Article Text

Abstract
BACKGROUND AND AIMS Body gastritis caused by Helicobacter pylori infection appears to inhibit gastric acid secretion. The aim of this study was to determine the effects ofH pylori infection on gastric acid secretion and clarify its mechanisms with reference to interleukin 1β (IL-1β).
METHODS (1) Mongolian gerbils were inoculated orally with H pylori. Before, six, and 12 weeks after inoculation, serum gastrin levels, gastric acid output, and IL-1β mRNA levels in the gastric mucosa were determined. Pathological changes were also determined according to the updated Sydney system. (2) Effects of recombinant human IL-1 receptor antagonist (rhIL-1ra) on gastric acid output and serum gastrin levels were also determined.
RESULTS (1) Scores for activity and inflammation of gastritis and serum gastrin levels were significantly increased, and gastric acid output was significantly decreased six and 12 weeks after inoculation withH pylori. IL-1β mRNA levels in the gastric mucosa were also elevated six and 12 weeks after inoculation withH pylori. (2) Acid output and serum gastrin levels in the infected groups returned to control levels after rhIL-1ra injection.
CONCLUSIONS Gastric acid secretion is decreased and serum gastrin levels are increased in Mongolian gerbils infected with H pylori. This change in gastric acid secretion appears to be mediated by IL-1β induced by H pylori infection.
- Helicobacter pylori
- interleukin 1β
- Mongolian gerbils
- gastric acid
- gastrin
Abbreviations used in this paper
- IL-1β
- interleukin 1β
- rhIL-1ra
- recombinant human IL-1 receptor antagonist
- CFU
- colony forming units
- PCR
- polymerase chain reaction
Statistics from Altmetric.com
Helicobacter pylori colonises the human gastric mucosa and causes chronic gastritis and peptic ulceration in some subjects.1 ,2 The effects of H pylori infection on gastric acid secretion depend on several variables. It has been reported that basal and peak acid output, and plasma gastrin levels were significantly higher in patients withH pylori positive duodenal ulcers than inH pylori negative controls.3 ,4In some patients with H pylori related body gastritis, gastric acid output was low or absent, whereas acid secretion was improved and serum gastrin levels decreased significantly with eradication of H pylori.5In contrast, a large proportion of H pyloriinfected non-ulcer patients have no abnormality in gastrin release or gastric acid output.6 Recently, we reported that the degree of acid suppression depended on the distribution of theH pylori infection and on the degree of gastritis in the body, and serum gastrin levels increased in patients with duodenal and gastric ulcers.7 However, the mechanisms by which H pylori affects acid secretion are still unclear.
Interleukin 1β (IL-1β), which plays an important role in inflammatory and immunological responses,8 has been shown to inhibit gastric acid secretion in rats.9 ,10 Production of IL-1β in gastric mucosa was increased in patients withH pylori associated gastritis.11 ,12 Recently, El-Omar et al reported that IL-1 gene polymorphisms, in relation to enhanced production of IL-1β, were associated with an increased risk of hypochlorhydria induced by H pyloriinfection in gastric cancer relatives.13
It has been reported that Mongolian gerbils can be easily infected withH pylori, resulting in the occurrence of gastritis, ulcers, and neoplasms which mimic H pylori positive patients with gastric ulcer and gastric cancer.14 ,15 However, gastric acid secretion and IL-1β levels in the gastric mucosa of Mongolian gerbils have not been studied previously. Hence in this study we investigated the effects ofH pylori infection on gastric acid secretion and serum gastrin levels in Mongolian gerbils. In addition, we examined if IL-1β was associated with acid secretion in this animal model infected with H pylori.
Methods
ANIMALS
Six week old specific pathogen free male Mongolian gerbils (MGS/Sea), mean body weight 50 g, were purchased from Seiwa Experimental Animal Ltd (Fukuoka, Japan) and housed in polycarbonate cages in isolators. They were fed a commercial rodent diet and provided with water ad libitum. All animal experimentation was performed in accordance with institutional guidelines.
BACTERIAL STRAINS, MEDIA, AND GROWTH
A H pylori standard strain (ATCC43504) was obtained from American Type Culture Collection (Maryland, USA). This strain is cytotoxin associated protein positive and vacuolating cytotoxin positive. The bacteria were grown on Brucella broth (BBL; Becton Dickinson and Co., Cockeyville, Maryland, USA) containing 10% horse serum and incubated for 24 hours under microaerobic conditions at 37°C.
BACTERIAL INOCULATION
The culture of H pylori (0.5 ml, 1×108 colony forming units (CFU)) was administered orally, using a feeding needle, to Mongolian gerbils after fasting for 24 hours.16 Brucella broth without H pylori was given to the control groups. Four hours after inoculation, Mongolian gerbils were fed and housed in the same manner as the former for six and 12 weeks.
