Article Text

Abstract
Background —Helicobacter pyloristrains possessing the cagA gene are thought to induce interleukin 8 (IL-8) in gastric mucosa. However, it is still unclear whether a relation exists between the cagA gene and the expression patterns of cytokines other than IL-8.
Aims —To investigate the relation between the cagA gene and the production of various cytokine proteins using an enzyme linked immunosorbent assay (ELISA).
Patients and methods —In 184 patients, the cagA gene was detected by polymerase chain reaction (PCR), and levels of production of IL-1β, IL-6, IL-7, IL-8, IL-10, and tumour necrosis factor α (TNF-α) in antral biopsy specimens were measured by ELISA.
Results—Mucosal levels of IL-1β, IL-6, IL-8, and TNF-α were significantly higher in H pyloripositive than in H pylori negative patients. Furthermore, the mucosal levels of IL-1β and IL-8 were significantly higher in specimens infected with cagApositive strains than in those infected with cagAnegative strains. In H pylori positive patients, the mucosal level of IL-8 was closely correlated with that of IL-1β (p<0.0001), and the mucosal level of IL-6 was closely correlated with that of TNF-α (p<0.0001).
Conclusion—These findings suggest that the ability to induce cytokines differs among the strains;cagA + strains induce various kinds of cytokines and may cause severe inflammation, whereascagA − strains induce IL-8 and IL-1β only weakly and may cause only mild inflammation. However, as most patients infected with the cagA + strains have gastritis, these strains may not be equivalent to ulcerogenic strains.
- cytokines
- Helicobacter pylori
- cagA gene
- interleukin 8
- interleukin 1β
Statistics from Altmetric.com
Helicobacter pylori associated gastritis is characterised by severe infiltration of neutrophils and mononuclear cells in the gastric mucosa.1 Accumulation and activation of these cells has been suggested to be induced by the local production of cytokines. Recent studies have demonstrated that mucosal levels of interleukin (IL) 1β, IL-6, IL-8, and tumour necrosis factor α (TNF-α) were significantly higher in H pylori positive patients than in H pylori negative patients using enzyme linked immunosorbent assay (ELISA).2-4 We have also studied cytokine expression patterns in gastric mucosal biopsy specimens using reverse transcription polymerase chain reaction (RT-PCR) and found that the levels of IL-6, IL-7, IL-8, IL-10, and TNF-α mRNA expression were significantly higher in H pylori positive than in H pylori negative gastritis.5 ,6 Taken together, these results and those of our previous studies using ELISA and RT-PCR suggest that IL-1β, IL-6, IL-7, IL-8, IL-10, and TNF-α play important roles in gastric inflammation caused by H pylori infection.
Furthermore, several potential virulence factors derived from H pylori are also considered to attract or activate neutrophils and mononuclear cells. Of these, an immunodominant 120–130 kDa antigen termed CagA has been well investigated.7 ,8Crabtree et al 9 reported that mucosal immune recognition of CagA was strongly associated with epithelial neutrophil infiltration. Recently, Crabtree et al 10 ,11found that CagA/cagA gene positive strains stimulated higher levels of IL-8 release than CagA/cagA gene negative strains using gastric epithelial cell lines. In our previous study,6 we also found that IL-8 mRNA expression was significantly more common in gastric mucosal biopsy specimens fromcagA gene positive gastritis than those fromcagA gene negative gastritis using RT-PCR. Recently, however, Sharma et al 12 demonstrated that there was no decrease in IL-8 induction by isogenic strains with a mutant cagA gene disrupting CagA expression. Tummuruet al 13 reported that the picBgene, which is present upstream of the cagA gene, was essential for induction of IL-8 from gastric epithelial cells in vitro. Therefore, CagA/cagA gene may be a phenotypic marker for induction of IL-8. Strains possessing the picB gene, however, have also been reported to express the cagA gene in vivo.13
It therefore seems that cagA gene positive strains (cagA gene positive and picB gene positive) have the ability to induce IL-8 release from gastric mucosa. However, it is still unclear whether a relation exists between cagAgene positive strains and the expression patterns of cytokines other than IL-8. It is important to investigate the relation between thecagA gene and the production of cytokines in vivo.
The present study was designed to examine the production of various cytokines in gastric antral biopsy specimens using quantitative ELISA with a large patient group, and to determine the presence of thecagA gene by PCR. We then determined which factors play important roles in H pylori associated gastritis.
Methods
PATIENT POPULATION
A total of 184 patients with dyspepsia (97 men and 87 women; age range 23–80 years, mean 56.7 years), none of whom had received non-steroidal anti-inflammatory drugs, proton pump inhibitors, or antibiotics within the previous three months, participated in this study. Endoscopic findings in the patients were as follows: normal mucosa, 20 patients; chronic gastritis without ulcer, 78 patients; gastric ulcer, 57 patients; and duodenal ulcer, 32 patients. Patients with gastric carcinoma were excluded from the study. We chose only those patients at the active to healing stage, without an ulcer scar. Seventy two patients were smokers. Informed consent was obtained from all patients and the protocol was approved by the ethical committee of Kyoto Prefectural University of Medicine.
