Article Text

Abstract
Background and aim —Some of the recently observed functional features characteristic of immunocompetent cells residing in the human intestinal lamina propria could be mediated by interleukin- 10 (IL-10). To investigate the role of IL-10 in the human intestinal mucosa, the regulation of IL-10 production by lamina propria T lymphocytes (LPL-T) was determined and compared with that of peripheral blood T lymphocytes (PBL-T).
Methods —Following activation by using different stimuli, IL-10 release by LPL-T and PBL-T into the supernatant was measured by enzyme linked immunosorbent assay (ELISA). In parallel, cell growth was determined by [3H]-thymidine incorporation.
Results —Neither LPL-T nor PBL-T release IL-10 constitutively. Triggering through CD2 or the T cell receptor (TCR)/CD3 complex in the presence of autologous monocytes induces significantly greater IL-10 secretion by LPL-T than by PBL-T. Engagement of the CD45 receptor enhances IL-10 release and proliferation of CD2 triggered CD45RO+ PBL-T. In contrast, it reduces CD2 induced IL-10 production by LPL-T without altering cell growth significantly .
Conclusions—Activated LPL-T release relatively high amounts of IL-10. Enhanced IL-10 production by activated LPL-T, in comparison with activated PBL-T, is not only related to the presence of a higher proportion of CD45RO+ T cells in the intestinal lamina propria, but is also caused by increased sensitivity of LPL-T to CD2 co-stimulation. The differential responsiveness of LPL-T, compared with PBL-T, to CD45 engagement demonstrates that CD45 could be involved in the altered CD2 reactivity of LPL-T.
- CD2
- CD45
- interleukin 10
- lamina propria
- T cell subsets
- T lymphocytes
Statistics from Altmetric.com
Human intestinal lamina propria T lymphocytes (LPL-T) are constantly exposed to a large number of luminal antigens, yet systemic immune responses do not develop. The mechanisms underlying this particular type of unresponsiveness in the intestinal compartment involve different levels of immune regulation. Firstly, antigen specific reactivity by LPL-T is down regulated as a result of impaired signal transduction through the T cell receptor (TCR)/CD3 complex.1-3 Mediators believed to be involved in reduced antigen reactivity of LPL-T are produced locally. These are small, non-protein molecules with oxidative properties.4 ,5Responses to stimulation through, for example, CD2 and CD28, are unaffected or even enhanced.1-3 ,6 Secondly, the surface densities of adhesion molecules such as CD54, CD58, and CD80 are reduced on mucosal macrophages.7 Lack of these co-stimulatory molecules could contribute to the inability of intestinal macrophages to induce T cell proliferation7 and thus prevent progression of antigen specific immune reactions. In addition, immunomodulation by cytokines could play a role in maintaining hyporesponsiveness in the gut. This notion is supported by the finding that interleukin (IL) 10 and IL-2 knockout mice develop chronic enterocolitis.8 ,9
Given its functional properties, IL-10, in particular, could contribute to the specialised state of the intestinal immune system. Firstly, IL-10 is known to suppress T cell proliferation either by acting directly on T lymphocytes or by interfering with T cell–antigen presenting cell interactions.10-15 Secondly, IL-10 down regulates gene expression and synthesis of inflammatory cytokines/chemokines, such as TNFα, IL-1, IL-6, IL-8,16-19 and MIP-1α20 by activated monocytes and macrophages; decreases surface expression of accessory molecules like CD80, CD86, and CD54 on human antigen presenting cells (monocytes, macrophages, dendritic cells)21-23; and induces antibody production by activated B lymphocytes.24 ,25 This means that particular features of the intestinal immune response could be related to the activity of IL-10. Conversely, characteristic immunological changes in chronic inflammatory bowel disease, such as enhanced mucosal IL-1β, IL-6, TNFα, and IFN-γ production26-28 could be explained by altered mucosal IL-10 homoeostasis.