MEASUREMENT OF GASTRIC ACID OUTPUT AND SERUM GASTRIN LEVELS
Six and 12 weeks after inoculation, nine Mongolian gerbils infected with H pylori and nine control gerbils were operated on and studied. The pylorus was ligated under ether anaesthesia to trap gastric juice in the stomach. After operation for pylorus ligation, the gerbils woke up from anaesthesia and were housed in cages without food or water for three hours. Three hours later, Mongolian gerbils were sacrificed under ether anaesthesia. The stomach was immediately removed and opened along the greater curvature. Whole stomach weight was measured. The longitudinal half of the stomach was used for quantitative determination of H pyloriand the other half of the stomach was used for histological examination and extraction of RNA for analysis of IL-1β mRNA. Blood samples were obtained to determine serum gastrin levels. Gastric juice was collected, the volume was measured, and concentrations of acid were determined by titration with 0.1N NaOH to pH 7.0 on an automatic titrator (TOA Electronics Ltd., Tokyo, Japan). Serum gastrin levels were measured using a gastrin radioimmunoassay kit (Gastrin-RIA Kit II; Dainabot Co., Ltd, Tokyo, Japan) and the polyethylene glycol method. Antibodies used in this gastrin radioimmunoassay method strongly recognise the tyrosin residue at position 12 and the methionyl residue at position 15 on gastrin (1–17). As a result, intra- and interassay coefficients were 3.8–5.5% and 3.8–6.7%, respectively. Mean recovery of gastrin added to serum samples was 106 (12.2)%. Nine animals that were six week old were sacrificed beforeH pylori inoculation and acid output and serum gastrin levels measured as described above.
INJECTION OF INTERLEUKIN 1 RECEPTOR ANTAGONIST
Recombinant human IL-1 receptor antagonist (rhIL-1ra) was overexpressed in Escherichia coli as a 17 kd protein by cloning IL-1ra cDNA into a non-fused expressing vector provided by China Newtech Development and Trade Corporation (Beijing, China). Nine Mongolian gerbils infected with H pylori and nine control gerbils 12 weeks after inoculation were injected with rhIL-1ra intraperitoneally at a dose of 10 units/g, immediately after pylorus ligation. Three hours later they were sacrificed. Gastric acid output and serum gastrin levels were analysed as described above.
QUANTIFICATION OF VIABLE H PYLORI
After the stomach was opened along the greater curvature, the longitudinal half of the stomach was homogenised in 10 ml of phosphate buffered saline with a Polytron (Kinetica, Steinhofhalde, Sweden). Aliquots (100 μl) of the dilutions were applied to the plated agar media containing vancomycin (10 mg/l), polymyxin B (2500 IU/l), and amphotericin B (2 mg/l) for isolation of H pylori (Poamedia, Eikenkagaku Co., Ltd, Tokyo, Japan). The plates were incubated for seven days at 37°C under microaerobic conditions. Gram negative and oxidase, catalase, and urease positive spiral curved rods were identified as H pylori.The number of colonies was determined. The number of viableH pylori was expressed as CFU per stomach.
HISTOPATHOLOGY
Each stomach was fixed in 10% neutral buffered formalin. After fixation, a strip from the stomach, extending from the antrum to the body, was collected and embedded in paraffin. Strips were sectioned into 4 μm pieces and stained with haematoxylin-eosin and Giemsa.
In the present study, one pathologist semiquantitatively scored the strips blindly without knowledge of H pyloristatus or duration of infection. In each strip, microscopic investigations were performed to document histopathological changes. Polymorphonuclear neutrophil activity, chronic inflammation, glandular atrophy, intestinal metaplasia, and H pyloridensity in the gastric mucosa were graded 0, 1, 2, and 3 according to the updated Sydney system.17
SEQUENCE ANALYSIS OF MONGOLIAN GERBIL IL-1β GENE AND β-ACTIN GENE
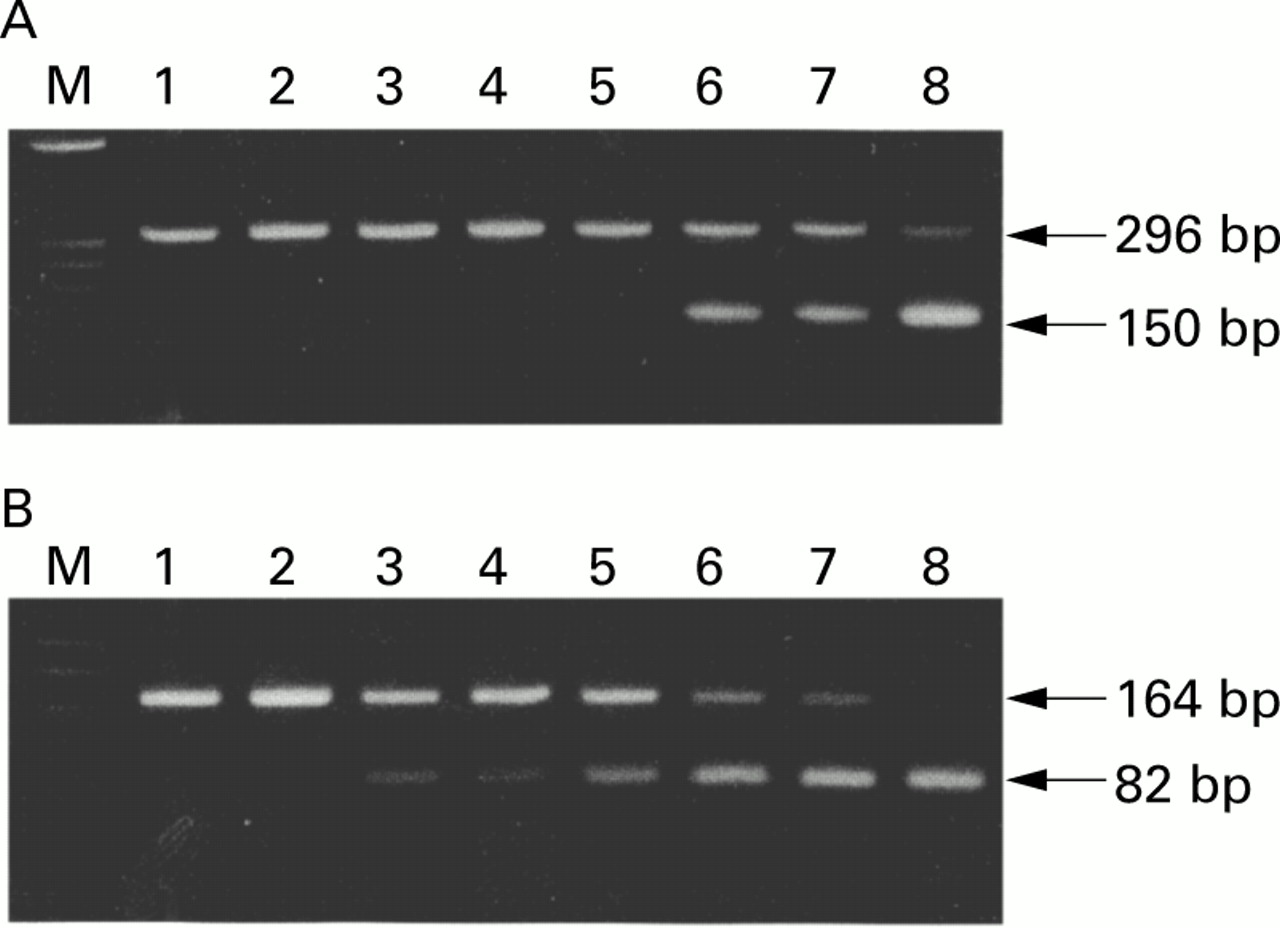