Five biopsy specimens were taken from the antrum, using the same size forceps, from similar topographical sites at each endoscopy; two were used for histological examination, one for H pyloriculture, and two for cytokine measurement. In addition, three biopsy specimens were taken from the corpus: two for histological examination and one for H pylori culture. The presence of H pylori was determined by culture and histological examination. Patients were classified as H pylori positive if at least one of the examinations gave a positive result.
HISTOLOGY
The biopsy specimens were embedded in paraffin wax and stained with haematoxylin and eosin and Giemsa stains. Specimens were examined without knowledge of the experimental results. The histological severity of gastritis was graded from normal to severe based on the degree of mononuclear cell (MNC) and polymorphonuclear leucocyte (PMN) infiltration according to the Sydney system.14 The density of H pylori was graded as absent, scant, moderate, and heavy colonisation also according to the Sydney system.14
H PYLORI CULTURE
The biopsy specimens were spread with an applicator and plated on Skirrow agar medium containing 8% (v/v) horse blood, vancomycin (10 μg/ml), polymyxin B (2.5 U/ml), and trimethoprim (5 μg/ml) and incubated at 37°C under microaerophilic conditions (Campy-Pak Systems, BBL, Cockeysville, Maryland, USA) for up to five days. The organisms were identified as H pylori by Gram staining, colony morphology, and positive oxidase, catalase, and urease reactions. Multiple colonies on the agar were collected together and cultured again at 37°C under microaerophilic conditions in brain heart infusion broth (Nissui Co. Ltd, Osaka, Japan) containing 5% (v/v) horse serum for up to four days. Aliquots of 1 × 108 cfu/ml were stored at –80°C in brain heart infusion broth containing 15% (v/v) horse serum and 15% (v/v) glycerol.
PREPARATION OF GENOMIC DNA FROM CLINICAL ISOLATES
The culture medium was centrifuged for five minutes at 10 000 g and the bacterial pellets were washed three times with phosphate buffered saline (PBS; pH 7.4). The washed pellets were each suspended in 200 μl distilled water and boiled for 10 minutes. The supernatants, obtained by centrifugation (10 000 g for five minutes), were stored at –20°C until use as PCR templates.
POLYMERASE CHAIN REACTION FOR DETECTION OF THEcagA GENE
Bacterial DNA was denatured by heating to 95°C for 10 minutes and cooled on ice for five minutes. Bacterial DNA (5 μl) was added to 50 μl reaction mixtures containing 5 μl 10× PCR reaction buffer (500 mM KCl, 100 mM Tris-HCl (pH 8.8), 15 mM MgCl2, 1% Triton X-100, 200 mM of each dNTP (Pharmacia Biotech AB, Uppsala, Sweden), 200 nM of each primer, 1.0 unit Taq DNA polymerase (Boehringer, Mannheim, Germany), and H2O). Primers (5′-GATAACAGGCAAGCTTTTGAGG-3′ and 5′-CTGCAAAAGATTGTTTGGCAGA-3′) were used to amplify a 349 bp product from the middle of the cagA gene.8Primers (5′-GCGCAATCAGCGTCAGGTAATG-3′ and 5′-GCTAAGAGATCAGCCTATGTCC-3′) specific for H pylori16S rRNA were used as a positive control for the presence of H pylori.15 Amplification was performed using a Takara PCR Thermal Cycler 480 (Takara Biomedicals, Otsu, Japan) for 30 cycles, each of which consisted of one minute at 95°C for denaturation, one minute at 50°C for annealing, and one minute at 72°C for extension. The final cycle included an extension step for seven minutes at 72°C to ensure full extension of the product. Aliquots (10 μl) of each PCR product were analysed by electrophoresis on 1.5% agarose (Dojin Chemical Co. Ltd, Kyoto, Japan) gels containing ethidium bromide and the bands were examined under ultraviolet light for the presence of the amplified DNA.
CYTOKINE MEASUREMENT
For cytokine measurement, two biopsy specimens were immediately placed in 2.0 ml PBS (pH 7.4), frozen on dry ice, and stored at –80°C until use. Samples were homogenised using a tissue homogeniser (Kontes, Vineland, New Jersey), and aliquots of homogenate supernatants, obtained by centrifugation (10 000 g for 10 minutes), were assayed for total protein by a modified Lowry method. The mean concentration of total protein was 0.57 mg/ml (range 0.51–1.76 mg/ml), and the supernatants diluted to 0.50 mg/ml total protein were frozen at –80°C until assay. In some cases, biopsy specimens were too small to enable measurement of all cytokines.
