Article Text

Abstract
BACKGROUND Lamina propria (LPLs) and intraepithelial (IELs) lymphocytes are markedly increased in coeliac mucosa, and are thought to play a crucial role in the generation of villous atrophy in coeliac disease (CD). However, the mechanisms by which they mediate the killing of enterocytes in this condition are still poorly characterised.
AIM We investigated Fas mediated cytotoxicity and apoptosis of both LPLs and IELs, isolated from 10 untreated coeliac patients, 10 coeliac patients on a gluten free diet, and 10 biopsied controls.
METHODS Fas and Fas ligand expression were assessed by flow cytometry and immunocytochemistry. Lymphocyte cytotoxicity against Fas expressing Jurkat cells was determined by the Jam test. The effect of the antagonist ZB4 anti-Fas antibody on apoptotic activity exerted by coeliac lymphocytes against enterocytes was analysed. Lymphocyte apoptosis was assessed by oligonucleosome ELISA.
RESULTS LPLs and IELs showed increased apoptotic activity and higher levels of Fas ligand expression in untreated CD compared with treated CD patients and controls. Enterocyte apoptosis observed after coculturing coeliac lymphocytes and enterocytes in the presence of ZB4 antibody was reduced. In active CD, LPLs manifested increased apoptosis whereas IELs showed decreased apoptosis.
CONCLUSIONS Our results support the involvement of the Fas/Fas ligand system in CD associated enterocyte apoptosis. Increased LPL apoptosis is likely to downregulate mucosal inflammation whereas decreased IEL apoptosis could be responsible for autoimmune and malignant complications of CD.
- apoptosis
- coeliac disease
- cytotoxicity assay
- Fas/Fas ligand system
- intraepithelial lymphocytes
- lamina propria lymphocytes
Abbreviations used in this paper
- CD
- coeliac disease
- FasL
- Fas ligand
- IELs
- intraepithelial lymphocytes
- LPLs
- lamina propria lymphocytes
- PBS
- phosphate buffered saline
- FCS
- fetal calf serum
- TUNEL
- terminal deoxynucleotidyl transferase mediated digoxigenin-deoxyuridine triphosphate nick end labelling
Statistics from Altmetric.com
- apoptosis
- coeliac disease
- cytotoxicity assay
- Fas/Fas ligand system
- intraepithelial lymphocytes
- lamina propria lymphocytes
Coeliac disease (CD) is a T cell mediated enteropathy characterised by villous atrophy suggested to be due to increased enterocyte apoptosis,1 ,2 and by a high density of lamina propria (LPLs) and intraepithelial (IELs) lymphocytes.3Activated T LPLs are markedly increased in coeliac mucosa4and their crucial role in inducing mucosal flattening is well recognised.5 IELs have been shown to mediate Fas ligand (FasL) cytotoxicity against Fas expressing enterocytes in a murine model of graft versus host disease,6 ,7 a condition which shares many pathological and immunological similarities with CD. Moreover, in a recent study8 we have shown that in untreated CD, increased IEL FasL expression correlated significantly with the level of enterocyte apoptosis.
Thus to demonstrate effective involvement of LPLs and IELs in determining increased enterocyte apoptosis in CD, we evaluated the Fas/FasL cytotoxic mechanism9 in this condition. In addition, as lymphocyte apoptosis may contribute to limit expansion of activated T cells,10 and to delete autoreactive T cell clones,11 we also investigated LPL and IEL apoptosis, as dysregulation of programmed cell death of these cells may favour the autoimmune and malignant complications of CD.
Patients and methods
PATIENTS AND TISSUES
Size appropriate and well oriented endoscopic biopsy specimens (12–15 per patient) were obtained from the second part of the duodenum in 10 untreated coeliac patients (mean age 37.3 years, range 23–61), 10 coeliac patients on a gluten free diet (mean age 34.8 years, range 20–65) for at least 12 months, and 10 consenting subjects (mean age 39.2 years, range 28–62) undergoing upper gastrointestinal endoscopy for functional dyspepsia. Subjects using steroidal or non-steroidal anti-inflammatory drugs or presenting with an inflamed mucosa at histology were excluded from the study. Some of the biopsy samples were processed according to standard methods for traditional histology and others were used to obtain purified LPL, IEL, and enterocyte suspensions.