To explore the regulation of IL-10 in the intestinal immune system, we investigated the capacity of isolated human LPL-T to produce IL-10 in comparison with autologous PBL-T. Our results indicate that LPL-T produce significantly more IL-10 than do PBL-T after activation. This cannot merely be explained by the different composition of T cell subsets in the lamina propria versus the blood, but is mainly a result of an enhanced responsiveness of LPL-T to CD2 triggering.
Reasons for the well documented increased CD2 sensitivity of LPL-T have not been elucidated as yet. Our results suggest that CD45, a tyrosine phosphatase physically associated with CD2 in T cells,29might be involved, as engagement of CD45 was found to produce a CD2 responsive state in PBL-T similar to that existing constitutively in LPL-T.
Methods
REAGENTS
CD2 monoclonal antibodies T112 1OLD4C1 and T113 1mono2A6 as well as CD4 monoclonal antibody 19THY5D7 were kindly provided by Drs S F Schlossman and E L Reinherz, Dana–Farber Cancer Institute, Boston, MA, USA. Anti-CD3 monoclonal antibody (OKT-3) was obtained from the American Type Culture Collection (Rockville, MD, USA). UCHL-1 (anti-CD45) was purchased from Ortho Diagnostics (Neckargmünd, Germany), and MEM-56 (anti-CD45) was kindly provided by Dr V Horejsi (Academy of Science, Prague, Czech Republic). B211/AICD45.1 (anti-CD45) and Eri-8/AICD8.1 (anti-CD8) were produced in our own laboratory.30 ,31 Interleukin 2 was a generous gift of Biotest (Dreieich, Germany).
PREPARATION OF T LYMPHOCYTES AND MONOCYTES
Colon specimens were obtained from patients undergoing bowel resection for colon cancer. The mucosal tissue used in this study was taken from the resection margin with the ‘new resection margin’ being microscopically normal. Lamina propria mononuclear cells were isolated according to a modified method of Bull and Bookman.32 The fresh tissue was washed extensively in HBSS without Ca2+ or Mg2+ and antibiotics (Gibco, Paisley, UK). The mucosa was dissected and incubated with DTT 1 mM (Sigma, St Louis, MO, USA) in HBSS without Ca2+ or Mg2+ for 15 minutes. After washing twice the mucosa was then cut into 2–4 mm pieces which were incubated in a shaking waterbath at 37°C with 0.7 mM EDTA (Sigma) in HBSS without Ca2+ or Mg2+ for 45 minutes. This incubation was repeated twice and followed by washing four times until the supernatant became clear. Subsequently, the pieces were incubated in a shaking waterbath at 37°C for 10–12 hours in a solution of RPMI 1640 (Gibco) with 2% fetal calf serum (FCS) (Gibco), 45 U/ml collagenase, 27 U/ml deoxyribonuclease I (Sigma), 2%l-glutamine, and antibiotics (Gibco). The cell suspension was separated from undigested tissue by filtration through a 60 mm nylon mesh (Fa. Eckert, Waldkirch, Germany) and washed in RPMI 1640 plus 2% FCS.
The cells were resuspended in 100% Percoll consisting of nine parts Percoll (Pharmacia, Uppsala, Sweden) and one part 10× phosphate buffered saline (PBS) (Seromed, Berlin, Germany). After centrifugation for 10 minutes at 1000 g, the layer between 67.5% and 44% was recovered and washed twice. These cells were then mixed with 30 μl of 5% sheep red blood cell (SRBC) suspension per 1×106 cells, spun at 700 rpm for five minutes and incubated for 45 minutes at room temperature to allow E-rosette formation. Subsequently, the pellet was gently resuspended and centrifuged on Ficoll–Hypaque for 20 minutes at 1000 rpm and at 1800 rpm. Supernatant and E-rosette negative cells were removed and the pellet was treated with hypotonic buffer to lyse SRBC. The E-rosette positive cells were washed three times and resuspended in RPMI 1640 plus 10% FCS and 2% l-glutamine for use.
