Article Text

Abstract
Background—Populations at low risk of colonic cancer consume large amounts of fibre and starch and pass acid, bulky stools. One short chain fatty acid (SCFA), butyrate, is the colon’s main energy source and inhibits malignant transformation in vitro.
Aim—To test the hypothesis that altering colonic transit rate alters colonic pH and the SCFA content of the stools.
Patients—Thirteen healthy adults recruited by advertisement.
Methods—Volunteers consumed, in turn, wheat bran, senna and loperamide, each for nine days with a two week washout period between study periods, dietary intake being unchanged. Before, and in the last four days of each intervention, whole gut transit time (WGTT), defaecation frequency, stool form, stool β-glucuronidase activity, stool pH, stool SCFA concentrations and intracolonic pH (using a radiotelemetry capsule for continuous monitoring) were assessed.
Results—WGTT decreased, stool output and frequency increased with wheat bran and senna, vice versa with loperamide. The pH was similar in the distal colon and stool. Distal colonic pH fell with wheat bran and senna and tended to increase with loperamide. Faecal SCFA concentrations, including butyrate, increased with senna and fell with loperamide. With wheat bran the changes were non-significant, possibly because of the short duration of the study. Baseline WGTT correlated with faecal SCFA concentration (r=−0.511, p=0.001), with faecal butyrate (r=−0.577, p<0.001) and with distal colonic pH (r=0.359, p=0.029).
Conclusion—Bowel transit rate is a determinant of stool SCFA concentration including butyrate and distal colonic pH. This may explain the inter-relations between colonic cancer, dietary fibre intake, stool output, and stool pH.
- bowel cancer
- colonic pH
- fibre
- intestinal transit
- pH
- short chain fatty acids
Statistics from Altmetric.com
There are theoretical grounds for believing that high colonic pH is relevant in the pathogenesis of colonic cancer.1-3When faecal pH has been measured in populations with a low risk of colonic cancer, it has been lower than in higher risk groups.2 ,4 ,5 Faecal and colonic pH are chiefly determined by a balance between the production and absorption of the weak base, ammonia, and of the weak acids, the short chain fatty acids (SCFA). Ammonia and SCFA are produced in the colon by bacterial fermentation of protein and carbohydrate, respectively, most of which are derived from the diet.6 Short chain fatty acids are produced by fermentation of carbohydrates that have escaped absorption in the small intestine, notably starch and non-starch polysaccharide (NSP, dietary fibre). As the colon is traversed SCFA are absorbed and the luminal pH gradually increases to around neutral.7This rise in pH fails to occur in rural Africans (who have a low risk of colonic cancer).8 The preservation of colonic acidity in Africans could be a result of the high polysaccharide content of their diet providing surplus substrate for SCFA production. Alternatively, it could be caused by the more rapid colonic transit reported to be characteristic of rural Africans,9 ,10delaying the absorption of SCFA, or to a combination of the two. If transit rate is important, it should be possible to mimic the African situation by accelerating transit by using laxatives.
One SCFA, butyrate, is the colon’s main source of energy and has several properties suggesting it has anti-neoplastic activity.11 Recently, it has been shown to be protective against malignant transformation in vitro.12 High colonic pH is likely to be associated with low levels of butyrate in the distal colon and thus increased predisposition to cancer. Although studies have provided conflicting data, increasing faecal SCFA concentration (and thus distal colonic concentration) has been shown to reduce distal colonic cell proliferation,13 and in dimethylhydralazine treated rats it reduces malignant transformation.14
We set out to test the hypothesis that colonic transit rate influences colonic pH and the SCFA, including butyrate, content of the stools, dietary intake being unchanged. We used wheat bran as well as senna to speed up transit. Bran was expected to have a substantial effect, as previous reports indicate that it lowers stool pH15 and increases faecal SCFA levels.16 ,17 Potential tumour promoters such as deoxycholic acid and steroids (that is, oestrogens) are dependent for their production on pH sensitive bacterial enzymes in the colon.1 ,18-20 These enzymes include β-glucuronidase. We therefore measured faecal β-glucuronidase activity to determine whether this also changed with alteration of colonic transit rate.