CELL ISOLATION
Biopsy specimens were collected in sterile medium (phosphate buffered saline (PBS) calcium and magnesium free, supplemented with 100 U/ml penicillin, 100 μg/ml streptomycin, and 5% fetal calf serum (FCS)). The epithelial layer was removed with 1 mM ethylenediamine-tetraacetic acid (EDTA; Sigma Chemical Co, St Louis, Missouri, USA) and 1 mM dithiothreitol (Sigma). After continuous agitation for one hour at 37°C, the single cell suspension was pelleted from the supernatant and washed once with 5 ml of RPMI 1640 medium (Gibco, Life Technologies Ltd, Paisley, UK) supplemented with antibiotics and 10% FCS. IELs and enterocytes were then separated on a Percoll density gradient (Pharmacia Fine Chemicals, Pharmacia, Inc., Uppsala, Sweden). A discontinuous density gradient (25%, 40%, and 75%) was used. Cells that layered between the 40% and 75% fractions were collected as IELs whereas cells that layered between the 40% and 25% interface were collected as enterocytes. To release lamina propria mononuclear cells, the remaining tissue was washed three times with RPMI 1640 medium and treated with type 1A collagenase 128 U/ml (Sigma) for three hours with continuous agitation at 37°C. The crude cell suspension was allowed to stand for five minutes to permit sedimentation of debris. Cells from the supernatant were washed twice with RPMI 1640 medium containing 10% FCS, 100 U/ml penicillin, and 100 μg/ml streptomycin, and layered on Ficoll-Hypaque solution (Pharmacia) to isolate LPLs by a density gradient method. Single cell suspensions were resuspended in 1 ml of PBS and kept on ice until used. The viability of single cell suspensions was determined by trypan blue exclusion. Cells were not used if viability did not exceed 90%. The purity of enterocyte and IEL populations, determined by processing cell cytocentrifugates with anticytokeratin pan antibody at 1:400 dilution (Boehringer Mannheim, Indianapolis, Indiana, USA), exceeded 95% and 80%, respectively. The purity of the LPL population, assessed by flow cytometric analysis on a FACS II analyser (FACScan; Becton Dickinson Co., San Jose, California, USA), was 90%. Purified enterocytes as targets and both LPLs and IELs as effectors were used in the cytotoxicity assay.
IMMUNOFLUORESCENCE AND FLOW CYTOMETRY
Freshly isolated LPLs were washed twice in PBS containing 2% FCS and incubated at 4°C for 30 minutes with phycoerythrine labelled anti-Fas mouse MoAb IgG1 (Coulter/Immunotech, Inc., Westbrook, Minnesota, USA) and fluorescein isothiocyanate labelled anti-FasL mouse MoAb IgG2 (Upstate Biotechnology, Lake Placid, New York, USA). Based on forward and side scatter, cells were gated on the area corresponding to lymphocytes and analysed by a FACScan (Becton Dickinson). Fluorescein isothiocyanate labelled IgG1 and phycoerythrine labelled IgG2 (Becton Dickinson) were used as control reagents to exclude non-specific binding and to determine the optimal setting of fluorescence quadrants.
IMMUNOCYTOCHEMISTRY
Isolated IELs were placed on electrostatic slides by means of cytocentrifugation, fixed by incubation for 10 minutes in cold acetone/methanol 1:1, and air dried. Afterwards, cells were incubated for five minutes in 3% hydrogen peroxide to quench endogenous peroxidase activity, rinsed in PBS, and incubated with bovine serum albumin 2% in PBS for 30 minutes. Subsequently IELs were incubated for 60 minutes at room temperature with the antihuman FasL IgG1 mouse monoclonal antibody (clone NOK-1; Pharmingen, San Diego, California, USA) at 1:150 dilution. Cells were rinsed in PBS and treated with a secondary biotinylated antibody and peroxidase conjugated streptavidin. Finally, diaminobenzidine reaction and nuclear counterstaining with Harris' haematoxylin were performed. As a positive control, the TM4 cell line (immortalised mouse Sertoli cells) was used. As a negative control, slides of IELs were processed without the primary antibody. Sections were examined using conventional light microscopy in a blinded fashion by an expert observer. Counts were performed at a constant magnification (×1000) by a differential count of at least 500 cells and the results expressed as mean percentage.