Peripheral blood was taken during the operation. Peripheral blood mononuclear cells were isolated by Ficoll–Hypaque density gradient centrifugation. The isolated cells (5–10×106/ml) were incubated for two hours at 37°C in RPMI 1640 supplemented with 10% FCS and 2% l-glutamine on plastic Petri dishes (Greiner, Frickenhausen, Germany). Non-adherent cells were further processed for isolation of T cells. Adherent cells were harvested by using a rubber policeman (Costar, Cambridge, MA, USA), irradiated (50 Gy) and used as monocytes (PBMO).
ISOLATION OF T CELL SUBSETS
For negative selection of T cell subsets, PBL-T (107/ml) from healthy donors were incubated with 1 ml of UCHL-1 or MEM-56 supernatant for 30 minutes at 4°C. Following two washes the cells were incubated with goat–anti-mouse IgG microbeads (Milteny Biotec GmbH, Bergisch-Gladbach, Germany) for 15 minutes at 4°C. Magnetic activated cell sorting (MACS) was performed, following the manufacturer’s instructions.
For further isolation of CD4 and CD8 subsets the same procedure was applied using Eri-8 ascites and the 19Thy5D7 monoclonal antibody. The resulting T cell subsets were over 90% pure as shown by flow cytometric analysis.
PROLIFERATION ASSAY
Cells (5×104/well) were cultured in round bottomed microtitre plates (Costar) in RPMI 1640 supplemented with 10% FCS, 2%l-glutamine and antibiotics as well as reagents, in a total volume of 200 μl/well. Individual wells were pulsed with 1 μCi of [3H]-thymidine (Amersham, Braunschweig, Germany) at day 4 for 16 hours and then harvested on a cell harvester (Inotec, Wohlen, Switzerland). [3H]-thymidine incorporation was measured in a liquid scintillation spectrometer (Beckmann, Munich, Germany). Results of proliferation are shown as means of triplicate values (SD<15%).
PRODUCTION OF SUPERNATANT
Cells (2×105) were cultured in round bottomed microtitre plates (Costar) in RPMI 1640 supplemented with 10% FCS, 2% l-glutamine and antibiotics as well as reagents, in a total volume of 200 μl/well. After 60 hours, the supernatants were harvested, centrifuged to remove cells and frozen at −20°C until assayed.
IL-10 IMMUNOASSAY
The IL-10 content of the supernatants was determined by a cytokine specific ELISA (Serva, Heidelberg, Germany). The tests were performed according to the manufacturer’s instructions.
STATISTICAL ANALYSIS
The paired Student’s t test was used to compare IL-10 production by CD2 stimulated LPL-T and by PBL-T in the presence or absence of CD45 monoclonal antibody.
Results
Freshly isolated LPL-T and autologous PBL-T were incubated with, either: (a) medium; (b) CD2 monoclonal antibodies T112/3, in the presence or absence of SRBC; (c) CD3 monoclonal antibody OKT-3 coupled to Sepharose beads plus IL-2; or (d) anti-CD3 monoclonal antibody OKT-3 plus autologous irradiated blood monocytes (PBMO). Supernatants were harvested after 60 hours. T cell proliferation was measured in parallel.
As shown in table 1, neither LPL-T nor PBL-T released measurable amounts of IL-10 constitutively. Triggering via CD2 in the presence of SRBC consistently induced greater IL-10 release from LPL-T than from PBL-T. Differences in IL-10 production were most evident after CD2 ligation by monoclonal antibodies T112/3 or after stimulation via OKT-3 plus autologous monocytes. Relatively low IL-10 generation was induced after triggering through CD3, even in the presence of IL-2.
IL-10 production and cell growth of PBL-T and LPL-T in response to various stimuli
Parallel determination of T cell proliferation revealed that cell growth does not correlate with IL-10 production because the differences between the two cell populations were clearly less pronounced for proliferation than for IL-10 production. Moreover, differential IL-10 secretion within one cell population (LPL-T and PBL-T) also does not correlate with cell growth. Thus, T cell proliferation and IL-10 production are regulated individually.
