Article Text

Abstract
BACKGROUND Deoxycholic acid (DCA), implicated in the pathogenesis of gall stones and colorectal cancer, is mainly formed by bacterial deconjugation (cholylglycine hydrolase (CGH)) and 7α-dehydroxylation (7α-dehydroxylase (7α-DH)) of conjugated cholic acid (CA) in the caecum/proximal colon. Despite this, most previous studies of CGH and 7α-DH have been in faeces rather than in caecal contents. In bacteria, CA increases 7α-DH activity by substrate-enzyme induction but little is known about CA concentrations or CA/7α-DH induction in the human colon.
AIMS AND METHODS Therefore, in fresh “faeces”, and in caecal aspirates obtained during colonoscopy from 20 patients, we: (i) compared the activities of CGH and 7α-DH, (ii) measured 7α-DH in patients with “low” and “high” percentages of DCA in fasting serum (less than and greater than the median), (iii) studied CA concentrations in the right and left halves of the colon, and examined the relationships between (iv) 7α-DH activity and CA concentration in caecal samples (evidence of substrate-enzyme induction), and (v) 7α-DH and per cent DCA in serum.
RESULTS Although mean CGH activity in the proximal colon (18.3 (SEM 4.40) ×10−2 U/mg protein) was comparable with that in “faeces” (16.0 (4.10) ×10− 2 U/mg protein) , mean 7α-DH in the caecum (8.54 (1.08) ×10-4 U/mg protein) was higher (p<0.05) than that in the left colon (5.72 (0.85) ×10-4 U/mg protein). At both sites, 7α-DH was significantly greater in the “high” than in the “low” serum DCA subgroups. CA concentrations in the right colon (0.94 (0.08) μmol/ml) were higher than those in the left (0.09 (0.03) μmol/ml; p<0.001) while in the caecum (but not in the faeces) there was a weak (r=0.58) but significant (p<0.005) linear relationship between 7α-DH and CA concentration. At both sites, 7α-DH was linearly related (p<0.005) to per cent DCA in serum.
INTERPRETATION/SUMMARY These results: (i) confirm that there are marked regional differences in bile acid metabolism between the right and left halves of the colon, (ii) suggest that caecal and faecal 7α-DH influence per cent DCA in serum (and, by inference, in bile), and (iii) show that the substrate CA induces the enzyme 7α-DH in the caecum.
- deoxycholic acid
- 7α-dehydroxylation
- gall stones
- colorectal cancer
Abbreviations used in this paper
- CA
- cholic acid
- DCA
- deoxycholic acid
- 7α-DH
- 7α-dehydroxylase
- CGH
- cholylglycine hydrolase
- TLC
- thin layer chromatography
Statistics from Altmetric.com
Colonic bacteria play a major role in bile acid metabolism. Thus anaerobic bacterial enzymes convert conjugated primary bile acids, such as the glycine and taurine conjugates/amidates of cholic acid (CA), into unconjugated secondary bile acids, such as deoxycholic acid (DCA), in two major steps:
- (i)
- deconjugation, whereby the enzyme cholylglycine hydrolase (CGH) deconjugates (or deamidates) the conjugated bile acids to yield the corresponding unconjugated species, followed by the
- (ii)
- 7α-dehydroxylation pathway.
It is believed that these enzymatic reactions take place mainly in the caecum and ascending colon.1 ,2 Logically therefore the activities of these enzymes should be measured in the proximal colon or in samples obtained from the caecal lumen, rather than in stools. Before the present studies however, human CGH and 7α-dehydroxylase (7α-DH) activities were measured in isolated bacterial strains (single or mixed),3-7 or in faeces,8 rather than in colonic contents, no doubt because in humans the caecum and ascending colon are relatively inaccessible. In at least two respects however the luminal milieu in the proximal large bowel is different from that in the distal colon (or in fresh faeces). Firstly, the ratio of anaerobic: aerobic bacteria is greater in the faeces than in the caecum9 which is of relevance as 7α-dehydroxylation is carried out only by anaerobic bacteria. Secondly, the luminal pH is lower in the proximal than in the distal colon.10 ,11 In theory, such changes in luminal pH could affect the growth of bacteria in the colon.12 They could also influence the reaction rates of the two bile acid metabolising enzymes, both of which are pH dependent.1 ,2
The 7α-dehydroxylation pathway is inducible by its principal substrate CA.13 However, by the time bile acids reach the faeces almost all have been converted from primary to secondary bile acids.3 ,14 In theory therefore the absence of CA from the contents of the left colon/faeces means that there is no longer any substrate available for induction of 7α-DH. If such induction is important quantitatively, one might expect corresponding inequalities in CA 7α-DH activity between the right and left halves of the colon. Despite this, there is no information about differences in bile acid metabolising enzyme activities between caecal contents and faeces.
The results of studies by Berr and colleagues8 showed that faecal 7α-DH activity is significantly greater in patients with high versus those with low DCA pool sizes and/or DCA: CA molar ratios. To see if similar differences are present in caecal and faecal contents, we calculated the median per cent DCA in fasting serum (19.7%) and used this to divide patients retrospectively into “low” and “high” DCA subgroups, and compared caecal and faecal 7α-DH activities in the two populations.
The aims of this hypothesis driven study therefore were to determine whether:
- (i)
- the activities of the bile acid metabolising enzymes (CGH and 7α-DH), and the concentrations of unconjugated CA, are different in caecal and faecal samples,
- (ii)
- in patients with high versus those with low circulating DCA levels (as indicated by the per cent DCA in fasting serum8), the activities of the two enzyme pathways in question are different,
- (iii)
- there is a relationship between 7α-DH activity (in both caecal and faecal samples) and serum DCA,
- (iv)
- as a result of bacterial biotransformation (and/or absorption), the concentrations of endogenous CA change in the luminal contents, from the right to the left halves of the colon, and
- (v)
- there is evidence of induction of the 7α-DH enzyme pathway by its substrate (CA) in caecal aspirates.
Patients and methods
PATIENTS
Twenty patients were studied (11 stone free “controls” and nine with cholesterol rich gall bladder stones). Age range was 57–69 years and 15 were women. All 20 had undergone clinically indicated colonoscopy (during which caecal contents were sampled by aspiration, see below) and had been found, in retrospect, to have normal colons.
The decision to carry out the present study was taken halfway through a larger companion study15 which involved a total of 40 patients. However, once the decision had been reached, the next 20 patients were recruited sequentially with no further selection.