IL-1β, IL-6, IL-7, IL-8, IL-10, and TNF-α in biopsy homogenate supernatants were measured by ELISA using commercially available assay kits (IL-1β, IL-6, IL-7, IL-8, and TNF-α, Research and Diagnostic Systems, Minneapolis, Minnesota; IL-10, BioSource International, Camarillo, California, USA). These assays use the quantitative immunometric sandwich enzyme immunoassay technique. Amplification systems were used to increase the sensitivity, using the alkaline phosphatase reaction for IL-1β, IL-6, IL-7, and TNF-α and using the streptavidin peroxidase reaction for IL-10. These assays were performed in duplicate according to the manufacturer’s instructions. In our laboratory, the ELISA sensitivities of IL-1β, IL-6, IL-7, IL-8, IL-10, and TNF-α were approximately 0.125, 0.150, 0.25, 10.0, 0.50, and 0.50 pg/ml, respectively. Interassay variation of IL-1β, IL-6, IL-7, IL-8, IL-10, and TNF-α was less than 10.5%, 11.0%, 11.5%, 10.0%, 12.5%, and 11.0%, respectively. Intraassay variations of these cytokines were all less than 12.5%.
STATISTICS
The mucosal levels of IL-1β, IL-6, IL-7, IL-8, IL-10, and TNF-α were expressed as pg/mg biopsy protein. Statistical analysis was performed using the Mann-Whitney U test for non-parametric data. Correlation coefficients were calculated by the Spearman rank test. A p value of less than 0.05 was accepted as statistically significant.
Results
EXPRESSION OF cagA GENE AND HISTOLOGICAL AND ENDOSCOPIC FINDINGS