Differential IL-10 release by PBL-T and LPL-T might result from the differing composition of T cell subsets in the two compartments: whereas the T cell population in the intestinal lamina propria consists almost exclusively of CD45RO+ T cells (∼90%),3 ,33 ,34the blood population contains approximately equal numbers of CD45RA+ and CD45RO+ T cells.3 ,35 ,36 To explore this possibility, we fractionated PBL-T of healthy volunteers into CD45RO+ and CD45RA+ subsets by negative selection and determined their IL-10 production in response to the same stimuli as used before. Table 2 shows that, irrespective of the mode of activation, only CD45RO+, but not CD45RA+, cells were capable of releasing IL-10 (the small amounts of IL-10 detectable after stimuation by OKT-3 plus monocytes probably originate from the monocytes). Lack of IL-10 production by CD45RA+ T cells cannot be attributed to general hyporesponsiveness of these cells as, for example, OKT-3 plus monocytes, which did not induce IL-10 release, caused vigorous proliferation of CD45RA+ T cells.
Differential regulation of IL-10 release and proliferation of activated CD45RO+ and CD45RA+ PBL-T
Further fractionation of the CD45RO+ and CD45RA+ populations into CD4+ and CD8+ subsets revealed that the CD45RO+CD4+ subset is the main source of IL-10 (table 3).
IL-10 is released mainly by CD45RO+ CD4+ PBL-T
However, the fact that LPL-T consist predominantly of CD45RO+ cells does not explain why IL-10 production in LPL-T was found to exceed noticeably that of PBL-T (compare table 1 with table 2).
Given that the tyrosine phosphatase CD45 was not only found to be physically associated with the CD2 receptor29 but also functionally important for its signalling capacity,30 the enhanced responsiveness of LPL-T to CD2 triggering raised the question of whether CD45 might contribute to this phenomenon. Indeed, as shown in fig 1, addition of a CD45 monoclonal antibody (AICD45.1/B211) did not only augment proliferation, but also IL-10 release by PBL-T, and to a similar extent by CD45RO+ PBL-T (table 4), thereby mimicking the state of CD2 responsiveness observed in LPL-T (at least at higher T112/3 