To identify Mongolian gerbil IL-1β complimentary DNA (cDNA) clones, we selected similar parts of the sequence between murine IL-1β cDNA and the related sequence of human IL-1β in the biologically active portion.18-20 We chose a forward primer (corresponding to human IL-1β positions 4445–4462 in exon 4) and a reverse primer (positions 6673–6691 in exon 6) for polymerase chain reaction (PCR) (fig 1), and we amplified a segment of approximately 296 bp from the cDNA in the gastric mucosa of Mongolian gerbils. The PCR product was cloned into pCRII using the original TA cloning kit (Invitrogen, San Diego, California, USA). The nucleotide sequence of the insert was determined by the dideoxy chain termination procedure.21

Oligonucleotide sequences of polymerase chain reaction (PCR) primers and competitive templates. The primer pair for Mongolian gerbil interleukin 1β (IL-1β) amplifies a 296 bp band. The primer pair for β-actin amplifies a 164 bp band. The competitive template for Mongolian gerbil IL-1β is 150 bp long, and that for β-actin is 82 bp long. PCR products of cDNA are distinguished by size from those of competitors.
To identify Mongolian gerbil β-actin cDNA clones, we chose one forward primer (corresponding to human β-actin positions 390–411) and one reverse primer (positions 973–993) (fig 1).22 We amplified a segment of approximately 164 bp from the cDNA and determined the sequence as mentioned above.
ANALYSIS OF IL-1β BY QUANTITATIVE REVERSE TRANSCRIPTION-POLYMERASE CHAIN REACTION
Total RNA was extracted from the gastric mucosa of the Mongolian gerbil soon after removing the stomach. RNA extraction was performed using an RNA extraction kit (RNA zoll; Tel-Test, Inc., Friendswood, Texas, USA). After DNase treatment, a 5 μg portion of total RNA solution was subjected to reverse transcription using 200 U of reverse transcriptase (Superscript Rnase H-Reverse Transcriptase; Gibco BRL, Life Technologies, Inc., Rockville, Maryland, USA).
The primer pair for Mongolian gerbil IL-1β (fig 1) amplifies the 296 bp band from the cDNA. For competitive PCR, we prepared a competitive template (competitor) containing the same primer binding and a subset of the same template sequences as the target competing for primer binding and amplification (fig 1). The PCR products are distinguished by size (cDNA 296 bp; competitor 150 bp). Nine precise 10-fold serial dilutions ranging from 1.0×103 pg/μl to 1.0×10-1 pg/μl of competitor template were prepared.
We prepared a master mix containing, in a final volume of 50 μl, Mongolian gerbil IL-1β forward primer and reverse primer (0.5 μM each as the final concentration), dNTPs (200 μl each as the final concentration), 5 μl of 10× PCR buffer, 1.25 units of Taq polymerase (Ex taq; Takara Shuzo, Shiga, Japan), and 2.0 μl of cDNA solution. An aliquot of 48 μl of this mixture was added to 2.0 μl of previously prepared competitive template of known concentration in several dilution series. PCR was performed with an automatic thermal cycler (DNA Thermal Cycler PJ2000; Perkin-Elmer, Norwalk, Conneticutt, USA). After heating at 94°C for five minutes, the samples were immediately cycled 35 times with a one minute denaturing step at 94°C, one minute primer annealing step at 55°C, and one minute extension step at 72°C. The final cycle included extension for eight minutes at 72°C to ensure full extension of the product.
The completed reactions were analysed by electrophoresis of a 10 μl aliquot through 3.0% (wt/vol) agarose gels containing 0.5 μg/ml ethidium bromide. The bands were visualised by excitation with UV light, and 296 bp (Mongolian gerbil IL-1β) and 150 bp (competitor) bands were detected (fig 2A). The gel was photographed, and the intensity of ethidium bromide luminescence was measured with a CCD image sensor (Densitograph AE-6900-F; Atto, Tokyo, Japan). The ratio of the 296 bp (cDNA) to the 150 bp (competitor) band was plotted for each dose of competitor added to the reaction tube. The point of equivalence (that is, where there was a 1:1 ratio) was where IL-1β equalled the competitor and represented the concentration of IL-1β mRNA in the unknown (fig 3).