JAM TEST
To investigate whether FasL expressed on LPLs and IELs was functional, we analysed the ability of LPLs and IELs to kill Fas expressing Jurkat cells. DNA fragmentation in Fas+ target cells (human T cell leukaemia, Jurkat) after four hours of coculture with LPLs or IELs was performed as previously described.12Briefly, Jurkat cells (2×104) were pulsed for 24 hours with 2 μCi/ml of 3H thymidine (6.7 Ci/mmol; NEN, Boston, Massachusetts, USA), washed, and then incubated either alone or with effector cells at different effector/target ratios (5:1, 2.5:1, 1.25:1, and 0.625:1 for LPLs; and 3:1, 1.5:1, and 0.625:1 for IELs). At the end of the culture, cells were harvested on a glass fibre filter, and3H thymidine present on the filters was measured by liquid scintillation counting. The results are expressed as mean (SD) cpm of five replicate cultures. The percentage of fragmented DNA was calculated as follows: 100×(mean cpm target cells cultured alone−mean cpm effector cells cultured with target cells/mean cpm target cells cultured alone).
CYTOTOXICITY ASSAY
To determine if enterocyte apoptosis in CD involves interaction between Fas receptor expressed on target cells (enterocytes) and FasL expressed on effector cells (LPLs or IELs), the effect of an antagonist ZB4 anti-Fas antibody (Ancell Corporation, Bayport, Minnesota, USA) was analysed. LPLs and IELs (effector cells) and enterocytes (target cells), isolated from both untreated coeliac patients and biopsied controls, were incubated at 37°C in a 5% CO2 atmosphere for four hours in the absence or presence of 2 μg/ml ZB4 anti-Fas blocking added to the culture medium. The effector/target ratios were 100:1 in experiments involving LPLs and 20:1 in those involving IELs. Aliquots of both untreated coeliac and biopsied control enterocytes were cultured without effector cells to give an estimation of spontaneous enterocyte apoptosis either in the absence or presence of ZB4. Afterwards, cocultured cells were placed on electrostatic slides by means of cytocentrifugation, fixed by incubation for 10 minutes in paraformaldehyde 3.7%, air dried, and permeated by means of acetone/methanol 1:1. Finally, cells were processed for terminal deoxynucleotidyl transferase mediated digoxigenin-deoxyuridine triphosphate nick end labelling (TUNEL) detection using the peroxidase ApopTag Kit (Oncor, Gaithesburg, Maryland, USA) and observed under light microscopy to evaluate the percentage of apoptotic enterocytes by a differential count of at least 300 cells at a constant magnification (×1000).
OLIGONUCLEOSOME ELISA
Qualitative and quantitative in vitro determination of cytoplasmic histone associated DNA fragments (mono and oligonucleosomes) from LPLs and IELs were performed with the Cell Death Detection ELISA (Boehringer Mannheim).
MUCOSAL MORPHOMETRY
To quantify the persistence of mucosal abnormality in treated CD, we measured the surface area to volume ratio (S/V) of duodenal mucosa. S/V in biopsy specimens was estimated by a Weibel graticule (Graticules Ltd, Tunbridge Wells, UK) inserted in the microscope eyepiece, according to the method of Dunnil and Whitehead.13 A record was made of the number of times the lines cut the mucosal surface (c) and the number of hits (h)—that is, end points of the lines falling on the lamina propria. At a constant magnification (×125) the length (l) of each line of the graticule was 1.7×10−2 cm. The ratio c/lh was then calculated and regarded as a measure of S/V.
Those treated patients with an S/V ratio lower than 57 were considered to have some persistent degree of mucosal abnormality, according to our previous study.14
STATISTICAL ANALYSIS
Data are expressed as median (range). Statistical comparisons were carried out using the Mann-Whitney U test for non-parametric data. A level of p<0.05 was considered statistically significant.