SAMPLING PROXIMAL COLONIC CONTENTS
As a prelude to colonoscopy, the contents of the left colon were washed out using a phosphate enema, with the aim of leaving those in the right colon undisturbed. The use of this instant enema enabled us to examine the large bowel mucosa of the left colon endoscopically, after which the scope was advanced into the unprepared right colon, proximal to the hepatic flexure.
To facilitate collection of the caecal/proximal colonic contents, 20 ml of sterile saline were injected into the colonic lumen through the biopsy channel of the endoscope. The caecal contents were then homogenised in situ using rotatory movements of the tip of the colonoscope, and repeated aspiration/reinjection cycles of the luminal contents through the biopsy channel. The resultant mixture was aspirated into a trap pregassed with nitrogen. In order to transfer samples from the endoscopy department to the microbiology laboratory under anaerobic conditions, we used the GasPak anaerobic pouch system. The homogenised caecal aspirates reached the laboratory within one hour of collection.
SAMPLING FRESH “FAECES”
As noted above, before patients were endoscoped they were given a phosphate enema which stimulated the passage of “faeces” (left colonic contents). A 1.0 g aliquot of this enema washout material was immediately homogenised with 25% (w/v) saline, and rapidly (again within one hour) transferred to an anaerobic environment, using the GasPak anaerobic pouch system. In this way, the “caecal” (proximal colonic aspirate) and the “faecal” (aliquot from the left colonic washout) samples were collected from the same patient, at the same time.
BILE ACID METABOLISING ENZYME ACTIVITIES IN CAECAL ASPIRATES AND FAECES
The activities of both bile acid metabolising enzymes (see below) were measured in proteins precipitated from sonicates of the caecal and faecal samples, using saturated ammonium sulphate, as previously described.16 For both enzymes, the results are expressed as units (U) of enzyme activity where I U=1 μmol of product formed per mg of bacterial protein per minute.
Cholylglycine hydrolase
The substrate for the deconjugation reaction was 10 mM taurocholic acid (0.5 ml substrate incubated with 0.5 ml of the protein extract), as previously described.17 The product of this reaction (CA) was extracted using a C18 column,18 isolated by thin layer chromatography (TLC)19 and measured with the 3α-hydroxysteroid dehydrogenase assay20 at 340 nm.
Cholic acid 7α-dehydroxylation
The reaction conditions for this enzyme were the same as those described elsewhere.17 In brief, 0.5 ml of the protein extract was incubated with non-isotopic CA (2 mM, used to catalyse the reaction by substrate-enzyme induction13), plus14C-CA (200 000 dpm). Bile acids in the reaction mixture were then extracted twice using ethyl acetate, separated, and identified using TLC, and the [14C] radioactivity in the DCA fraction measured in a liquid scintillation counter.
Cholic acid concentrations in caecal and faecal samples
To measure levels of endogenous CA in caecal and faecal samples, a blank was run as follows: a 0.5 ml aliquot of the sample was incubated with 0.5 ml of sterile saline using the conditions described above for the deconjugation reaction (except that no substrate was added).
As before, after the incubation was complete, 1 ml of 0.1 M NaOH was added to the reaction mixture and the sample centrifuged to separate the supernatant and precipitate phases. The supernatant fluid was then loaded onto a C18 column to extract bile acids present in the original sample.
Bile acids were separated using TLC, and the area corresponding to the CA standard was scraped from the TLC plate, reacted with 3α-hydroxysteroid dehydrogenase, and measured spectrophotometrically as previously described.20
Serum DCA
Concentrations of DCA (total, conjugated, and unconjugated fractions) were measured in fasting serum using gas chromatography-mass spectrometry21 ,22 and the proportions calculated, as previously described.23 In this paper however we refer only to the unconjugated fraction of DCA in fasting serum.
As noted above, the median per cent DCA in fasting serum was 19.7%, and despite the fact that concentrations of DCA in fasting serum were normal in all 20 individuals, this median was used to divide patients, retrospectively, into “low” and “high” serum DCA subgroups, as follows:
- (i)
- “low” DCA subgroup (<19.7% DCA in fasting serum: n=10); mean (SEM) serum DCA concentration 0.28 (0.06) μM.
- (ii)
- “high” DCA subgroup (>19.7% DCA in fasting serum: n=10); mean (SEM) serum DCA concentration 0.54 (0.10) μM.
We then used univariate analyses to examine the inter-relationships between per cent DCA in fasting serum and the activities of the two bile acid metabolising enzymes in both caecal and faecal samples.
Evidence of substrate-enzyme induction in caecal aspirates
To test the hypothesis that the substrate (CA) might induce the enzyme system (7α-DH) in the proximal colon, we used univariate analysis to examine the relationship between these variables in caecal aspirates.
Relationship between 7α-DH activity and per cent DCA in fasting serum
The role of caecal 7α-DH activity in DCA metabolism was addressed in detail in our companion paper.15 To avoid duplication therefore, the relationship between the dehydroxylating enzyme system and serum DCA is confined in this study to the correlation coefficients between 7α-DH (in both caecal and faecal samples) and per cent DCA in serum.
ETHICAL CONSIDERATIONS
The study was approved by the research ethics committee of Guy's and St Thomas' Hospitals. All patients gave their written informed consent.
STATISTICAL ANALYSES
Unless otherwise stated, all results are expressed as mean (SEM).
The significance of differences in results between the two groups was tested with the heteroscedastic t test using Excel Software version 5.0 (Microsoft Corporation, 1 Microsoft Way, Redmond, WA 98052-6399, USA); p values <0.05 were considered to be statistically significant.
Univariate analyses were performed using SPSS software; the statistical significance of the correlation coefficients was examined using an unpaired two tailed Student's t test.