monoclonal antibody concentrations). In contrast, CD2 induced IL-10 production in LPL-T is down regulated in the presence of B211 (average decrease, 33%; p=0.001, n=5) while cell proliferation is not affected or is slightly decreased. This suggests that CD45 is able to regulate not only CD2 promoted proliferation but also IL-10 production in PBL-T as well as LPL-T, and that this regulation differs between the two T cell populations.

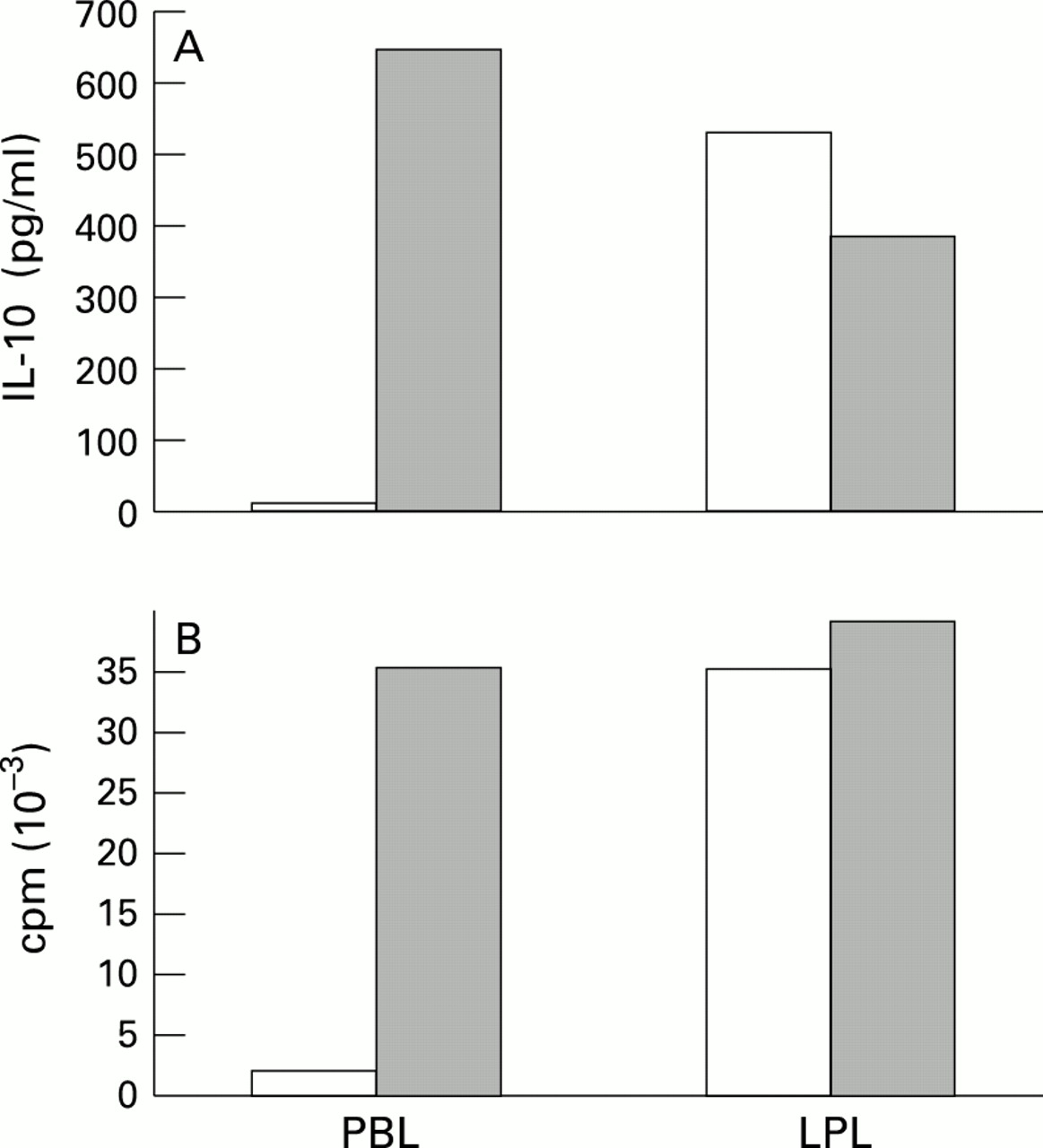{kind=link}
: PBL-T and LPL-T were activated by CD2 monoclonal antibodies T112 and T113 (1/1000 v/v) in the absence (open columns) or presence (shaded columns) of CD45 monoclonal antibody AlCD45.1/B211 (1/400 v/v). (A) Supernatants were harvested after 60 hours and IL-10 content (results expressed in pg/ml) were determined by using ELISA. (B) Cell growth was measured by incorporation of [3H]-thymidine after four days. Results are expressed as the mean cpm of triplicate cultures. SD < 20%. The results shown are representative of four different experiments.
Effect of anti-CD45 monoclonal antibody B211 on IL-10 release and proliferation of CD2 activated CD45RO+ and CD45RA+ PBL-T
Discussion
A possible role for IL-10 in intestinal immunoregulation has been proposed by several authors.9 ,37-40 Indeed, many features of the intestinal immune system, such as inhibition of T cell proliferation in response to TCR/CD3 stimulation,1-3decreased expression of adhesion molecules on mucosal macrophages7 or increased IL-2 receptor expression1 ,34 ,41 could be explained by the activity of IL-10 in the intestinal mucosa. Although it was shown that IL-10 is present in the intestinal mucosa,38 ,39 the cellular source of this cytokine and requirements for its production in the gut have not yet been analysed in detail. For this reason we investigated IL-10 production by LPL-T compared with PBL-T in response to various stimuli.
That T cells rather than monocytes are a major source of IL-10 is supported by the fact that monocyte enriched populations (70–80% monocytes) when activated produce less IL-10 than do purified T cells (>90%) under the experimental conditions used.