Results
FasL EXPRESSION BY LPLs AND IELs
Flow cytometry was used to analyse expression of FasL on the plasma membrane of LPLs. Otherwise, as the number of viable isolated IELs (mean 1.5×106, range 0.5–2.3) was insufficient to perform all of the experiments, immunocytochemistry instead of flow cytometric analysis was employed to evaluate FasL expression by these cells.
Figure 1 shows flow cytometric analysis of FasL expression by LPLs isolated from biopsy specimens obtained from untreated coeliac patients, treated coeliac patients, and biopsied controls. In untreated CD, the percentage of FasL+ LPLs (median 39.3%, range 10.3–61.4) was significantly higher compared with treated CD (median 2.6%, range 0.2–10.7) and controls (median 0.5%, range 0.1–1.7). In treated CD, the percentage of FasL+ LPLs was still significantly higher than in controls.

Percentages of FasL+ lamina propria lymphocytes (LPLs) in 10 untreated coeliac patients compared with those of 10 treated coeliac patients and 10 biopsied controls. Those coeliac patients on a gluten free diet and with a persistent degree of villous atrophy are indicated with a different symbol (solid square ). Horizontal bars represent median values.
Figure 2 shows that in untreated CD almost all IELs were FasL+ and their percentage (median 80.8%, range 75.4–92.3) was significantly higher than in treated patients (median 27.5%, range 18.2–63.5) and controls (median 5.8%, range 1.2–10.9). In treated CD, FasL+ IELs were still significantly higher than in controls.

Percentages of FasL+ intraepithelial lymphocytes (IELs) in 10 untreated coeliac patients compared with those of 10 treated coeliac patients and 10 biopsied controls. Those coeliac patients on a gluten free diet and with a persistent degree of villous atrophy are indicated with a different symbol (solid square ). Horizontal bars represent median values.
After treatment, levels of FasL expressed by both LPLs and IELs were higher in those three patients with persistent villous atrophy.
JAM TEST
Figure 3 shows the killing activity of LPLs freshly isolated from untreated coeliac mucosa (fig 3A), treated coeliac mucosa (3B), and normal mucosa (3C). In untreated CD (3A), this activity was related to the effector target ratio and remained evident at the lowest ratio of 0.625:1. At each ratio this killing activity was significantly higher (p<0.0005) than those observed when LPLs from treated (3B) or normal mucosa (3C) were used. In treated CD, LPL cytotoxicity was significantly higher than in normal mucosa only at the highest effector target ratio.

FasL mediated cytotoxicity by lamina propria lymphocytes (LPLs) (effectors) obtained from 10 untreated coeliac patients (A), 10 treated coeliac patients (B), and 10 biopsied controls (C) against Jurkat cells (targets). 3H labelled Jurkat cells were cultured with LPLs at various effector/target ratios (5:1; 2.5:1; 1.25:1; and 0.625:1). The percentage of DNA fragmentation of Jurkat cells was assayed after 12 hours. FasL mediated cytotoxicity by intraepithelial lymphocytes (IELs) (effectors) obtained from 10 untreated coeliac patients (D), 10 treated coeliac patients (E), and 10 biopsied controls (F) against Jurkat cells (targets). 3H labelled Jurkat cells were cultured with IELs at various effector/target ratios (3:1; 1.5:1; and 0.625:1). The percentage of DNA fragmentation of Jurkat cells was assayed after 12 hours.
Regarding the cytotoxicity of IELs, at each ratio their killing activity was significantly higher (p<0.001) using IELs isolated from untreated coeliac mucosa (3D) than that using IELs from treated (3E) or normal mucosa (3F). Similar to LPLs, a gluten free diet, which also led to a clear improvement in duodenal histology, did not normalise cytotoxicity of IELs.