Results
CHOLYLGLYCINE HYDROLASE
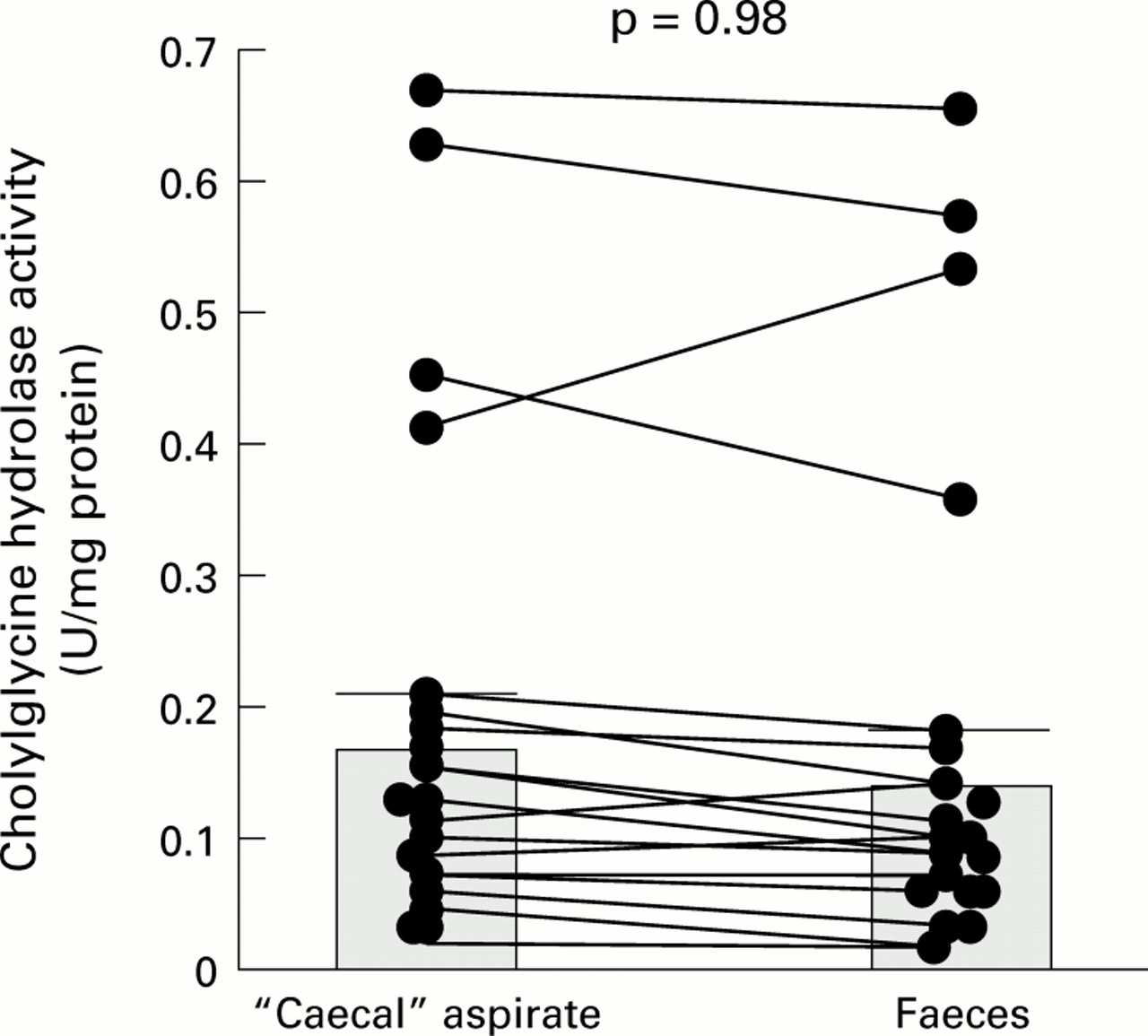
The paired results, comparing CGH activity in caecal and faecal samples from the same individuals, are shown graphically in fig1.
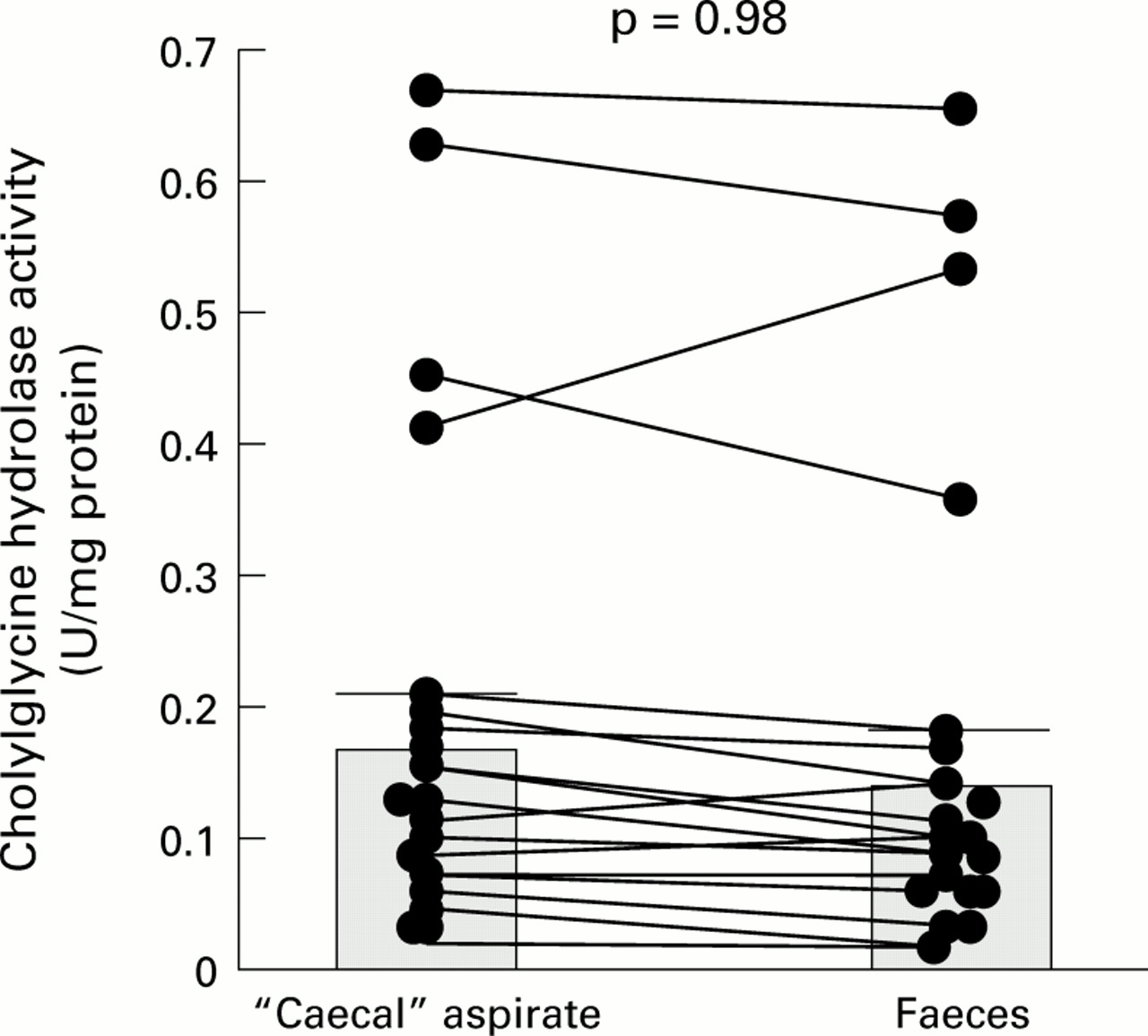
Paired results for cholylglycine hydrolase activity in “caecal” and faecal samples: individual data points with means (SEM).