Subjects and Methods
Thirteen adults were recruited through advertisements placed in local hospitals. Volunteers were selected if they were omnivorous, healthy, on no medication, and not obese or pregnant. All were by chance non-smokers. The study was approved by the Research Ethics Committee of the United Bristol Healthcare Trust.
Raw wheat bran (Prewett’s, sieved to contain only particles 1.4–3.0 mm diameter) and senna tablets (Senokot, Reckitt and Coleman), were used to accelerate intestinal transit. Loperamide tablets (Imodium, Janssen Pharmaceuticals) were used to slow down transit. Each agent was taken for nine days and there were two to four week washout periods between agents to obviate any carryover effects. Each agent was taken in the maximum tolerated dose. Wheat bran was taken in divided doses with meals. The interventions were studied in a set order (wheat bran, senna, loperamide) because successful randomisation would have required an impracticably large number of volunteers to ensure a balanced distribution between the six possible orders.
Before and at the end of each intervention period volunteers were weighed and their height measured to calculate their body mass index and asked to complete a four day dietary record (two week days and two weekend days) using scales or household measures. Where insufficient dietary information was recorded the subjects were telephoned for clarification. The records were analysed for individual nutrients (total energy, total fibre (Southgate), insoluble non-starch polysaccharide (NSP), soluble NSP, total NSP, total fat, saturated fat, polyunsaturated fat, protein, carbohydrate, extrinsic sugar, and alcohol) using a computer program based on McCance and Widdowson’s food tables21 and the publications of Englyst et al.22 ,23
Before and at the end of each intervention period whole gut transit time (WGTT) was measured as a proxy for colonic transit by a modification of a published method.24 Twenty radio-opaque marker pellets contained within a capsule were swallowed (different shapes each day) on each of four consecutive mornings. The first two stools passed at least 24 hours after ingestion of the last set of markers were collected, flattened and radiographed. By counting the number of markers in each stool then applying the following formula the mean WGTT was calculated from the two stools.
WGTT (h) = (t1s1 +t2s2 ... +t8s8 )/(s1 + s2 ... + s8 )
where s = number of markers of a given shape in a stool sample, that is, 0–20, and t = time in hours since ingestion of this marker pellet to the passing of the stool. The subscripts 1–8 identify the four different shapes of marker pellet in the two stool samples. Thus, there are up to four types of pellets in each stool, identified by subscripts 1–4 in the first and 5–8 in the second stool.
Within 12 hours of passing, the stools were weighed and the second stool was liquidised, tested for pH with a Gelplas combination pH electrode probe, then frozen at −20°C for later β-glucuronidase activity analysis by using a modification of an established method.25 A pilot study had shown that stool pH did not vary significantly within 24 hours of defaecation.
Stool output per week was calculated as the mean weight of the two collected stools multiplied by the subject’s stated number of defaecations per week. Volunteers also completed a diary of their stool appearance, on a 1–7 scale, which is sensitive to transit time26 ,27 and of dates and times, enabling the calculation of interdefaecatory intervals (IDI).
The pH sensitive, radio-transmitting capsule (see below) was swallowed with the third day marker capsule. The capsule and markers were taken in the morning after an overnight fast, volunteers being asked not to eat or drink for the next hour.
The experimental agents were commenced immediately after completion of baseline assessments and continued until the on-treatment assessments of dietary intake and WGTT were completed.
pH RADIOTELEMETRY
The pH sensitive capsule comprises a transducer, battery, and radio transmitter sealed in a glass capsule7 (Remote Control Systems, Amersham, UK). The capsule was calibrated before each use in buffers at pH 9.1 and 4.0 at 37°C and the 90% response time (an indication of the battery’s reliability) was measured. This gives the time for the capsule to attain 90% of the total change in pH after transfer from one of the buffers to the other. After each use the capsule was cleaned and a further calibration check was done. If the pH of either buffer was now over- or under-recorded by ⩾0.2 pH units or the response time was greater than three seconds, the study was rejected and repeated.
