an electron micrograph of an apoptotic cell in the liver of BALB/c-wt mice administered IL-12 and IL-18 for four days. An apoptotic cell detached from neighboring cells and the cytoplasm condensed, reserving cellular integrity was observed. Furthermore, these mice showed increased levels of ALT in sera (fig 7). Similar pathological changes were observed in the livers of TNF-α KO mice (fig 6C) who also showed increased levels of ALT in sera (fig 7). MRL lpr/lpr mice given the same treatment also showed fatty degeneration in the liver (fig 6F). The liver of iNOS KO mice injected with IL-12 and IL-18 did not show pathological fatty changes (fig 6E). Also, increased levels of ALT were less than those of wild-type and TNF-α KO mice (fig 7). These findings indicate that NO has some role in inducing fatty degeneration of the liver. In contrast, IFN-γ KO mice injected with IL-12 and IL-18 did not show pathological changes in their livers (fig 6D), and ALT levels in sera were almost the same as those in PBS injected mice (fig 7), indicating that IFN-γ KO mice were also resistant to the action of IL-12 and IL-18 in inducing fatty liver degeneration.
Discussion
We have shown that administration of a mixture of IL-18 and IL-12 to wild-type mice induced weight loss and intestinal inflammation in an IFN-γ dependent manner. These mice displayed marked histopathological changes such as erosions of the small intestines as well as fatty degeneration of the liver (fig 4B, E; fig 6B). Induction of these changes required combined administration of IL-18 and IL-12. We previously reported a synergistic effect of IL-18 and IL-12 on IFN-γ production by T cells.10 ,11 ,16 ,17 Our molecular analysis revealed that IL-12 rendered T cells responsive to IL-18 by induction of IL-18 receptor expression.17 Apart from this mechanism, IL-18 and IL-12 were shown to independently activate different transcription factors (nuclear factor κB and activator protein 1), which in combination may synergistically activate the promoter site of the IFN-γ gene.16 ,34 ,35
IL-12 and IL-18 induced mucosal inflammation was characterised by the presence of villous atrophy, and loss of Paneth and goblet cells (fig4B, E). This inflammation was also associated with an increase in the numbers of apoptotic and mitotic cells (data not shown). Less frequent findings included erosions of mucosa. We also showed that injection of a mixture of IL-12 and IL-18 induced high levels of IFN-γ production and mucosal inflammation with a fatty liver in wild-type but not IFN-γ KO mice, clearly indicating that IFN-γ has an important role in the induction of these pathological changes. This is considered to be Th1 mediated colitis and IFN-γ has an important role. However, injection of high doses of IFN-γ (10 μg/mouse) failed to replace the action of IL-12 and IL-18 (data not shown), suggesting that IFN-γ may require the presence of other factors induced by the action of IL-12 and IL-18 to have pathological effects.
In some forms of chronic experimental colitis, the development of pathogenic T cells is influenced by the IL-12/IFN-γ pathway.31 Hence IL-12 alone may be able to induce colitis in mice. We injected IL-12 (20 μg/g/mouse) daily into mice but they did not show pathological changes; higher doses may be necessary. Recently, there have been some reports of Th2-mediated colitis29 ,30 and IL-18 may potentiate the Th2 responses.36-38 Hence high doses of IL-18 may induce colitis in mice. We injected IL-18 5 μg/mouse daily for five days and observed clinical changes but there were no pathological changes such as weight loss or diarrhoea (data not shown). In this study, we injected both IL-12 and IL-18 to induce colitis but this may not be an absolute requirement.
In this study, we found that in common with BALB/c-wt mice treated with IL-12 and IL-18, MRL-lpr/lpr mice lacking functional Fas due to mutation suffered weight loss with diarrhoea and manifested intestinal inflammation after treatment. This suggests that these pathological changes were induced in a Fas independent manner. Further studies are needed to elucidate factors other than IFN-γ in inducing these pathological effects.