As the results in fig 1 show, most of the CGH results were tightly bunched (<0.2 enzyme units/mg protein). However, in a subgroup of four patients (one stone free, three stone carriers), CGH activities were appreciably higher (range 0.35–0.66 U/mg protein) than those in the remaining 16 patients. None the less, when the activities in all 20 patients were pooled, there were no significant differences between mean enzyme activities for the caecal (0.18 (SEM 0.04) enzyme units/mg protein) and faecal (0.16 (0.04) enzyme units/mg protein) samples. Furthermore, when the CGH results in the low and high DCA groups were compared (table 1), again there was no difference in mean values between the two subgroups.
Cholylglycine hydrolase (CGH) activities in caecal and “faecal” samples from the low and high serum deoxycholic acid (DCA) subgroups (see text)
CHOLIC ACID 7α-DEHYDROXYLATION
Corresponding paired results for 7α-DH activity in caecal and faecal samples (n = 20) are shown in fig 2.
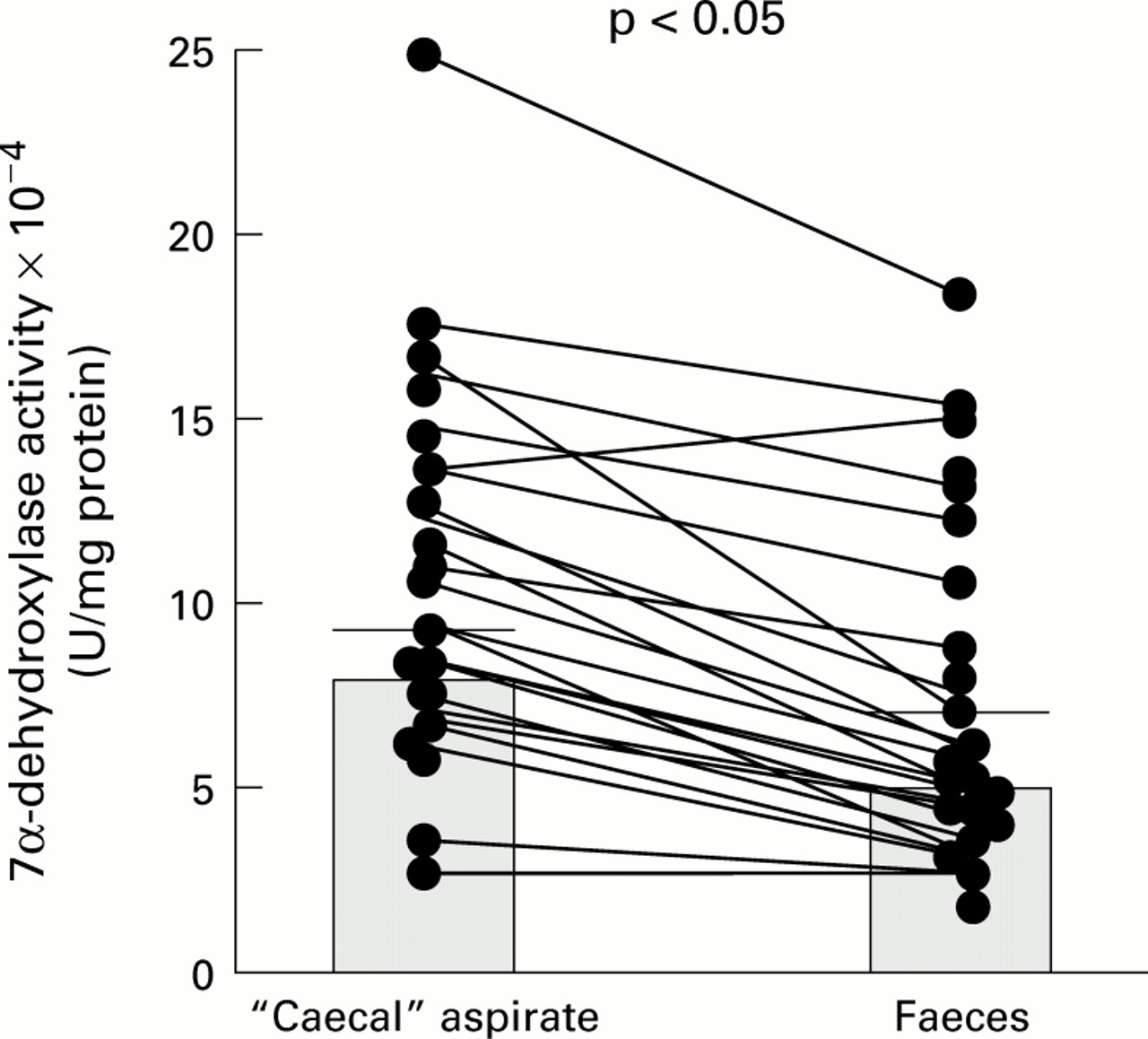
Paired results for 7α-dehydroxylase activity in the “caecal” and faecal samples: individual data points with means (SEM).
With three exceptions, 7α-DH activity was always higher in caecal aspirates than in matched faecal samples such that for all 20 patients the mean value in the caecum (8.54 (1.08) ×10−4 U/mg protein) was 49% greater than that in faeces (5.72 (0.85) ×10−4 U/mg protein; p<0.05).
CAECAL AND FAECAL 7α-DH ACTIVITIES IN THE LOW VERSUS HIGH SERUM DCA SUBGROUPS
These results are shown graphically, in different ways, in figs 3and 4.