CYTOTOXICITY ASSAY
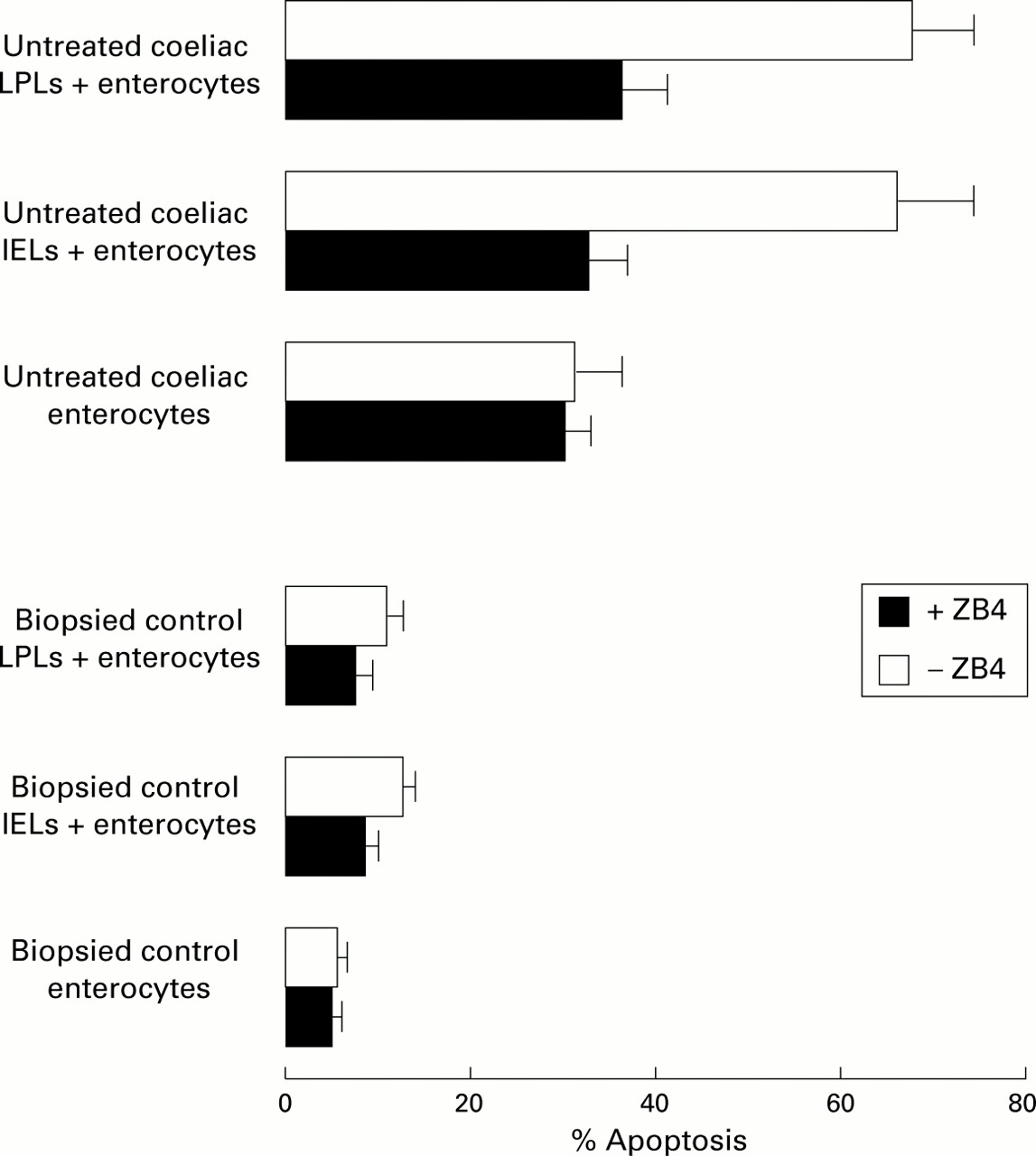
To confirm that the Fas/FasL cognate interaction was the mechanism by which LPLs and IELs mediated the killing of enterocytes in CD, the antagonist ZB4 anti-Fas antibody which blocks the Fas/FasL interaction was added to the coculture of coeliac lymphocytes and enterocytes. After four hours of coculture, most target untreated coeliac enterocytes when incubated with untreated coeliac LPLs or IELs were TUNEL+ (68.4 (8.1)% and 67.4 (9.3%), respectively). In contrast, when ZB4 anti-Fas blocking was added to the coculture, both percentages significantly decreased (36.4 (5.1)% and 32.8 (4.8)%, respectively), as shown in fig 4. These data suggest that Fas/FasL binding may play a central role in determining enterocyte apoptosis in CD. On the other hand, when both control LPLs and IELs were cocultured with normal enterocytes, the proportions of TUNEL+ target cells were poor (11.9 (2.5)% and 12.7 (3.5)%, respectively) and further decreased (7.4 (2.0)% and 8.8 (2.4)%, respectively) when ZB4 anti-Fas blocking was added to the coculture. The percentages of untreated coeliac TUNEL+ enterocytes, which were cultured without effector cells, did not significantly change in the absence (31.1 (5.2)%) or presence (28.9 (4.0)%) of ZB4 anti-Fas blocking antibody. The percentages of control TUNEL+ enterocytes, when cultured without effector cells, also did not significantly change in the absence (6.1 (1.3)%) or presence (5.9 (1.4)%) of ZB4 anti-Fas blocking antibody.