Of the 184 patients examined, 141 (76.6%) were H pylori positive by culture or histological staining (table 1). Ten patients were positive for H pylori only by histological staining. Of the 133 patients with positive culture forH pylori, all (100%) were positive for 16S rRNA and 120 (90.2%) were positive for the cagA gene. The positive rate for the cagA gene was 100% in patients with duodenal ulcer and 86.8% in patients with chronic gastritis without ulcer.
Expression of the cagA gene and endoscopy findings
Antral biopsy specimens infected with H pylori had significantly more severe infiltration of PMNs (p<0.0001) and MNCs (p<0.0001) than those without H pylori (table 2). Furthermore, specimens infected with cagA +strains had significantly more severe infiltration of PMNs (p<0.01) and MNCs (p<0.01) than those infected withcagA − strains. In H pyloripositive specimens, PMN and MNC infiltration was significantly more severe in patients with duodenal ulcer than in those with chronic gastritis without ulcer (data not shown).
Expression of the cagA gene and histology findings
PRODUCTION OF CYTOKINES AND H PYLORI INFECTION
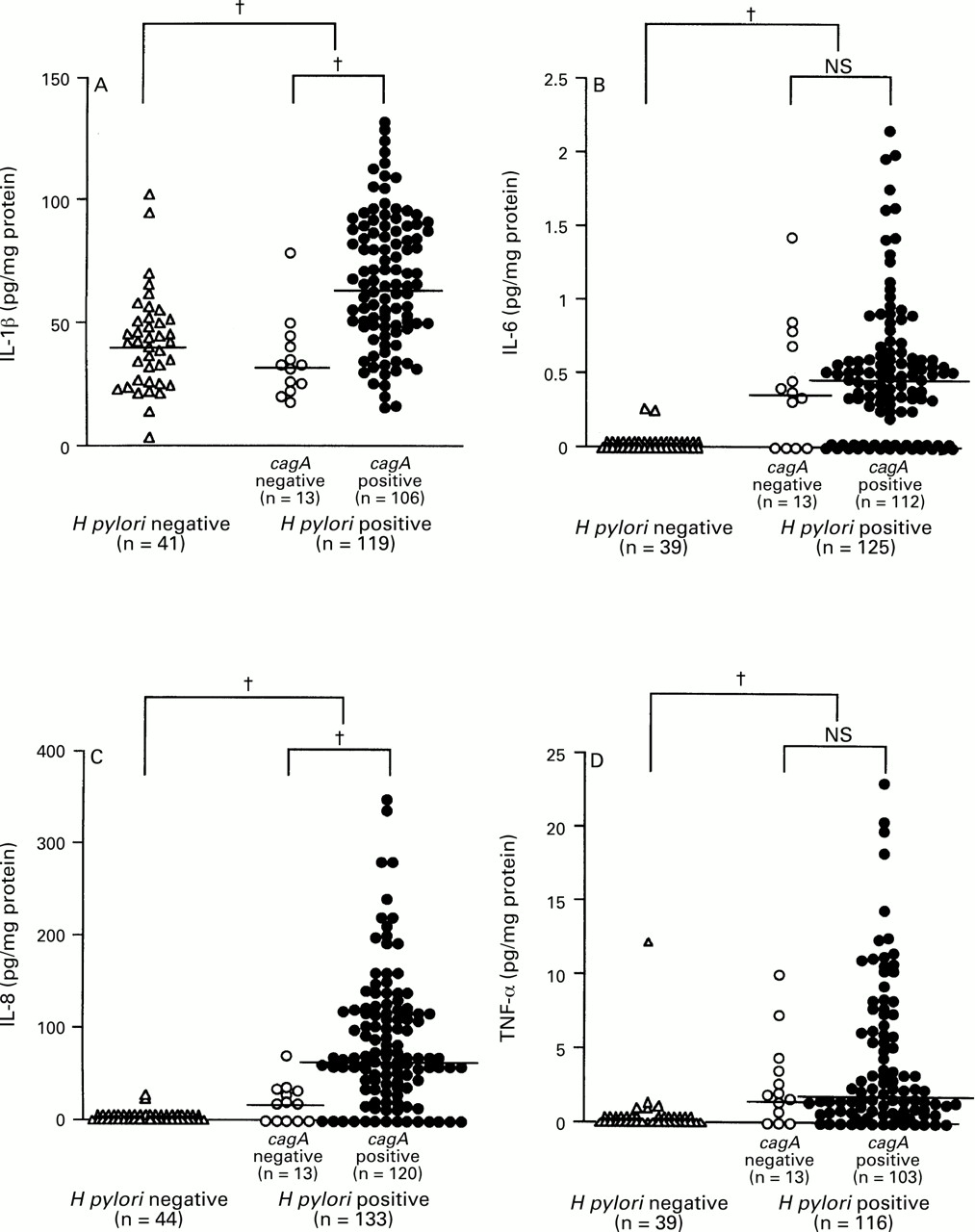
The mucosal levels of IL-1β, IL-6, IL-8, and TNF-α were significantly higher in H pylori positive than in H pylori negative specimens (IL-1β: median 62.0 pg/mg, range 15.4–130.0, versus median 42.3 pg/mg, range 3.6–102.7, p<0.0001; IL-6: median 0.50 pg/mg, range 0.00–2.15, versus median 0.00 pg/mg, range 0.00–0.25, p<0.0001; IL-8: median 60.9 pg/mg, range 0.0–346.5, versus median 0.0 pg/mg, range 0.0–20.5, p<0.0001; TNF-α: median 1.70 pg/mg, range 0.00–22.98, versus median 0.0 pg/mg, range 0.0–12.07, p<0.0001) (fig 1). IL-6, IL-8, and TNF-α were detected in only two (4.5%), two (4.5%), and five (11.3%) H pylori negative specimens, respectively. There were eight H pylori negative specimens in which at least one of IL-6, IL-8, or TNF-α was detected and seven of these had an MNC infiltrate. The detection rate of IL-1β, however, was 100%, irrespective of H pylori infection.
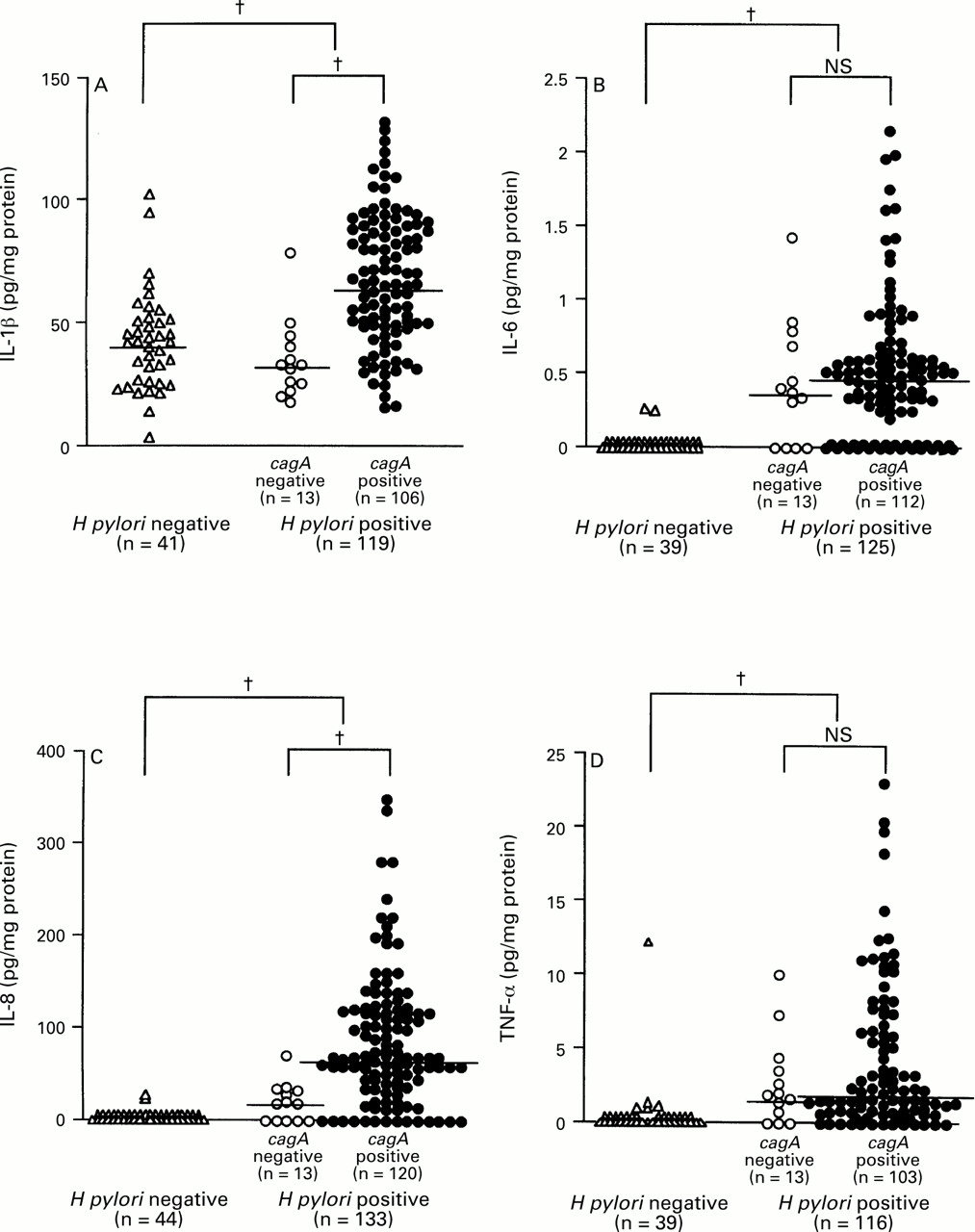
: Production of (A) IL-1β, (B) IL-6, (C) IL-8, and (D) TNF-α and H pylori infection. Large brackets indicate the comparison between H pylori positive and negative specimens and small brackets the comparison between cagA+ and cagA– specimens. Bars indicate median values for each group. †p<0.0001 by Mann-Whitney U test.
The mucosal levels of IL-7 and IL-10 were not increased in H pylori positive compared with H pylori negative specimens (IL-7: 7.50 pg/mg, range 2.83–22.46, versus 6.80 pg/mg, range 3.26–18.66; IL-10: 32.7 pg/mg, range 9.3–158.6, versus 29.0 pg/mg, range 10.6–89.6). Two specimens with mucosal levels of IL-8 over 300.0 pg/mg (340.1 and 346.5 pg/mg) also had high levels of IL-10 (130.4 and 100.4 pg/mg). However, the mucosal level of IL-10 was higher in H pylori positive specimens with a sensitivity under the detection level of IL-8 (median 37.8 pg/mg, range 12.6–124.6) than in H pylori positive specimens overall. The specimen with the highest level of IL-7 had severe metaplastic mucosa in the antrum.