Paired results linking “caecal” and faecal 7α-dehydroxylase activities in the low (A) versus high (B) serum deoxycholic acid (DCA) subgroups: individual data points with means (SEM).
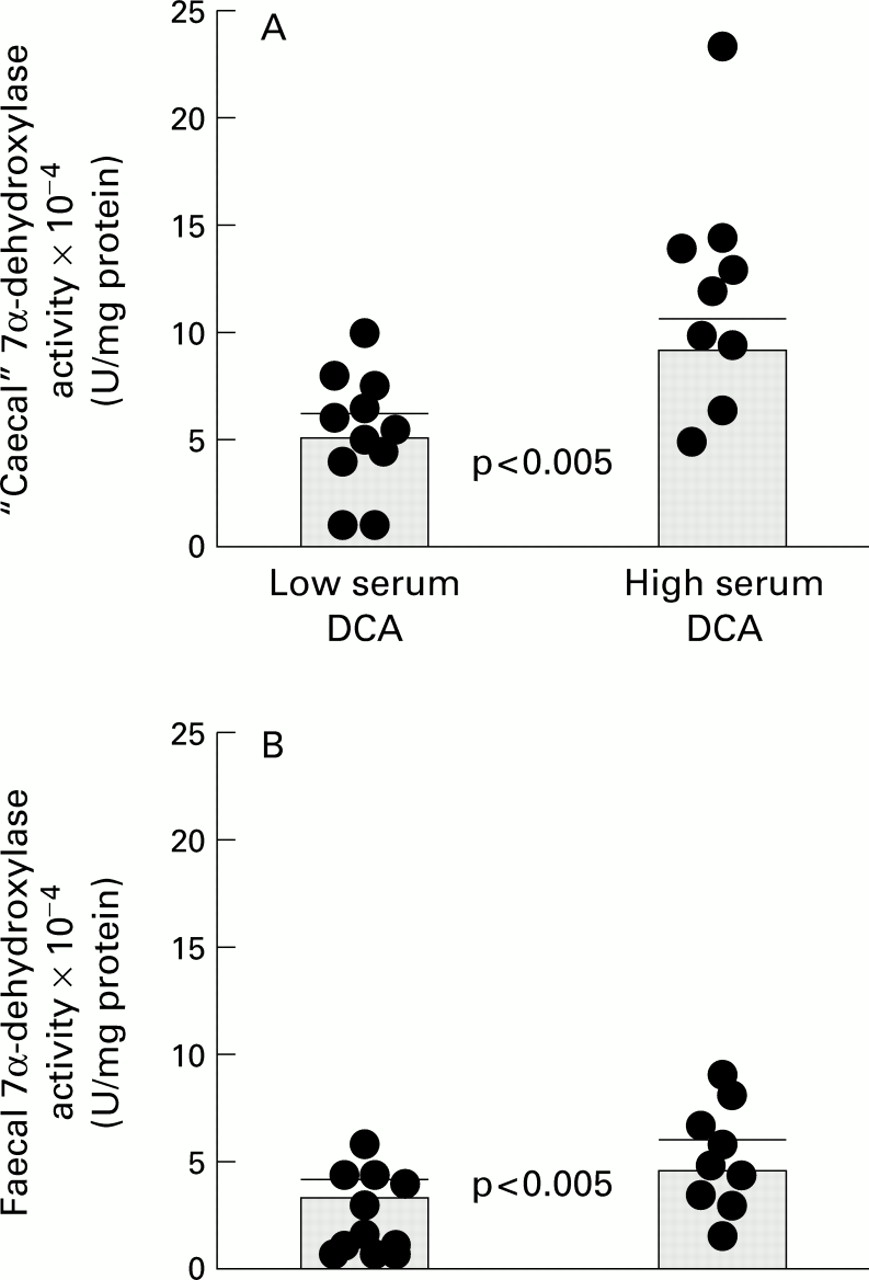
Non-paired results comparing “caecal” (A) and faecal (B) 7α-dehydroxylase activities in the low versus high serum deoxycholic acid (DCA) subgroups: individual data points with means (SEM).
(i) Paired results, linking caecal and faecal 7α-DH activities in the two subgroups (fig 3)
The same pattern of results, as that noted above when 7α-DH activity was compared in paired caecal and faecal samples from all 20 individuals, was seen when the corresponding linked comparisons were made in the low and high serum DCA subgroups. Thus in the 10 individuals with “low” proportions of DCA in fasting serum (less than the median of 19.7%), the mean 7α-DH activity in “faeces” (3.75 (1.25) ×10−4 U/mg protein) was 29% less than that found in the caecum (5.28 (0.76) ×10−4 U/mg protein) in this study (p<0.01). Similarly, in the 10 patients with “high” percentages of DCA in fasting serum (greater than the median of 19.7%), the mean 7α-DH activity in faecal samples (4.47 (1.39) ×10−4 U/mg protein) was just under half (45%) that found in the washout from the proximal colon (9.84 (0.9) ×10−4 U/mg protein). However, this difference was not statistically significant.