Our results demonstrate that there is neither significant constitutive IL-10 secretion in PBL-T (which is in accord with previous studies42-44) nor in LPL-T. Hence, unstimulated LPL-T are probably not a major source of IL-10 in the intestinal mucosa of healthy individuals. Instead, other cells such as epithelial cells38 must be responsible for the basic IL-10 secretion involved in mediating permanent ‘immuno-unresponsiveness’ in the normal mucosa. In contrast to quiescent LPL-T, activated LPL-T release large amounts of IL-10. As cells are activated during inflammation it is likely that the principal task of IL-10 production by LPL-T is not in maintaining basic immunosuppression but in controlling inflammation in the gut.
Compared with antigen receptor stimulation via OKT-3 plus IL-2, CD2 provides a strong signal for proliferation and IL-10 secretion in both LPL-T and PBL-T (the latter is in line with findings by Schwartzet al 45). Even so, stimulation induced IL-10 production is considerably higher in LPL-T than in PBL-T. This difference is not related solely to the presence of a higher percentage of IL-10 producing CD45RO+ T cells in the lamina propria compared with the blood. The actual basis for the increased IL-10 secretion by LPL-T seems to be their enhanced responsiveness to CD2 triggering. Upon addition of T112/3 monoclonal antibodies cell growth and IL-10 production are higher in LPL-T than in CD45RO+ PBL-T. Increased IL-10 generation by LPL-T in response to stimulation by OKT-3 in the presence of autologous irradiated monocytes may also be related to their increased responsiveness to CD2. CD45RO+ PBL-T are, however, predominantly triggered via the TCR/CD3 complex and therefore secrete less IL-10. LPL-T responses are more strongly influenced by the CD2 initiated accessory pathway (or that of other accessory receptors) and hence release greater amounts of IL-10.
Our findings confirm previous observations by ourselves1 ,2 ,5 and Targan et al,6 who demonstrated enhanced responsiveness of LPL-T to CD2 at the level of cytokine release, that is, TNF, IL-4, IL-2, and IFN-γ release. Regarding IL-4 secretion we have obtained very similar results as reported by this group. Thus, IL-4 production appears to be controlled in a fashion analogous to that of IL-10 production.
So far, the basis for the altered CD2 responsiveness of LPL-T remains obscure. Clearly, the present and earlier reported experimental findings1 ,2 are unrelated to treatment procedures in the course of LPL-T isolation as judged from control experiments with analogously treated PBL-T. A possible molecule involved in enhanced CD2 sensitivity is CD45, a membrane bound tyrosine phosphatase that is physically associated with CD2 in T cells.29 Various monoclonal antibodies against CD45 can increase T cell responsiveness to CD2 while concomitantly decreasing that to CD3.29 ,30 Indeed, we found that addition of anti-CD45 enhances PBL-T responsiveness to CD2 as reflected by increased proliferation and IL-10 production. Conversely, the same CD45 monoclonal antibody exerted a negative effect on IL-10 production by LPL-T. This could imply that, compared with PBL-T, in LPL-T either the functional status of CD45 is altered or the CD2 pathway is changed, leading to a modified response to CD45 engagement.
A possible role of IL-10 in the pathogenesis of chronic inflammatory bowel disease has been suggested by the finding that IL-10 knockout mice develop enterocolitis. Furthermore, topical application of IL-10 in patients with inflammatory bowel disease has been reported to down regulate pro-inflammatory cytokines and to improve clinical symptoms.37 Our results provide a basis for further investigations analysing potential alterations in IL-10 production and CD2 reactivity of LPL-T in chronic intestinal inflammation.
Acknowledgments
This work was supported by a grant from the Bundersministerium für Bildung und Forschung (BMBF)/Verbund KBF. We would like to thank Dr B Schraven for critically reading the manuscript. Also, we are grateful to Dr U Moebius for helpful discussion.
References
Linked Articles
- COMMENTARY