The pH readings were downloaded to a computer for analysis using Flexisoft II ambulatory pH analysis software (Oakfield Instruments, Witney, UK). For each part of the colon a mean value was calculated. For analysis the following conventions were followed7: proximal colon, first four hours of colonic recording; distal colon, last four hours of recording; mid colon, four hours midway between the proximal and distal recording periods.
Short chain fatty acids were measured by gas–liquid chromatography on a packed column (Chromosorb WAW10% SP 1200/1% H3PO4) of diethyl ether extracts.28 We used β-methyl valeric acid as an internal standard and a mixed SCFA external standard. The temperature program was 110–150°C at 16°C/min with two minutes at 110°C and two minutes at 150°C. Calibration curves were obtained by chromatographing standard solutions at five different concentration levels. The calibration curves were prepared by plotting the area ratios (weight of active per weight of internal standard).
Data were assessed as parametrically or non-parametrically distributed by Ryan Joiner tests and bar histograms and were expressed as means with standard deviations or medians with interquartile ranges. Changes from baseline were assessed for significance by using paired two-tailed Student’s t and Wilcoxon rank sum tests as appropriate. Associations were assessed by using Spearman correlation coefficients. These were calculated on the pooled off-treatment readings to determine whether the artificial changes produced by the experiments had any counterpart in spontaneous variations.
Results
Of the 13 volunteers (three male, 10 female) who entered the study (table 1), 11 completed all the protocols. Two women with long basal WGTT measurements withdrew from the loperamide section of the study. Consumption of wheat bran averaged (SD) 28.3 (8.7) g/day. Table 2shows that the intended changes occurred in WGTT, namely reductions with bran and senna (greater with senna) and an increase with loperamide. These were reflected in higher stool output with bran and senna (again greater with senna) and lower stool output with loperamide, though the first two changes just escaped statistical significance. The expected changes also occurred in the frequency of defaecation and in stool form, and all these were significant (table2).
Demographic data and anthropomorphic measurements at entry into the study
Changes in transit time and defaecatory measurements (median, interquartile range) with wheat bran, senna and loperamide
In the proximal colon (mean baseline pH 5.21) there were no significant changes in pH with any of the three interventions (data not shown). In the mid colon senna induced a significant fall averaging 0.46 pH units but with bran there was only a non-significant trend in the same direction (table 3). In the distal colon senna again induced a substantial drop in pH, averaging 0.48 pH units, and here wheat bran too had a significant acidifying effect. Loperamide was without effect in both the mid and distal colon. Stool pH did not significantly change with any intervention though there were trends in the expected direction with senna and loperamide.
pH in the mid and distal colon and in the stool before and during treatment with wheat bran, senna and loperamide
Faecal SCFA concentrations were affected in a consistent manner but in opposing directions by senna and loperamide (table 4). Acetic, propionic, and butyric acid concentrations all rose substantially with senna. The biggest increase was with butyric acid which rose more than threefold. Loperamide caused substantial reductions in propionic and butyric acids; the fall with acetic acid just escaped statistical significance. Bran resulted only in non-significant trends, albeit in the expected direction. Because changes in WGTT with wheat bran were not as impressive as those induced by senna, we looked at the subjects consuming wheat bran who showed the greatest reduction in WGTT (n=6). Changes in stool acetic acid (158–249 μmol/g wet weight, p=0.015, 95% confidence interval (CI) 8.17–47.15), propionic (55–58 μmol/g wet weight, p=0.059, 95% CI 0.26–8.73) and butyric acid (43–81 μmol/g wet weight, p=0.59, 95% CI 2.3–13.9) were seen. Senna increased valeric acid concentrations, but there were no changes in the branched chain acids isobutyric and isovaleric acid (which were present at very low concentrations) with any of the interventions (data not shown).
Faecal SCFA concentrations before and during treatment (μmol/g wet weight)
Faecal β-glucuronidase activity (geometric mean, mmol/g/h) fell with wheat bran from 42 (95% CI, 29–61) to 28 (21–39) with the difference of the log10 transformed data being −1.49 (ratio of the geometric means −2.06 to −1.08; p=0.019). The fall in activity with senna and rise in activity with loperamide were not significant.