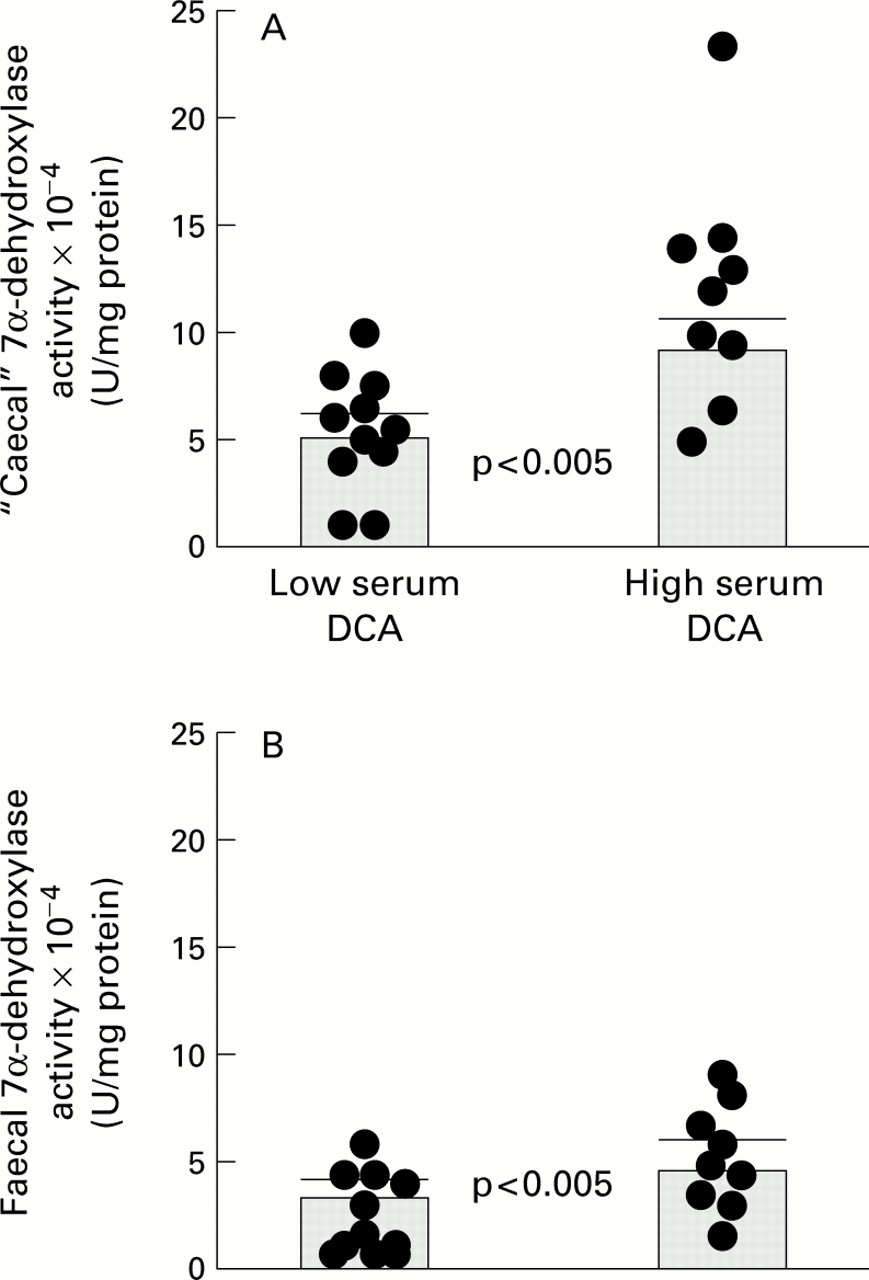
(ii) Non-paired results, comparing caecal and faecal 7α-DH activities, in the two subgroups
Figure 4 compares 7α-DH results in the low versus high serum DCA subgroups for caecal and faecal samples.
Mean caecal 7α-DH activity in the high DCA subgroup (11.95 (1.68) ×10−4 U/mg protein) was 118% greater than that in the low DCA subgroup (5.48 (0.98) ×10−4 U/mg protein; p<0.005). There was a similar pattern of results for faecal 7α-DH, the mean activity in the high DCA subgroup (9.27 (1.17) ×10−4 U/mg protein) being 160% greater than that in patients with low serum DCA (3.56 (0.59) ×10−4 U/mg protein; p<0.005).
ENDOGENOUS CHOLIC ACID CONCENTRATION
The paired data for CA concentrations in the caecal and faecal samples from all 20 individuals are shown in fig 5.
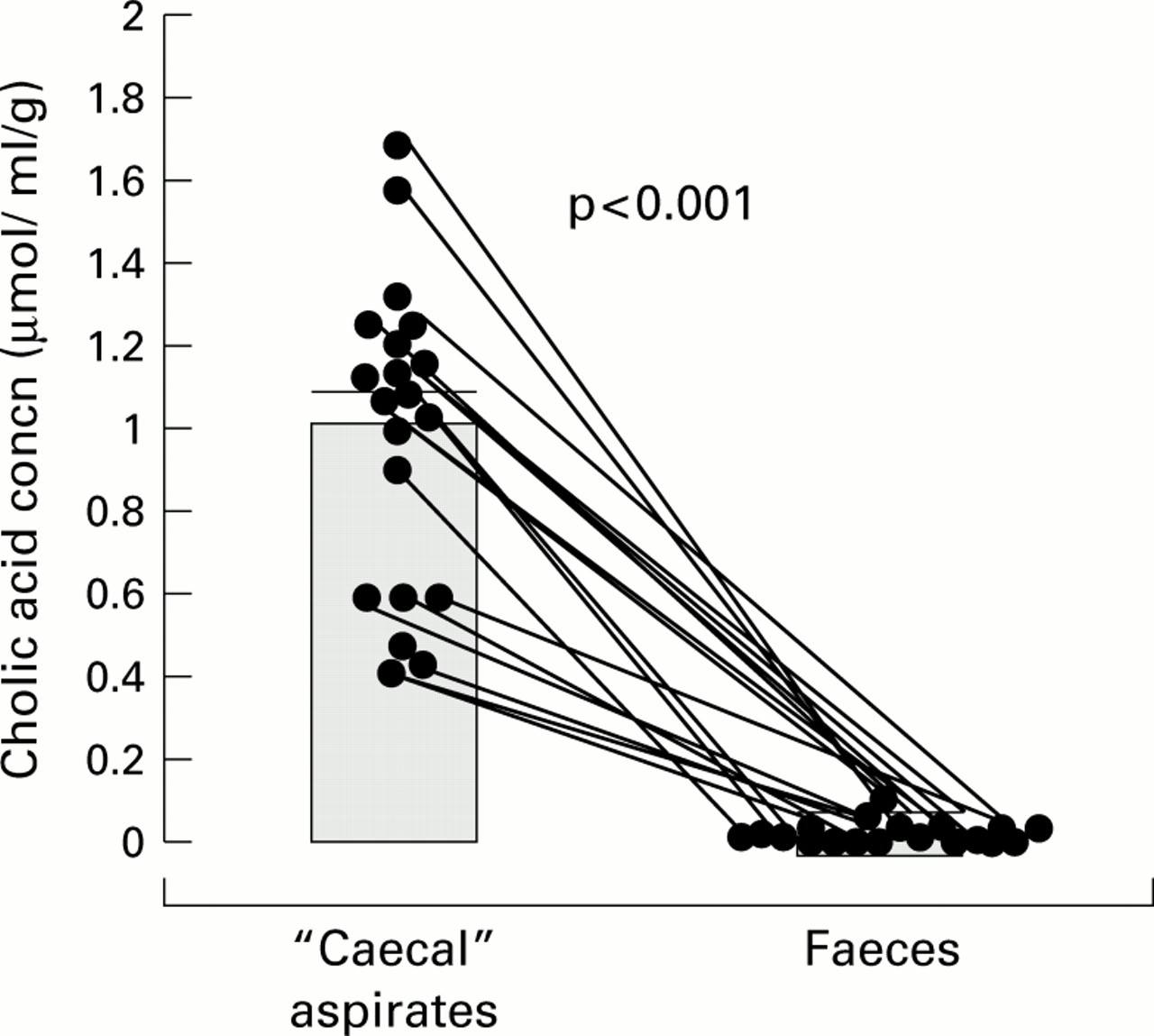
Endogenous cholic acid concentrations in paired “caecal” and faecal samples: individual data points with means (SEM).