The mucosal levels of IL-1β and IL-8 were significantly higher incagA + than in cagA −specimens (IL-1β: median 65.5 pg/mg, range 15.4–130.0, versus median 32.6 pg/mg, range 17.6–78.5, p<0.0001; IL-8: median 63.0 pg/mg, range 0.0–346.5, versus median 16.7 pg/mg, range 0.0–68.8, p<0.0001) (fig 1). On the other hand, the mucosal levels of IL-6 and TNF-α in cagA + specimens were almost the same as those in cagA – specimens (IL-6: median 0.49 pg/mg, range 0.00–2.15, versus median 0.36 pg/mg, range 0.00–1.42; TNF-α: median 1.54 pg/mg, range 0.00–22.98, versus median 1.85 pg/mg, range 0.00–9.96).
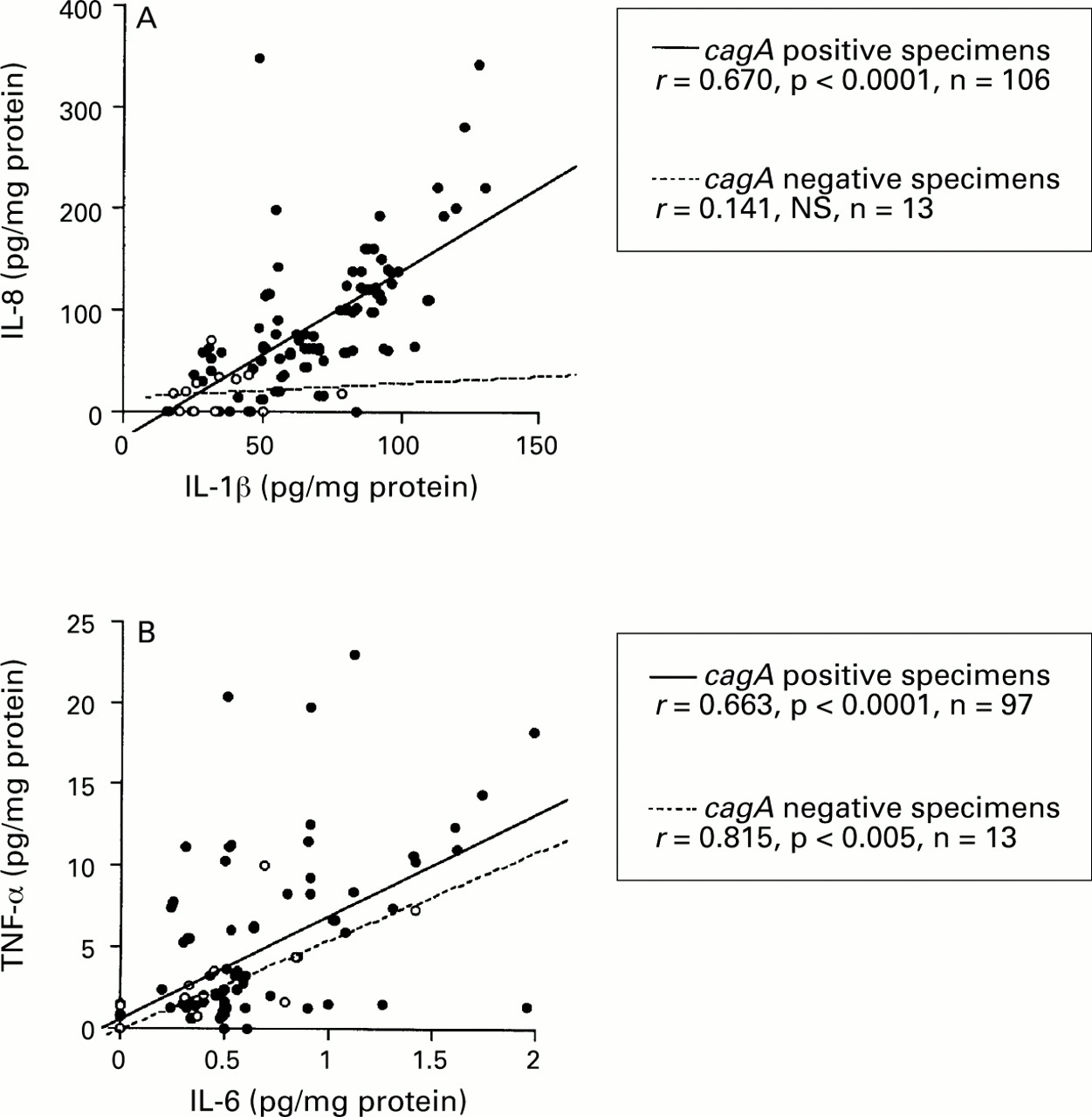
In cagA + specimens, the mucosal level of IL-8 was significantly correlated with that of IL-1β (r=0.670, p<0.0001), and the mucosal level of IL-6 was significantly correlated with that of TNF-α (r=0.663, p<0.0001) (fig 2). There were no significant correlations among the other combinations of cytokines (data not shown). IncagA – specimens, however, the mucosal level of IL-8 was not correlated with that of IL-1β, although the mucosal levels of IL-6 and TNF-α were significantly correlated (r=0.815, p<0.005).

: Correlation between (A) IL-1β and IL-8, and (B) IL-6, and TNF-α production. Filled circles, cagA+specimens; open circles, cagA− specimens. Correlation coefficients were calculated with the Spearman rank test.
PRODUCTION OF CYTOKINES AND HISTOLOGICAL AND ENDOSCOPIC FINDINGS IN PATIENTS WITH H PYLORI INFECTION
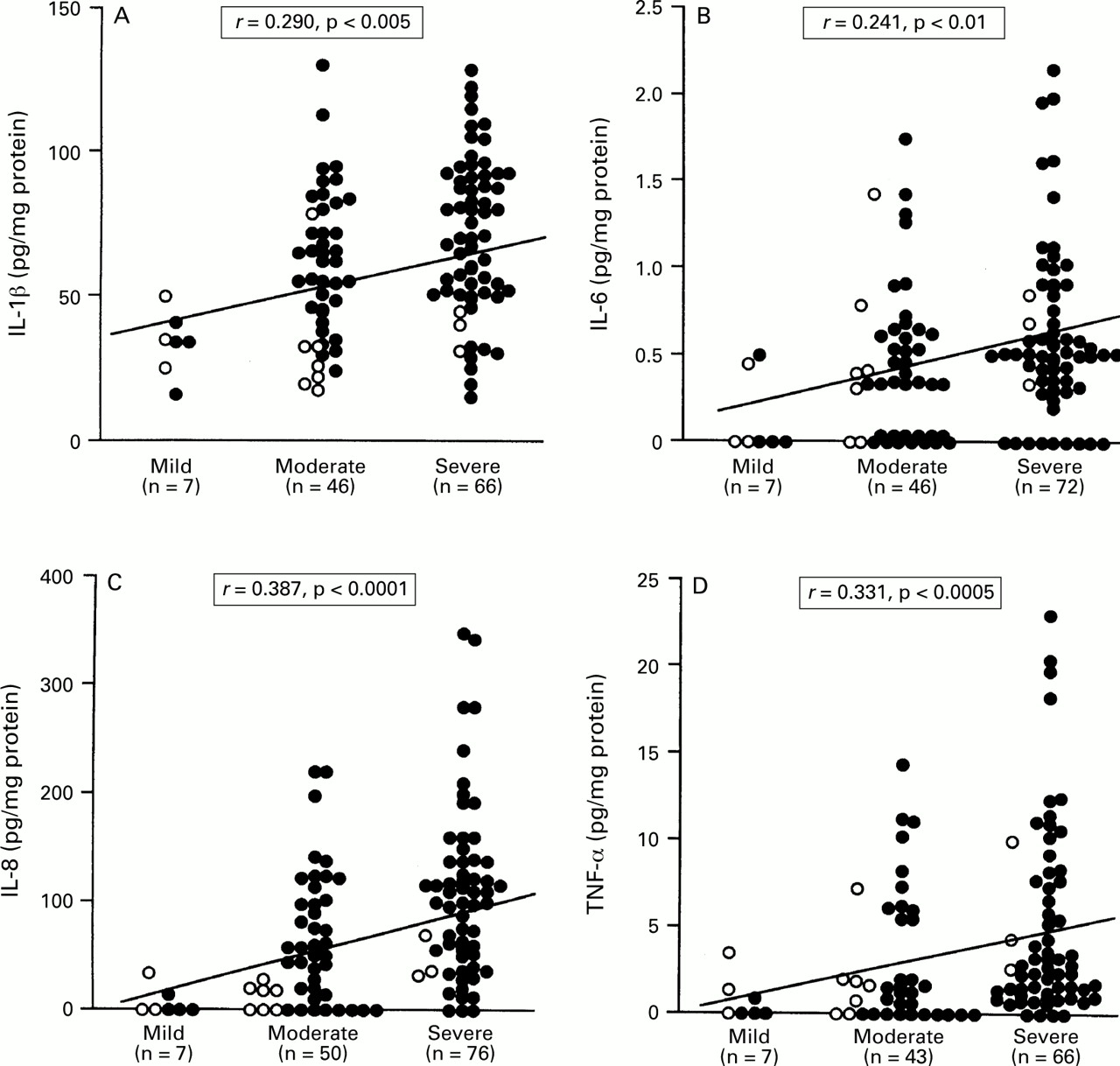
We examined the relation between cytokine production and histological/endoscopic findings in the antral biopsy specimens from patients with H pylori infection. The mucosal levels of IL-1β, IL-6, IL-8, and TNF-α were significantly correlated with the infiltration of MNC (IL-1β: r=0.290, p<0.005; IL-6:r=0.241, p<0.01; IL-8: r=0.387, p<0.0001; TNF-α: r=0.331, p<0.0005) (fig 3) and PMN (IL-1β:r=0.352, p<0.0005; IL-6: r=0.250, p<0.01; IL-8: r=0.582, p<0.0001; TNF-α: r=0.301, p<0.005) (fig 4). The mucosal levels of IL-7 and IL-10, however, were not correlated with the infiltration of MNC and PMN.

: Relation between the production of (A) IL-1β, (B) IL-6, (C) IL-8, and (D) TNF-α and MNC infiltration in patients with H pylori infection. Filled circles, cagA+specimens; open circles, cagA− specimens. Correlation coefficients were calculated with the Spearman rank test.

: Relation between the production of (A) IL-1β, (B) IL-6, (C) IL-8, and (D) TNF-α and PMN infiltration in patients with H pylori infection. Filled circles, cagA+specimens; open circles, cagA− specimens. Correlation coefficients were calculated with the Spearman rank test.
The mucosal level of IL-8 was significantly higher in patients with duodenal ulcer (median 114.5 pg/mg, range 0.0–346.5) than in those with gastric ulcer (median 57.4 pg/mg, range 0.0–279.6, p<0.05) and chronic gastritis without ulcer (median 58.0 pg/mg, range 0.0–190.7, p<0.005) (fig 5). The mucosal level of IL-6 was significantly higher in patients with duodenal ulcer (median 0.55 pg/mg, range 0.00–1.62) than in those with chronic gastritis without ulcer (median 0.36 pg/mg, range 0.00–1.42, p<0.05).

: Production of (A) IL-1β, (B) IL-6, (C) IL-8, and (D) TNF-α and endoscopic findings in patients with H pylori infection. Filled circles, cagA+ specimens; open circles, cagA− specimens. Bars indicate median values for each group. *p<0.05; †p<0.005 by Mann-Whitney U test.