No changes were seen in dietary intakes between the baseline records and the records done during the interventions (excluding wheat bran supplements). The average (SD) baseline NSP intake was 13.0 (3.8) g/day equivalent to 19.3 (5.2) g/day of dietary fibre (Southgate).
In the pooled off-treatment data (n=37) there were significant associations between WGTT and faecal SCFA concentrations (fig 1,r=−0.511, p=0.001), between WGTT and faecal butyric acid concentrations (fig 2, r=−0.577, p<0.001) and between WGTT and distal colonic pH (fig 3, r=0.359, p=0.029). Distal colonic pH was significantly associated with stool butyrate concentration (r=−0.343, p=0.019) but not total SCFA concentration (r=−0.269, p=0.112). There was no significant association between stool pH and stool total SCFA or butyrate concentration.
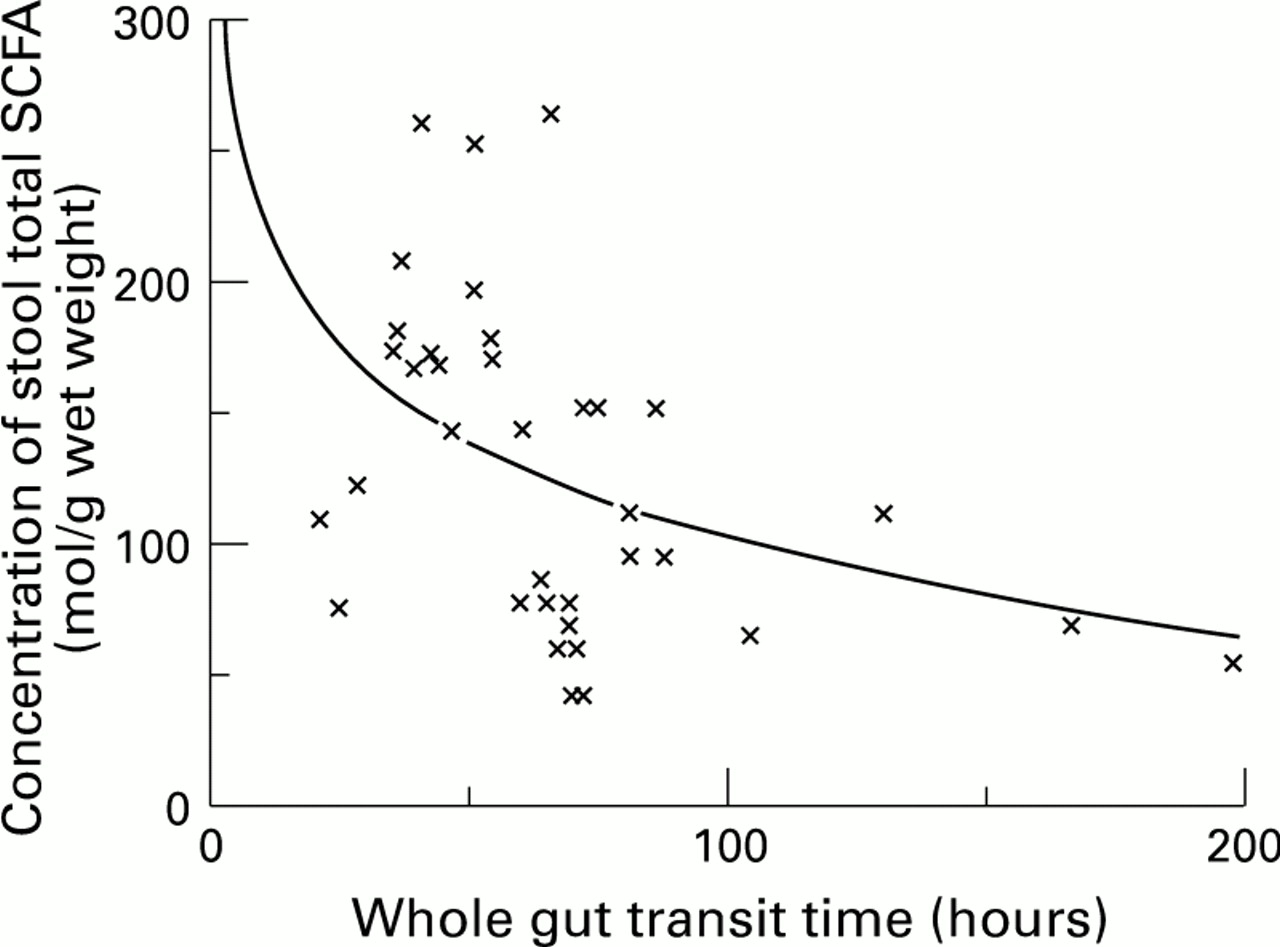
: The relation between whole gut transit time off-treatment and total stool SCFA concentration.