As the results in fig 5 show, the concentration of CA in the caecal aspirates varied considerably from 0.14 to 1.65 with a mean of 1.08 (0.14) μmol/ml. As predicted however, in every case there was a marked and consistent (p<0.001) fall in CA concentration between the right and left halves of the colon such that CA was absent, or virtually so, in most of the “faecal” samples. Moreover, when we compared CA concentrations in the caecal aspirates from the “low” and “high” serum DCA subgroups, mean CA concentrations were again significantly greater in the caecal aspirates than in the “faeces” (table 2).
Cholic acid (CA) concentrations in caecal and “faecal” samples from the low and high serum deoxycholic acid (DCA) subgroups (see text)
RELATIONSHIP BETWEEN 7α-DH ACTIVITY AND CA CONCENTRATIONS IN THE CAECAL SAMPLES
When the relationship between the activity of 7α-DH (U/mg protein) and endogenous CA concentration was studied in the caecal aspirates (fig 6), there was a weak (r=0.58) but significant (p<0.005) linear correlation between the two variables. However, in the faecal samples there was no such significant relationship.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Relationship between 7α-dehydroxylase activity and cholic acid concentration in the “caecal” samples.
RELATIONSHIP BETWEEN 7α-DH ACTIVITY AND SERUM DCA
There were significant linear relationships between the per cent DCA in fasting serum and the activities of 7α-DH in the caecal (r=0.89) and faecal (0.65) samples: p<0.005 at both sites.
Discussion
STUDY DESIGN
Although the broad aim of these studies was to compare various aspects of CA and DCA metabolism in caecal and faecal samples from the same individuals, as noted in the methods section the techniques used to sample colonic luminal contents have their limitations. Thus “caecal” samples were obtained endoscopically from a site somewhere proximal to the hepatic flexure. However, in the absence of fixed anatomical landmarks, it is difficult to be precise about the exact site of aspiration during clinical colonoscopy. Furthermore, although we refer throughout the paper to “faeces” it could be argued that the bile acid composition and the bacterial enzyme activities in material washed out from the left colon by a phosphate enema are not necessarily the same as those in freshly passed stools. Certainly the results of clinical and radiological studies show that formed faeces often linger in the descending and sigmoid colon for 24 hours or more, before they are evacuated.24 During that time, there could well be further changes in 7α-DH activity (although probably not in CA concentrations as these were already almost zero in the enema washout material). Hence the results of the present study probably underestimate the magnitude of the differences in DCA metabolism between the right and left halves of the colon.
In this study, the CGH and 7α-DH results were expressed as units of enzyme activity per mg protein in the colonic luminal contents. While we cannot assume that the protein of the caecal and faecal samples comes exclusively from bacteria, the fact that the CGH and 7α-DH activities are expressed in U/mg protein means that they approximate to enzyme specific activities—that is, the amounts of enzyme per bacterium.
DISTRIBUTION OF RESULTS FOR CGH AND 7α-DH ACTIVITIES AT BOTH SAMPLING SITES
There is no obvious explanation for the non-Gaussian distribution of the CGH results at both colonic sampling sites. However, in the subgroup of four individuals with high caecal CGH activities (three of whom had cholesterol gall stones), the corresponding faecal samples also had relatively high levels of CGH activity. Moreover, in these four patients mean 7α-DH activity both in the caecal aspirates (11.53 (4.01) ×10−4 U/mg protein) and in faeces (7.93 (2.87) ×10−4 U/mg protein) was also comparatively high, perhaps because high CGH activity yields high concentrations of unconjugated CA which in turn stimulate increased 7α-DH activity by substrate-enzyme induction.13 ,25 Whatever the reason for the non-Gaussian distribution of the results, the fact remains that there was no difference in CGH activities between caecal and faecal samples while there was an obvious and significant difference in 7α-DH at these two sites.
DIFFERENCES IN CGH AND 7α-DH ACTIVITIES BETWEEN CAECAL AND FAECAL SAMPLES
There are at least two possible explanations for the different pattern of results for CGH and 7α-DH activities between the right and left halves of the colon. Firstly, CGH is a ubiquitous enzyme present in many intestinal bacterial species—both aerobes and anaerobes.3 In contrast, CA 7α-DH is found in only a few strains of Gram positive anaerobes2 ,8 and the number of these micro-organisms is relatively low in faeces. In fact, as shown by the results in figs 1 and 2, the mean activity of CGH in the caecum was more than 200 times that of 7α-DH. Given the relative abundance of CGH, it is unlikely to be rate limiting in the intestinal bacterial conversion of conjugated CA to unconjugated DCA whereas 7α-dehydroxylation might well be.
Secondly, 7α-DH activity is known to be inducible by its substrate CA13 ,25 while CGH is not. Thus in the caecum and ascending colon, although CA concentrations were variable, they were relatively high (up to 1.65 mM; fig 5) and linearly related to 7α-DH activity (fig 6).
DISTRIBUTION OF RESULTS FOR ENDOGENOUS BILE ACID CONCENTRATIONS IN THE CAECUM
(i) Primary bile acids
To test the substrate-enzyme hypothesis (by studying the relationship between CA concentration and 7α-DH in the caecum), we were relying on spontaneous variations in caecal CA concentration, rather than on pharmacological manoeuvres designed to perturb the balance between CA and DCA by using broad spectrum antibiotics,8 ,29 drugs that increase or decrease the rates of transit through the intestine,30 ,31 or regimens that change intracolonic pH, such as oral lactulose.32 ,33 Instead, we relied on spontaneous variations in “endogenous” caecal CA concentrations which, in the event, ranged from 0.14 to 1.65 μmol/ml although the explanation for this scatter in results is unknown. Presumably it reflects either the “spillover” of conjugated CA (which has escaped absorption in the small intestine) into the colon or passage through the ileocaecal valve of unconjugated CA (which has been formed by deconjugation in the terminal ileum34), or both.