Antagonist ZB4 anti-Fas antibody reduces enterocyte apoptosis induced by both lamina propria lymphocyte (LPL) and intraepithelial lymphocyte (IEL) cytotoxic activity through a Fas/FasL interaction. Untreated coeliac and control enterocytes alone, and coculture of both untreated coeliac and control LPLs and IELs with enterocytes (at an effector target ratio of 20:1 and 100:1, respectively) were incubated for four hours in the presence or absence of 2 μg/ml ZB4 antagonist antibody. The percentage of apoptotic cells was determined by TUNEL detection.
LPL AND IEL APOPTOSIS
Cell Death Detection ELISA for oligonucleosomes on cell lysates was used to evaluate apoptosis of freshly isolated LPLs and IELs. Figure 5A shows increased LPL apoptosis in untreated CD (median 13.4, range 6.9–20.8) compared with treated coeliac patients (median 2.5, range 0.9–4.2) and biopsied controls (median 0.6, range 0–1.8). In treated CD, LPL apoptosis was still significantly higher than in biopsied controls. Moreover, fig 6 shows that LPLs showed higher expression of Fas receptor in untreated CD (median 29.8%, range 15.3–52.4 in 10 patients) compared with treated CD (median 4.9%, range 0.2–14.0 in 10 patients) and controls (median 2.7%, range 0.1–1.7 in 10 consenting subjects).

Detection of apoptosis on lamina propria lymphocytes (LPLs) (A) and intraepithelial lymphocytes (IELs) (B) by in vitro determination of cytoplasmic histone associated DNA fragments (mono and oligonucleosomes) in 10 untreated coeliac patients, 10 treated coeliac patients, and 10 biopsied controls. Quantitative results are expressed as an enrichment factor of mono and oligonucleosomes compared with controls. Horizontal bars represent median values.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Representative flow cytometric histograms of Fas expression on lamina propria lymphocytes (LPLs) isolated from duodenal biopsy specimens of an untreated coeliac patient, a treated coeliac patient, and a biopsied control.
Figure 5B shows that the pattern of IEL apoptosis was opposite to that of LPLs. In untreated coeliac mucosa (median 0.2, range 0–2.1), IEL apoptosis was lower than in treated coeliac patients (median 14.5, range 6.7–21.4) and biopsied controls (median 25.1, range 18.4–30.6). After treatment, IEL apoptosis was still significantly lower than in controls.
Discussion
In the present study, the cytotoxic activity of intestinal lymphocytes was quantified in CD. LPLs and IELs isolated from untreated coeliac mucosa shared higher levels of FasL and an increased and dose dependent killing activity against Fas expressing Jurkat cells compared with those isolated from treated coeliac and normal mucosa. In treated CD, both LPL and IEL cytotoxicity was still significantly higher than that of control lymphocytes. The lack of complete normalisation of cytotoxic activity by LPLs after a gluten free diet may be related to the capacity of dendritic cells to retain gliadin in its native form for a long time15 and/or to the inadvertent ingestion of this protein. In fact, as regards the persistent activation of IELs, it has been shown that minimal amounts of gliadin may expand IEL populations and activate local immunity, even when the mucosal structure looks otherwise normal.16 ,17 On the other hand, our finding of higher FasL expression by both LPLs and IELs in those treated coeliac patients with persistence of mucosal lesions is further evidence of the involvement of FasL mediated lymphocyte cytotoxicity against epithelial cells in this condition.