: The relation between whole gut transit time off-treatment and stool butyrate concentration.

{kind=link}
{kind=link}
{kind=link}
: The relation between whole gut transit time off-treatment and distal colonic pH.
Discussion
This study has shown that the physiochemical milieu within the colon can be profoundly altered simply by altering the rate of passage of intestinal contents. In our normal volunteers, accelerating transit with senna made the contents of the mid and distal colon more acid and increased SCFA concentrations in the stool, the greatest effect being on butyrate concentrations which tripled. Slowing down transit with loperamide reduced SCFA concentrations but had little if any effect on pH. Do these experimental phenomena have any relevance to real life? That they may do is suggested by the correlations we found in our subjects off treatment. Whole gut transit time, a proxy for colonic transit time, was significantly correlated with faecal SCFA concentrations and, specifically, with butyrate concentration. These correlations have been recently confirmed by El Oufir et al.29 It was also, albeit weakly, correlated with pH in the distal colon. Since the baseline measurements of WGTT, stool weight and stool form were similar in our volunteers to those in the general population,26 ,30 ,31 while their colonic pH was similar to those in other normal volunteers,7 it is perhaps reasonable to extrapolate from our findings to the population at large.
Undoubtedly many factors contribute to the aetiology of bowel cancer but there are compelling arguments to incriminate high intraluminal pH and low butyrate concentrations. This being so, the present data imply that intestinal transit rate could have a significant role. The factors determining intestinal transit rate are not well understood. Transit time varies notably within and between individuals, even on a constant diet,32 ,33 some of this variation being explained by psychological factors.34 Transit time is slower in women than men.26 ,30 ,35 An epidemiological study in Bristol identified oral contraceptive use as a transit slowing factor and alcohol use and dietary fibre intake as transit speeding factors.36 All this suggests that there may be scope for reducing the risk of bowel cancer by altering the transit time of individuals and populations.
The obvious and “natural” way to speed up transit through the intestine is to increase the intake of dietary fibre, especially wheat fibre which is particularly effective.37 It was for this reason that we studied the effect of wheat bran. We expected to find substantial falls in colonic pH and rises in faecal SCFA concentrations because bran and other sources of dietary fibre not only speed up transit but also provide substrate for bacterial production of SCFA. In the event, we found a modest drop in the pH of the distal colon but no rise in faecal SCFA concentrations (albeit trends in that direction). Changes in WGTT, stool output, interdefaecatory intervals, and stool form were less impressive than for senna or loperamide. One reason may be that our volunteers failed to take enough bran. The average dose of 28 g daily was as much as in most studies and as much as most people can tolerate. However, it is possible that some of our volunteers took less bran than they reported (despite our weighing their unused bran to calculate their daily consumption) or that, by chance, they were unusually insensitive to the effect of bran. On the basis that 1 g of fibre from raw wheat bran adds 7.2 g to the weight of the faeces,37 our volunteers, who ingested 28 g of bran daily, providing 11.2 g of fibre, should have increased their stool output by 81 g/day. In fact, median faecal weight rose to only 34 g/day. It is possible that the seven day study period was too short for the effect of bran to become fully apparent. In another bran feeding study faecal SCFA concentrations had increased after six weeks but not after two weeks, possibly because of the need for colonic bacteria to adapt to their new substrate.16 Others have found that in certain people bran fails to produce the expected rise in stool weight38 or fall in transit time.39 Our findings with bran, therefore, may not accurately reflect the usual ability of wheat fibre to modify colonic function. However, we are not the first to report that faecal pH is unaffected by bran.4 ,40 Stephen et al 40 found faecal pH to be inversely related to transit time (which we failed to do here, but have reported in other studies41). In humans, cellulose has been shown to lower stool pH while pectin failed to do so, probably because pectin is rapidly fermented and the SCFA are all absorbed, whereas cellulose is poorly fermented (fermentability is not a factor in explaining fibre’s laxative effects37 ,42) and, unlike pectin, accelerates colonic transit43 enabling the SCFA produced to reach the distal colon. Interestingly, the subgroup of volunteers taking wheat bran, and who had the largest reduction in WGTT, had a more pronounced rise in faecal SCFA concentrations.