(A) Mongolian gerbil interleukin 1β (IL-1β) (296 bp) versus competitor (150 bp). Lane M is the size marker (øX174 digested with Hae III). Lanes 1–8 contain eightfold serial amounts of competitor DNA ranging from 1.0×10-0.5 pg/μl to 1.0×103 pg/μl competing against a fixed dose of cDNA. (B) β-actin (164 bp) versus competitor (82 bp). Lane M is the size marker (øX174 digested with Hinf I). Lanes 1–8 contain eightfold serial amounts of competitor DNA ranging from 1.0×10-0.5pg/μl to 1.0×103 pg/μl competing against a fixed dose of cDNA.

(A) Logarithmic plot of the ratio of Mongolian gerbil interleukin 1β (IL-1β, 296 bp) to competitor (150 bp), corresponding to fig 2A. The quantity of IL-1β mRNA in the reaction tube was determined by calculating how much of the competitor was required to achieve equal amounts of product. The amount of IL-1β mRNA was then calculated by extrapolating from the intersection of the curves to the x axis where the amounts of IL-1β (296 bp) and competitor were equal. (B) Logarithmic plot of the ratio of β-actin (164 bp) to competitor (82 bp), corresponding to fig 2B.
The amount of β-actin mRNA was also measured by a similar method (fig2B). The sequences of oligonucleotide primers for β-actin and the competitive template are shown in fig 1.23 The amount of IL-1β mRNA in the gastric mucosa was standardised by β-actin and expressed as the ratio IL-1β/β-actin.
STATISTICAL ANALYSIS
All numerical data are expressed as mean (SEM). For statistical analysis, a Statview software package (Hulinks, Tokyo, Japan) for Macintosh (Apple Computer, Inc., Cupertino, California, USA) was used. The statistical significance of the differences in gastric acid output, serum gastrin levels, body weights, stomach weights, and gastric juice volume between the infected and control groups were determined by Student t test. Differences between values before, six, and 12 weeks after inoculation were determined by two way ANOVA with Fisher's multiple comparison test. Effects of rhIL-1ra on gastric acid output and serum gastrin levels in the infected and control groups were determined by one way ANOVA with Fisher's multiple comparison test. The statistical significance of the differences in gastritis scores between the infected and control groups were determined by the Mann-Whitney U test. Differences resulting in p values ⩽0.05 were considered significant.
Results
H PYLORI INFECTION, BODY WEIGHTS, AND STOMACH WEIGHTS OF MONGOLIAN GERBILS
H pylori was detected in all infected animals throughout the study. The number of viableH pylori in the stomach reached 105.0 CFU/gastric wall at six weeks and remained at 105.1 CFU at 12 weeks (table 1). Body weights of Mongolian gerbils time dependently increased from before to six and 12 weeks after inoculation but there were no significant differences between the infected and control groups (table1).
Changes in body weight, stomach weight, and gastric juice volume as a function of Helicobacter pylori (HP) infection
Stomach weights of Mongolian gerbils at six and 12 weeks after inoculation significantly increased compared with those before inoculation, and stomach weights of the infected groups were significantly heavier than those of the control groups, but no significant changes were observed from six to 12 weeks after inoculation (table 1).
HISTOPATHOLOGY
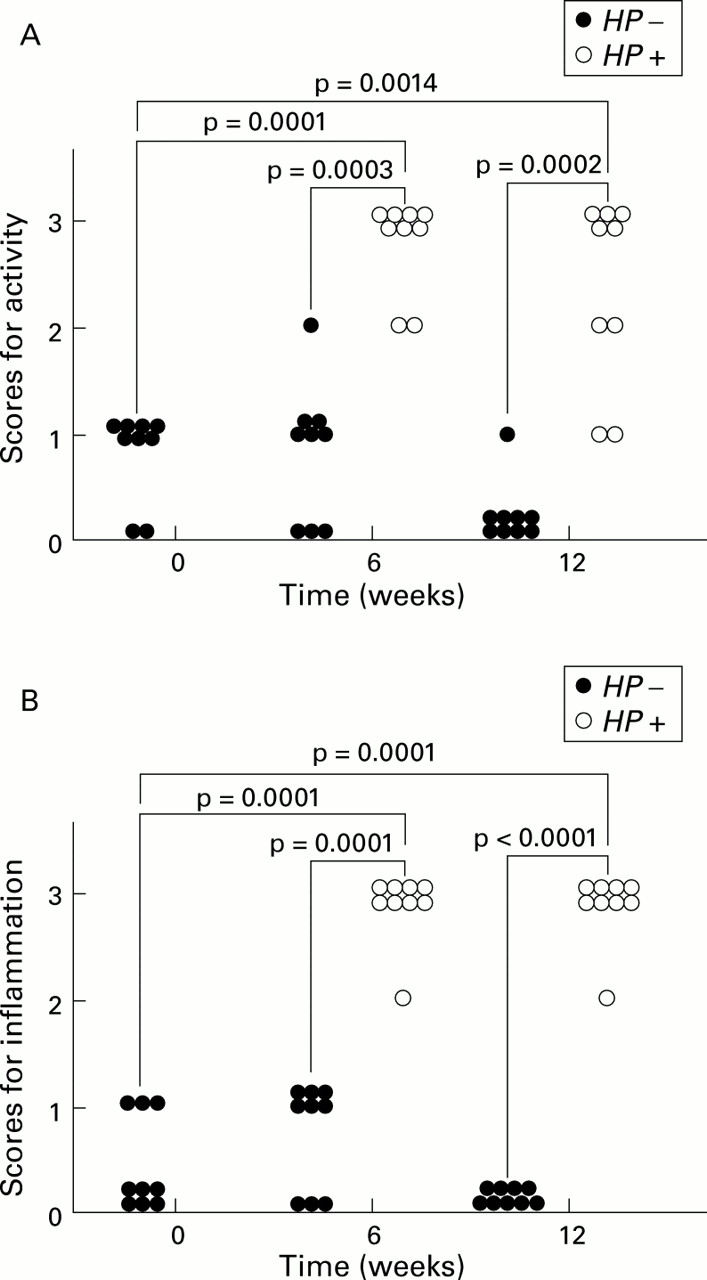
Marked infiltration of neutrophils and lymphocytes, mainly in the submucosa, spreading from the antrum to the body and focal aggregates of lymphocytes in the submucosa at the glandular border of the antrum and body, were observed in Mongolian gerbils infected withH pylori, at both six and 12 weeks after inoculation. In contrast, infiltrations of neutrophils and lymphocytes were few or none in the control groups (fig 4). Scores for activity were significantly higher in the infected groups than in the control groups at both six (p<0.001) and 12 (p<0.001) weeks after inoculation. Scores for activity six and 12 weeks afterH pylori inoculation were significantly