IFN-γ has various effects on macrophages and other cell types, and induces NO and TNF-α production.39 As TNF-α contributes to local inflammatory reactions that are harmful to the host, we examined the possibility that IL-12 and IL-18 induce pathological changes via TNF-α. However, TNF-α was not detected in sera obtained daily after initiation of injection of IL-12 and IL-18 by ELISA (data not shown). Furthermore, TNF-α KO mice treated with IL-12 and IL-18 suffered weight loss (fig 1B) accompanied by intestinal inflammation (fig 5B, E) and fatty liver (fig 6C). Hence we conclude that TNF-α is not responsible for inducing this pathological state. It is well known that IFN-γ induces TNF-α production.40 Nevertheless, administration of IL-12 and IL-18 failed to induce TNF-α, even if high levels of IFN-γ were observed. It is intriguing to speculate that IL-18 is a negative regulator of TNF-α production induced by IFN-γ.
IL-12 and IL-18 caused marked increases in serum levels of IFN-γ and NO in BALB/c-wt mice (figs 2A, 3A). On day 3, peak increases in both occurred (figs 2A, 3A). NO production was IFN-γ dependent because no such increase occurred in IFN-γ KO mice treated with IL-12 and IL-18 (fig 3A). TNF-α was reported to induce NO production.41However, as noted above, TNF-α was not detected in mice treated with IL-12 and IL-18 for unknown reasons, leading to no NO production in IFN-γ KO mice. Changes in serum levels of IFN-γ were closely correlated with those of NO, in terms of both levels and time after stimulation with IL-12 and IL-18. There are at least two possibilities to account for this correlation. Diminished IFN-γ production may have resulted in diminished NO production. Alternatively, increased NO production may have inhibited IFN-γ production by T cells or natural killer cells, possibly by damaging these cells. Indeed, thymi of BALB/c-wt mice treated with IL-12 and IL-18 showed marked atrophy with cell apoptosis (data not shown), possibly leading to a reduced T cell supply from the thymus. NO is an important cytotoxic molecule in the defence against microorganisms,42 tumour cells,43 or other NO sensitive cells.44Furthermore, NO and/or its oxidised forms have the capacity to induce apoptosis in murine peritoneal macrophages42 and thymocytes45 by directly damaging DNA.46Interestingly, NO can also serve as an antioxidant in the presence of reactive oxygen species and protect the host from the cytotoxic action of reactive oxygens.47 As noted above, we showed that IL-12 and IL-18 induced an increase in serum levels of NO (fig 3). As NO is a cytotoxic molecule that induces cell apoptosis, we suspected that NO might have a crucial role in inducing the pathological state. But iNOS KO mice injected with IL-12 and IL-18 also manifested weight loss accompanied by intestinal inflammation (figs 1B; fig 5C, F), which indicates that NO did not have a crucial role in inducing the pathological state.
The mechanism of induction of severe fatty liver by IL-12 and IL-18 remains unknown. As injection of a mixture of IL-12 and IL-18 failed to induce fatty degeneration in the liver of IFN-γ KO mice (fig 6D), IFN-γ seems to play an important role. But in iNOS KO mice injected with IL-12 and IL-18, increased levels of IFN-γ were observed (fig2B) but there were no fatty changes in the liver (fig 6E). Hence NO may have a crucial role in inducing fatty degeneration.
Sequential administration of P acnes and LPS has been shown to induce severe liver injury.4 ,6 ,18 But LPS up to 10 μg/mouse/day did not cause such serious manifestations or severe histological changes (data not shown). Even if there was some leakage of LPS from the gut, it was of little importance in inducing the pathological changes observed in mice injected with IL-12 and IL-18.
In summary, we demonstrated the pathological role of IL-12 and IL-18 in inducing weight loss with intestinal inflammation and fatty liver. The pathological changes in the intestine were Th1 cells and IFN-γ dependent but TNF-α, NO, and Fas L independent. Fatty changes in the liver were NO dependent. Further study of the control of IL-18 production may provide a clinical approach for the treatment of disorders which are Th1 dependent.
Acknowledgments
We thank Mr K Takemura and Ms. T Takada for assistance in electron microscopy. This study was supported by a Grant-in-Aid for Scientific Research and Hitech Research Center grant from the Ministry of Education, Science and Culture of Japan and by CREST (Core Research for Evolutional Science and Technology) of Japan Science and Technology Corporation (JST).
Abbreviations used in this paper
- IFN-γ
- interferon γ
- IL
- interleukin
- LPS
- lipopolysaccharide
- L
- ligand
- wt
- wild type
- TNF-α KO
- tumour necrosis factor α knock out
- iNOS KO
- inducible nitric oxide synthase knock out
- NO
- nitric oxide
- PBS
- phosphate buffered saline
- ALT
- alanine transaminase