The failure of stool total SCFA concentration to correlate significantly with stool pH or distal colonic pH (though it did with stool butyrate) was surprising, as was the failure of loperamide to raise distal colonic and stool pH although it lowered faecal SCFA. Presumably, other acids and bases present in the stool are important determinants of pH. These are known to include lactate, sulphate, tartrate, succinate, and bile acids among the acids, while ammonia is the chief base.6 There is a complex interplay between dietary sources of these substances, the rate at which acids and ammonia are produced by bacterial metabolism, interactions between them, and their absorption from the colon, but it is likely that many of these processes are affected by transit through the intestine. Although our study suggests alteration of colonic transit time influences the balance between production and absorption of SCFA we are unable to comment specifically on whether the rate and extent of fermentation is altered specifically, as this may have implications on energy salvage of foods from the colon. One very unlikely caveat is that loperamide and/or senna may alter the transport rates of SCFA or the faecal bacterial flora so influencing faecal SCFA. Further studies would be needed to exclude these remote possibilities.
Bacterial enzymes are involved in the metabolism of a wide range of compounds, with the subsequent production of carcinogens.44 One such enzyme is the pH dependent β-glucuronidase, which as well as being responsible for deconjugating glucuronides (for example, oestrogens) also deconjugates benzo-(a)-pyrene glucuronides and other potential carcinogens to more toxic products. Previous work has also shown a decrease in β-glucuronidase activity with an increase in dietary fibre45-47 and lower levels in vegetarians compared with omnivores.45 Presumably other pH sensitive enzymes were similarly affected. One such enzyme is the bile acid 7α-dehydroxylase which catalyses the formation of deoxycholic acid from cholic acid. In experiments similar to the present ones we showed that deoxycholate concentrations in bile were reduced by speeding up intestinal transit and increased by slowing down transit, with parallel changes in the cholesterol saturation of gall bladder bile and, therefore, in the risk of gallstone formation.24 Thus, it is possible that there are far-reaching metabolic effects of changes in intestinal transit time as well as changes in the risk of bowel cancer.
The failure of any intervention to alter pH in the proximal colon is not surprising as pH here is likely to depend mostly on the arrival of unabsorbed carbohydrates and their rapid fermentation by caecal anaerobes.48
A relatively novel feature of this study is that we used subjects’ observations of their stool form as an assessment of colonic function. Previous cross-sectional studies have established that the Bristol stool form scale is sensitive to transit time.26 ,27 ,49This is the first interventional study to show that change in transit time is accompanied by change in stool form, which further validates the scale as a simple proxy for transit time.
The decrease in SCFA concentration seen in stools when loperamide was taken suggests that any diet or lifestyle changes that lead to constipation can reduce the concentration of SCFA in the distal colon. This may help to explain the prevalence of distal colonic cancers in Western countries. The data from volunteers taking wheat bran and senna suggests that anything which increases intestinal transit rate will beneficially increase the concentration of butyrate in the distal colon independently of dietary intake of fermentable substrate.
Acknowledgments
We should like to thank the Kellogg Company of Great Britain and the trustees of the United Bristol Health Care Trust for their generous support of this work. We would also like to express our appreciation to Dr David Evans, GI Research Unit, The London Hospital, UK, for help in using the pH telemetry system. The technical assistance of Dr Christine Edwards and the staff of the Department of Human Nutrition, Glasgow University, York Hill Hospital, Glasgow, is gratefully acknowledged.