increased compared with those before inoculation (p=0.0001, p<0.01) (fig 5A). Scores for inflammation were significantly higher in the infected groups than those of the control groups both six (p=0.0001) and 12 (p<0.0001) weeks after inoculation. Scores for inflammation six and 12 weeks after H pylori inoculation were significantly increased compared with those before inoculation (p=0.0001, p=0.0001) (fig 5B). No significant changes in glandular atrophy were observed between the infected and control groups at both six and 12 weeks after inoculation. There was a tendency for detectable colonies of H pylori to be accompanied by both neutrophil and lymphocytic infiltration (data not shown). Slight goblet cell metaplasia was detected only in a few cases in the infected groups 12 weeks after inoculation (data not shown).

Histopathological changes in the gastric mucosa of Mongolian gerbils. (A) Inflammatory cell infiltrations were not observed in the control groups six weeks after inoculation. (B) Marked inflammatory cell infiltrations were observed in the infected groups six weeks after inoculation with H pylori. (C) Inflammatory cell infiltrations were not observed in the control groups 12 weeks after inoculation. (D) Severe inflammatory cell infiltrations were observed in the infected groups 12 weeks after inoculation with H pylori (haematoxylin-eosin, ×100).

(A) Changes in gastritis scores for activity from before to six and 12 weeks after inoculation in the infected (Helicobacter pylori (HP+)) and control (HP−) groups. The variables used for grading gastritis were rated on a four point scale: 0, absent; 1, mild; 2, moderate; and 3, severe, in accordance with the Sydney system. Gastritis scores for activity in the infected groups significantly increased six and 12 weeks after H pylori inoculation. No significant changes were observed in the control groups. (B) Changes in gastritis scores for inflammation from before to six and 12 weeks after inoculation in the infected and control groups. Gastritis scores for inflammation in the infected groups significantly increased six and 12 weeks after H pylori inoculation. No significant changes were observed in the control groups.
GASTRIC JUICE VOLUME AND GASTRIC ACID OUTPUT
The volume of gastric juice six and 12 weeks after inoculation was significantly increased compared with that before inoculation. Gastric juice volume was significantly greater in the infected groups than in the control groups (table 1).
Gastric acid output six and 12 weeks after H pylori inoculation in the infected groups was significantly decreased compared with that before inoculation (29.6 (9.1), 22.5 (4.3)v 51.5 (6.2) μEq/3 hours; p<0.05, p<0.01). Gastric acid output was significantly lower in the infected groups than in the control groups six weeks (29.6 (9.1)v 61.1 (9.8) μEq/3 hours; p<0.05) and 12 weeks (22.5 (4.3) v 44.1 (5.9) μEq/3 hours; p<0.01) after inoculation. No significant changes in gastric acid output were observed from before to six or 12 weeks in the control groups (fig 6).

Changes in gastric acid output from before to six and 12 weeks after inoculation in the infected (Helicobacter pylori (HP+)) and control (HP−) groups. Gastric acid output in the infected groups significantly decreased six and 12 weeks after H pylori inoculation. No significant changes were observed in the control groups. *p=0.0328; **p=0.0061 compared with before inoculation.
SERUM GASTRIN LEVELS
Serum gastrin levels six and 12 weeks afterH pylori inoculation in the infected groups were significantly increased compared with those before inoculation (153.9 (13.4), 157.3 (10.2) v 100.4 (6.7) pg/ml; p<0.01, p<0.001). Serum gastrin levels were significantly higher in the infected groups than in the control groups six weeks (153.9 (13.4) v 112.9 (9.2) pg/ml; p<0.05) and 12 weeks (157.3 (10.2) v 118.4 (3.4) pg/ml; p<0.01) after inoculation.
No significant changes in serum gastrin levels were observed in the control groups from before to six and 12 weeks after inoculation (fig 7).