(ii) Secondary bile acids
Factors influencing the concentration of dehydroxylated bile acids (such as DCA and lithocholic acid) in the caecum are equally complex. Thus McJunkin et al found that dehydroxylated bile acids were present in the caecum, even in patients with diarrhoeal disorders.35 At first sight this might suggest that not only deconjugation but also dehydroxylation begins in the terminal ileum. Thus the presence of DCA in the proximal colon depends not only on deconjugation and dehydroxylation of the amidates of CA but also on deconjugation of recycled conjugated DCA and on absorption of the nascent secondary bile acid. Measurement of dehydroxylated bile acid concentrations in, and absorption from, the colon fell outside the aims of the present study.
SIMILARITIES AND DIFFERENCES BETWEEN CA CONCENTRATION AND 7α-DH ACTIVITY IN CAECAL AND FAECAL SAMPLES
Initially, there seems to be no a priori reason why apparently independent variables such as bile acid concentration and bacterial enzymatic activity should change in parallel from one region of the large bowel to the next. None the less, the fact that they did so raises three further points for discussion.
Firstly, the observation that CA concentrations are relatively high in caecum/proximal colon but are almost zero in the faeces/enema washout of the distal colon seems initially to confirm the results of many previous studies3 ,14 which showed that by the time of defecation only secondary (bacterially modified) bile acids remain in faeces. However, the disappearance of CA from the colonic luminal contents during large bowel transit is due not only to 7α-dehydroxylation but also to CA absorption from the colon, mainly by passive non-ionic diffusion.26 As noted above, measurement of colonic bile acid absorption was not one of the objectives of the present study. Therefore, in the absence of the facts, we can only speculate about the relative contributions of 7α-dehydroxylation versus absorption to the decline of CA concentration during transit through the colon.
Secondly, the weak (r=0.58) but significant (p<0.005) linear relationship between CA concentration and 7α-DH activity in the caecum is consistent with the concept of substrate enzyme induction. However, the fact that there is continuing 7α-DH activity in the left colon, at a time when CA has virtually disappeared from the luminal contents, suggests that at least some of the dehydroxylating enzyme activity must be independent of its substrate.
Thirdly, the fact that 7α-DH activity is found on both sides of the colon (albeit in significantly reduced amounts on the left), suggests that the 7α-dehydroxylation pathway may be a pancolonic phenomenon. It also suggests that some DCA may be formed in, and absorbed from, the left colon. Indeed, based on the results of a multivariate analysis in our companion study,15 we tentatively reached just this conclusion. However, it is worth emphasising that the presence of 7α-DH activity in the luminal contents of the left colon cannot prove that the enzyme is biologically active there. Indeed, the precedent of a mismatch between the anatomical distribution of an enzyme's activity along the intestine and its maximal physiological site of action comes from studies of lactase activity in humans.27 Thus the peak activity of this disaccharidase is in the mid small bowel while the principal physiological site of lactose hydrolysis is normally in the duodenum and upper jejunum.
RESULTS IN “HIGH” VERSUS “LOW” SERUM DCA SUBGROUPS
The validity of dividing the 20 individuals retrospectively into two subgroups (“high” and “low” serum DCA) based on results above and below the median for per cent DCA in fasting serum is also open to question. In fact, as indicated in the methods section, absolute values for serum DCA concentrations were all within the normal range.28 None the less, the decision to divide patients on this basis (in much the same way that Berr and colleagues8had done previously) unmasked interesting and significant differences in mean caecal and faecal 7α-DH activities between the two subgroups.
Interpretation of the observation that patients in the high serum DCA subgroup also had high 7α-DH activities is also open to question. However, the implication of the results in fig 4 is that high activities of 7α-DH in both the right and left halves of the colon are associated with high percentages of DCA in fasting serum (and, by inference, high proportions of this bile acid in bile36 ,37 and in the bile acid pool38).
RELATIONSHIP BETWEEN ANAEROBIC INTESTINAL BACTERIA, 7α-DH ACTIVITY, AND DCA METABOLISM
The aims of the present study did not include measurements of anaerobic bacteria in intestinal contents or comparisons of results between stone free controls and gall stone patients. Instead, these were the objectives of our companion paper15 in which we showed (among other things) that mean caecal 7α-DH activity in gall stone patients was significantly greater than in stone free “controls”. Based on this and other evidence, we speculated that the principal mechanism for increased DCA formation in gall stone patients was due not to an increase in 7α-DH specific activity but to an increase in the numbers of total and Gram positive anaerobes (and therefore to an attendant increase in the total amount or biomass of the 7α-dehydroxylating enzymes in the colon). To prove this, one would need to carry out faecal dilution assays, which were not performed in the present study. However, from the results in our companion study,15 we know that high CGH and 7α-DH “specific activities” (such as those found in the subgroup of patients described here) are not significantly related to anaerobic bacterial counts.
Conclusions
Our results confirm that there are obvious regional differences in bile acid metabolism between the right and left halves of the colon. Therefore, one logical conclusion is that future studies of bile acid metabolising enzymes should be based on caecal/right colonic contents rather than on faeces. From a scientific standpoint, this conclusion is incontrovertible. However, the caecum and proximal colon are relatively inaccessible, and their contents cannot be sampled non-invasively whether by colonoscopy, peroral intubation, or during surgery, Moreover, although the right-left differences in colonic luminal CA concentrations were marked, those for 7α-DH activity, although statistically significant, were modest. In addition, the activity of 7α-DH in faeces usually, but not always, predicted its activity in the right colon. From a pragmatic standpoint therefore, it seems reasonable to conclude that sampling fresh faeces is an acceptable compromise for clinical studies of DCA metabolism.
Acknowledgments
This work was supported in part by grants from the John Ellerman Foundation and Novartis Pharma, Basel, Switzerland, to whom the authors are grateful. This work was presented in part at the 1998 Annual Scientific meeting of the British Society of Gastroenterology, and published in abstract form (Gut1998;42(Suppl 1):A11).
Abbreviations used in this paper
- CA
- cholic acid
- DCA
- deoxycholic acid
- 7α-DH
- 7α-dehydroxylase
- CGH
- cholylglycine hydrolase
- TLC
- thin layer chromatography