To demonstrate that both LPLs and IELs function effectively as killers of enterocytes via a Fas/FasL interaction in active CD, the effect of the antagonist ZB4 anti-Fas antibody on apoptotic activity exerted by coeliac lymphocytes against enterocytes was analysed. The significant reduction in enterocyte apoptosis observed after coculturing coeliac lymphocytes and enterocytes in the presence of ZB4 antibody is additional proof of the involvement of the Fas/FasL system in CD associated enterocyte apoptosis. This evidence couples with our previous demonstrations that Fas is upregulated on coeliac enterocytes2 ,8 and because of the proximity of IELs and enterocytes, it is conceivable that the Fas/FasL mechanism may well be implicated in in vivo enterocyte apoptosis in this condition. With regard to LPLs, their killing activity may be exerted by membrane bound FasL through pores within the basement membrane, which allow their intimate contact with basolateral projections of enterocytes.18 In addition, other mechanisms may be operating, such as the role of a soluble form of FasL released by cleavage by matrix metalloproteases,19 which have been shown to be overexpressed in untreated coeliac mucosa.20
As it has been shown that coeliac mucosa is a site of persistent T cell activation and proliferation,4 ,21 it was therefore of interest to determine whether such cells themselves showed an abnormal pattern of apoptosis. LPLs shared an increased apoptosis and higher levels of Fas receptor in untreated CD compared with controls. It is known that Fas/FasL expression is closely related to the process of lymphocyte activation, and that their interaction is crucial for its control and termination.22 Therefore, it is likely that in untreated CD the increased propensity of LPLs to undergo apoptosis is a homeostatic mechanism consequent to their greater activation.23
A novel result of the present study is decreased apoptosis of IELs in untreated CD. Intestinal mucosa is chronically exposed to an abundance of dietary antigens and exogenous pathogens, and this continuous antigen challenge may abnormally activate IELs.24 ,25 This activation does not normally result in inflammatory tissue damage, implying the involvement of an endogenous regulating mechanism. Our results in normal subjects confirm that a certain degree of lymphocyte apoptosis contributes to maintain IEL homeostasis by limiting expansion of activated T cells, and by ensuring the clearance of primed lymphocytes that are no longer required.26 Our finding of decreased IEL apoptosis is of particular relevance in the context of untreated CD. This defect, which is incompletely healed by a gluten free diet, may determine the non-deletion of autoreactive T cell clones, present in intestinally matured T cells,27 and/or expansion of IELs with an abnormal phenotype and a restricted repertoire, and this may precede the onset of a malignant condition. These findings are particular intriguing when considering that refractory sprue, which is an established complication of CD, is characterised by an abnormal IEL population,28 and that intestinal T cell lymphoma arises from IELs.29
The possible mechanisms responsible for decreased IEL apoptosis in untreated CD are at present under investigation in our laboratory. Upregulation of Bcl-2 molecules30 and/or lack of downregulatory signals by coeliac enterocytes for IELs31 are likely candidates. In addition, IEL resistance to apoptosis may be a consequence of enterocyte secretion of interleukin 15, a cytokine which has been shown to protect cells from induction of apoptosis32 and to be abundant in the epithelial compartment of active CD where it promotes the selective expansion of CD94+ IELs.33 Finally, in murine small intestine, IELs from villous tips and crypt portions show different propensities to undergo apoptosis.34 ,35 On this basis, it would be interesting to analyse whether the abnormal spatial segregation of IELs in CD may contribute to their decreased apoptosis.
In conclusion, the present study demonstrates that coeliac LPLs and IELs show reversed patterns of apoptosis but a similar increase in FasL mediated cytotoxic activity. It is noteworthy that this activity, which has been fully documented for IELs,8 ,36 has also been reported for LPLs in a variety of disease conditions.37-39
Abbreviations used in this paper
- CD
- coeliac disease
- FasL
- Fas ligand
- IELs
- intraepithelial lymphocytes
- LPLs
- lamina propria lymphocytes
- PBS
- phosphate buffered saline
- FCS
- fetal calf serum
- TUNEL
- terminal deoxynucleotidyl transferase mediated digoxigenin-deoxyuridine triphosphate nick end labelling