Changes in serum gastrin levels from before to 6 and 12 weeks after inoculation. Serum gastrin levels in the infected (Helicobacter pylori (HP+)) groups significantly increased six and 12 weeks after H pylori inoculation. No significant changes were observed in the control groups (HP−). *p=0.0014; **p=0.0008 compared with before innoculation.
SEQUENCE ANALYSIS OF MONGOLIAN GERBIL IL-1β GENE AND β-ACTIN GENE
Sequences of the 296 bp portion of IL-1β cDNA were aligned (fig8A). The nucleotide sequences of Mongolian gerbil and human IL-1β were closely related (252 of 296 (85.1%)), and those of the Mongolian gerbil and murine IL-1β were also closely related (267 of 296 (90.2%)). We also aligned Mongolian gerbil, human, and murine β-actin (fig 8B). The nucleotide sequences of Mongolian gerbil and human β-actin were closely related (159/164: 97.0%), and those of murine β-actin were also closely related to Mongolian gerbils (158/164: 96.3%).

(A) Alignment of the 296 bp portion of interleukin 1β (IL-1β) cDNA sequences for the Mongolian gerbil, human, and mouse. Oligonucleotides in bold type in human and mouse are different from those of Mongolian gerbil; 85.1% oligonucleotide identities were observed between Mongolian gerbil and human and 90.2% between Mongolian gerbil and mouse. (B) Alignment of the 164 bp portion of β-actin cDNA sequences for the three species. Oligonucleotides in bold type in human and mouse were different from those of the Mongolian gerbil; 97.0% oligonucleotide identities were observed between Mongolian gerbil and human and 96.3% between Mongolian gerbil and mouse.
IL-1β MRNA LEVELS
IL-1β mRNA levels six and 12 weeks after H pylori inoculation in the infected groups were significantly increased compared with those before inoculation (22.4 (8.6), 26.5 (6.2) v 0.0 (0.0); p<0.05, p<0.01). IL-1β mRNA levels in the gastric mucosa were significantly higher in the infected groups than in the control groups both six and 12 weeks after inoculation (22.4 (8.6) v 0.06 (0.06) (p<0.05) and 26.5 (6.2) v 0.02 (0.02) (p<0.001)) (fig 9).

Changes in interleukin 1β (IL-1β) mRNA levels from before to six and 12 weeks after inoculation in the infected (Helicobacter pylori (HP+)) and control (HP−) groups. Il-1β mRNA levels significantly increased six and 12 weeks after H pylori inoculation. IL-1β mRNA was virtually undetected in the control groups throughout the study. *p=0.0191; **p=0.0006 compared with before innoculation.
EFFECTS OF RECOMBINANT HUMAN INTERLEUKIN 1 RECEPTOR ANTAGONIST (rhIL-1ra) ON GASTRIC ACID OUTPUT AND SERUM GASTRIN LEVELS
Gastric acid output in Mongolian gerbils infected with H pylori after rhIL-1ra injection significantly increased to control levels compared with the infected groups who did not receive rhIL-1ra injection (45.1 (7.3)v 22.5 (4.3) μEq/3 hours; p<0.05), and was not significantly different from the control groups. No significant effects of rhIL-1ra on gastric acid output were observed in Mongolian gerbils not infected with H pylori (54.1 (10.7) v 44.1 (5.9) μEq/3 hours) (fig10).

Gastric acid output with (+) or without (−) recombinant human IL-1 receptor antagonist (rhIL-1ra) injection in the infected (Helicobacter pylori (HP+)) and control (HP−) groups. Gastric acid output significantly increased to control levels after rhIL-1ra injection in the infected groups whereas no significant changes were observed after rhIL-1ra injection in the control groups.
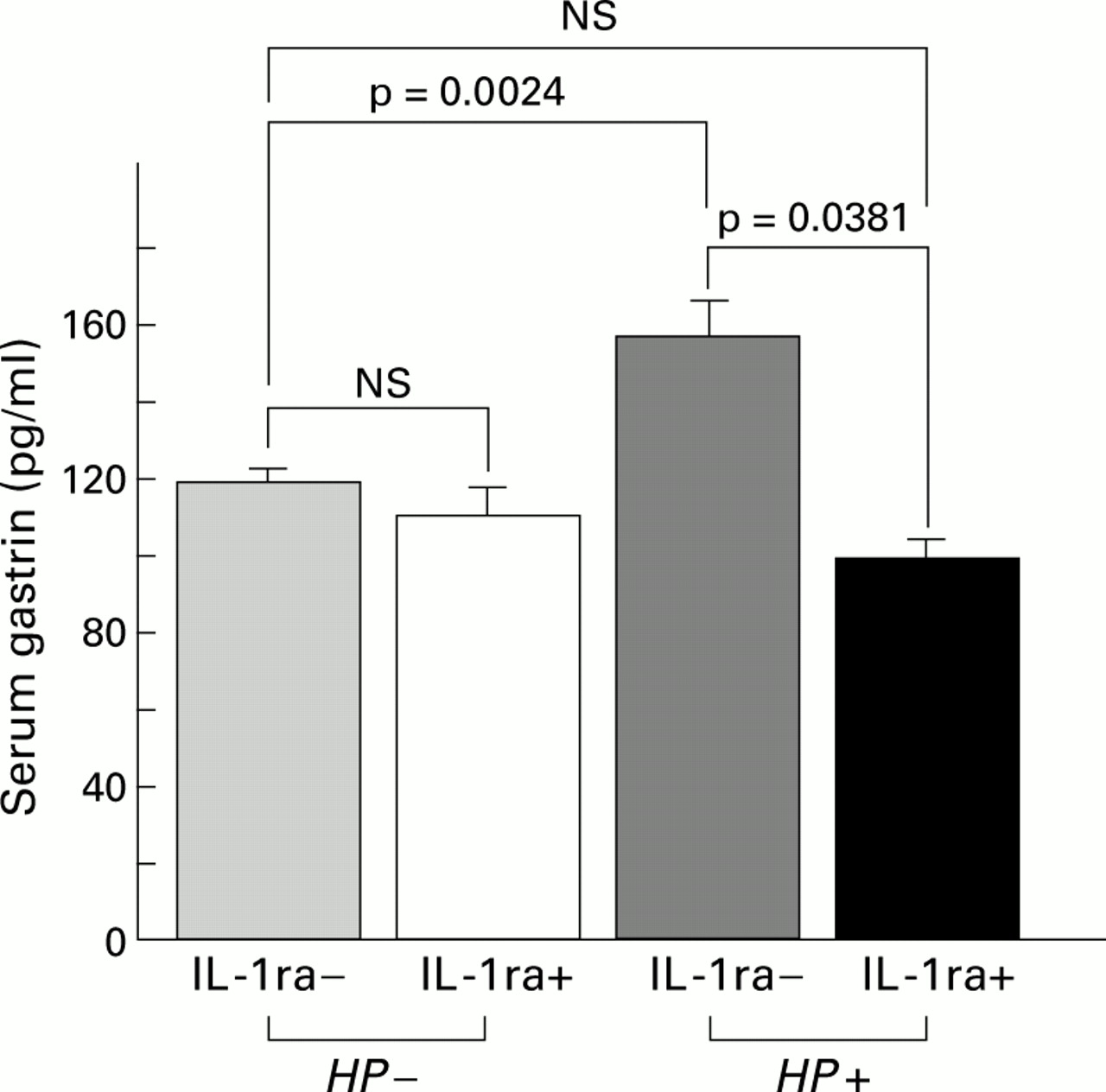
Serum gastrin levels in the Mongolian gerbils infected withH pylori after rhIL-1ra injection significantly decreased to control levels compared with the infected groups without rhIL-1ra injection (97.9 (5.8)v 157.3 (10.2) pg/ml; p<0.0001), and were not significantly different from the control groups. No significant effects of rhIL-1ra on serum gastrin levels were observed in Mongolian gerbils without H pylori infection (111.2 (7.5) v 118.4 (3.4) pg/ml) (fig11).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Serum gastrin levels with (+) or without (−) recombinant human IL-1 receptor antagonist (rhIL-1ra) injection in the infected (Helicobacter pylori (HP+)) and control (HP−) groups. Serum gastrin levels significantly decreased to control levels after rhIL-1ra injection in the infected groups whereas no significant changes were observed after rhIL-1ra injection in the control groups.
Discussion
In the present study, we determined changes in gastric acid output after H pylori infection and showed thatH pylori infection caused chronic active gastritis and suppressed gastric acid secretion, which was assumed to be mediated by IL-1β.
Recent studies revealed that gastric acid hyposecretion observed in patients with body gastritis recovered soon after eradication ofH pylori as inflammation was improved, even though gastric atrophy and intestinal metaplasia had not improved significantly.5 ,7 ,24 ,25 Inflammation of the gastric mucosa caused by H pylori infection may be closely related to gastric acid hyposecretion rather than atrophy. In the present study, severity of gastritis increased after H pylori infection but no significant changes in glandular atrophy in the gastric mucosa of Mongolian gerbils with or without H pylori infection were observed within the 12 week period of the present study. Therefore, gastric acid hyposecretion may not depend on glandular atrophy but on the severity of gastritis, at least for the duration of our study period.
H pylori itself and several substances such as N α-methyl histamine produced by H pylori and cytokines induced by H pylori infection, such as IL-1β and tumour necrosis factor α, have been reported to inhibit gastric acid secretion.26-29 In particular, IL-1β is known as a potent inhibitor of acid secretion; peripheral and central administration of IL-1β potently inhibits gastric acid secretion in experimental animals.9 ,10 ,29-31 IL-1β also directly inhibits acid secretion by cultured rabbit parietal cells,32 and impairs the function of enterochromaffin-like cells in vitro, which may be followed by markedly decreased acid output.33 Moreover, IL-1β mRNA levels and IL-1β release significantly increased in the gastric mucosa ofH pylori positive patients.34 ,35 In the present study, IL-1β mRNA levels in the gastric mucosa of Mongolian gerbils infected withH pylori were elevated in comparison with those of the control groups. In addition, gastric acid secretion in the infected groups was decreased in comparison with those of the control groups and recovered to control levels by rhIL-1ra. Therefore, IL-1β induced by H pylori infection is assumed to play an important role in gastric acid hyposecretion observed in body gastritis patients.
In the present study, marked infiltration of inflammatory cells, such as polymorphonuclear and mononuclear cells, in the gastric mucosa spreading from the antrum to the body and formation of lymphoid follicles in the submucosa in the transitional zone between the antrum and body were observed in Mongolian gerbils infected with H pylori. These histological findings were similar to the gastric mucosa of patients withH pylori positive gastritis.36 ,37 IL-1β is released mainly from activated monocytes and macrophages.8 Therefore, infiltration and activation of monocytes and macrophages induced byH pylori infection release IL-1β, resulting in gastric acid hyposecretion.
IL-1 receptor antagonist binds to the IL-1 receptor and blocks the binding of both IL-1α and IL-1β without inducing a signal of its own.8 The inhibitory effect of IL-1β on gastric acid secretion is suppressed by peripheral rhIL-1ra injection in rat.31 ,38 rhIL-1ra has been used as a potent blocker of IL-1 receptors of Mongolian gerbils in a previous report.39 Human, rat, and mouse IL-1β genes have similar sequences.40 In the present study, we identified part of the gene sequence of the biologically active portion of Mongolian gerbil IL-1β. There was a high level of conservation between human and Mongolian gerbil sequences. In addition, human, rat, and mouse IL-1ra genes have similar sequences.40 Therefore, rhIL-1ra was assumed to work as an antagonist at the IL-1β receptor of Mongolian gerbils. As demonstrated in our study, gastric acid hyposecretion in the infected groups returned to control levels after injection of rhIL-1ra. These results suggest that IL-1β induced byH pylori related body gastritis inhibits gastric acid hyposecretion.
In the present study, we showed that H pylori infection decreased gastric acid secretion but increased the volume of gastric juices. Therefore, secretion of some acid neutralising factors, such as bicarbonate and mucus, was assumed to be increased. A recent study has shown that IL-1β increases prostaglandin production.41 Prostaglandins increase bicarbonate and mucus secretion and inhibit basal and gastrin stimulated acid secretion.42 ,43 Therefore, an increase in the volume of gastric juices in spite of decreased acid secretion may be mediated by prostaglandins, which are induced by IL-1β.
We also determined serum gastrin levels in Mongolian gerbils infected with H pylori with reference to IL-1β. We showed that serum gastrin levels increased after H pylori infection. Many possible factors associated with hypergastrinaemia in H pylori infection have been reported. For example, the inhibitory effects of cholecystokinin on gastrin release were reduced by H pyloriinfection.44 Luminal ammonia elevated plasma gastrin concentrations in rats.45 Cytokines, such as IL-1β, IL-8, tumour necrosis factor α, and interferon γ release gastrin from rabbit, canine, and human antral G cells in primary culture.46-48 Intragastric hypoacidity may induce hypergastrinaemia.49 In the present study, serum gastrin levels and mucosal IL-1β levels significantly increased afterH pylori infection. After rhIL-1ra injection, serum gastrin levels decreased to control levels with an increase in gastric acid output. These findings highlight two reasons for hypergastrinaemia: a direct effect of IL-1β on gastrin release from G cells or gastrin release corresponding to a decrease in intragastric acidity. The inhibitory effect of low pH on the release of gastrin was observed in H pylori positive patients with and without duodenal ulcer disease, as well as inH pylori negative patients.50In the present study, after injection of rhIL-1ra, serum gastrin levels were decreased and gastric acid secretion was increased, although gastrin is a stimulant of acid secretion. Therefore, it is reasonable that IL-1β inhibits gastric acid secretion resulting in hypergastrinaemia in compensation for decreased acid secretion, and direct effects of IL-1β on gastrin release from G cells appear to be of limited relevance in vivo.
In conclusion, we have confirmed that H pylori infection causes gastritis from the antrum to the body which induces IL-1β in the gastric mucosa, which then inhibits gastric acid secretion resulting in hypergastrinaemia. Certainly, atrophy will spread out into the oxyntic mucosa, and parietal cell mass will decrease markedly, which contributes to gastric acid hyposecretion if the study period becomes longer. We also identified part of the sequence of a Mongolian gerbil IL-1β gene and determined mRNA levels of IL-1β in the gastric mucosa. Although we did not determine IL-1β release, it is considered that IL-1β levels correspond to mRNA levels.51 Therefore, IL-1β induced byH pylori infection is assumed to inhibit gastric acid secretion.
Lastly, our results should be interpreted within the context of the limitations of our study regarding the use of Mongolian gerbils for the following reasons: when Mongolian gerbils were infected withH pylori, the transitional zone between the antrum and corpus of the stomach was reported to gradually expand up into the fundic mucosa as the infection progressed.52 This was similar to the phenomenon observed in patients with body gastritis. However, the number of viable H pyloridetected in the gastric mucosa of Mongolian gerbils was much less than that of humans infected with H pylori andH pylori (Sydney strain) infected mice.14 ,53 Differences in sensitivity toH pylori may account for the differences in numbers.53 Moreover, basal gastric acid output of Mongolian gerbils was low and approximately 1/15th of the output in rats.53 This lower basal acid secretion of Mongolian gerbils may explain why duodenal ulcer is not observed in thisH pylori infected animal model, in spite of many reports of development of gastric ulcer and gastric cancer.15 ,16 ,52 ,53
Therefore, we consider this animal model is appropriate for the study of disorders with hyposecretion caused by H pylori infection. Our study will be helpful to clarify the mechanisms by which body gastritis caused by H pylori infection inhibits gastric acid secretion, which is now considered to be closely related to the development of gastric cancer.13
Acknowledgments
The study was supported by a Grant-in Aid for Scientific Research from the Ministry of Education, Science, Sports, and Culture, Japan. The abstract of the study was presented at the annual meeting of the AGA, San Diego, California, USA, May 2000. We thank Fumihiro Hirayama for technical advice.
Abbreviations used in this paper
- IL-1β
- interleukin 1β
- rhIL-1ra
- recombinant human IL-1 receptor antagonist
- CFU
- colony forming units
- PCR
- polymerase chain reaction