PRODUCTION OF CYTOKINES AND THE DENSITY OF H PYLORI
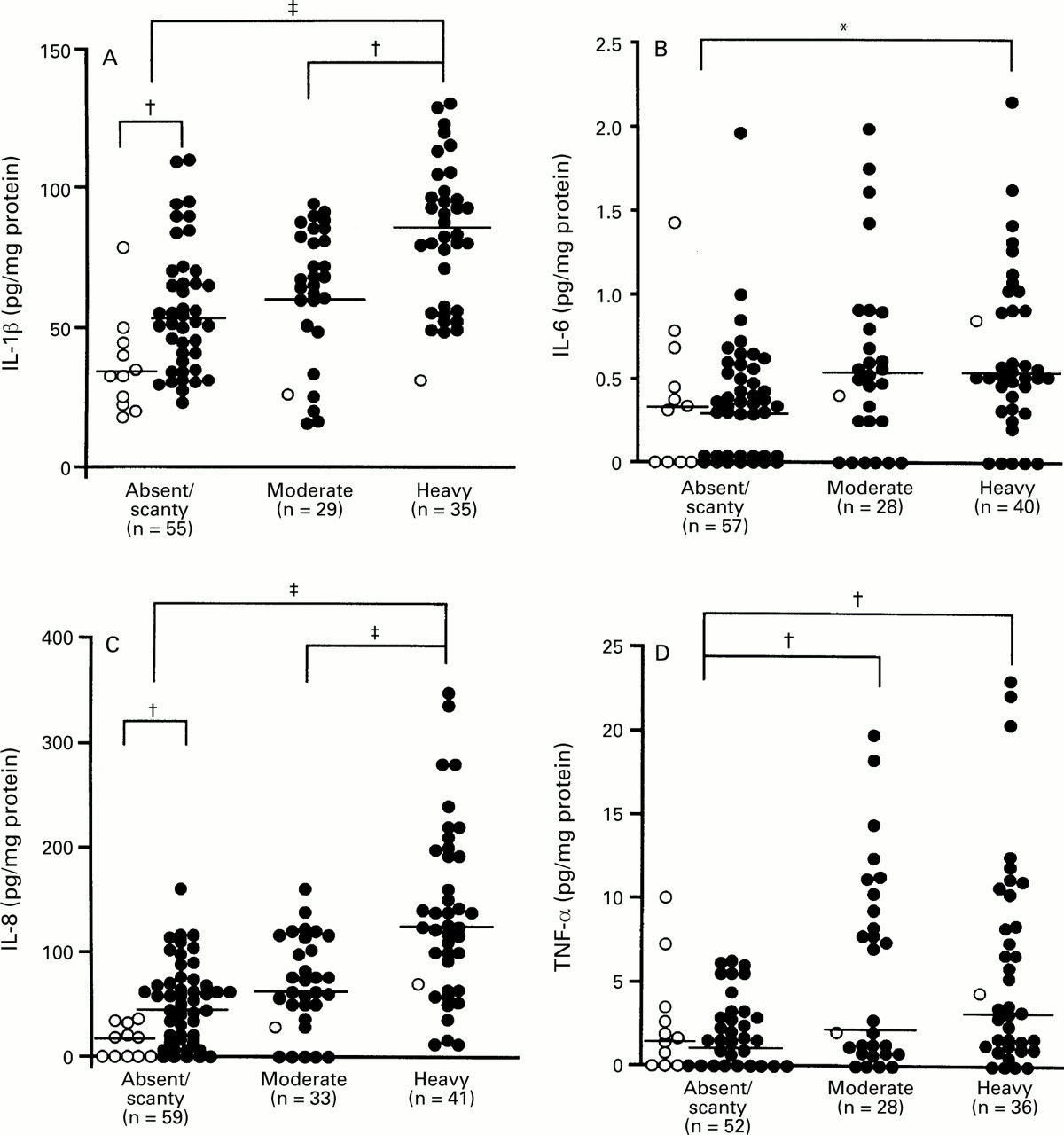
We examined the relation between cytokine production and the density of H pylori in the antral biopsy specimens from patients with H pylori infection (fig 6). Twelve patients were H pylori positive on histological analysis in specimens from the corpus but not in those from the antrum. The mucosal levels of IL-1β, IL-6, IL-8, and TNF-α were significantly higher in specimens with heavy colonisation (IL-1β: median 83.2 pg/mg, range 31.5–130.0; IL-6: median 0.52 pg/mg, range 0.00–2.15; IL-8: median 123.3 pg/mg, range 10.8–346.5; TNF-α: median 3.36 pg/mg, range 0.00–22.98) than in those without or with only scant colonisation (IL-1β: median 50.0 pg/mg, range 17.6–110.0, p<0.0001; IL-6: median 0.34 pg/mg, range 0.00–1.96, p<0.01; IL-8: median 39.4 pg/mg, range 0.0–159.7, p<0.0001; TNF-α: median 1.40 pg/mg, range 0.00–9.96, p<0.005). The mucosal levels of IL-1β and IL-8 were also significantly higher in specimens with heavy than in those with moderate colonisation (IL-1β: median 66.8 pg/mg, range 15.4–92.2, p<0.005; IL-8: median 60.8 pg/mg, range 0.0–159.1, p<0.0001). Furthermore, the mucosal level of TNF-α was significantly higher in specimens with moderate (median 2.13 pg/mg, range 0.00–19.68) than in those without or with only scant colonisation (p<0.005).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
: Production of (A) IL-1β, (B) IL-6, (C) IL-8, and (D) TNF-α and the density of H pylori colonisation. Filled circles, cagA+ specimens; open circles, cagA−specimens. *p<0.05; †p<0.005; ‡p<0.0001 by Mann-Whitney U test.
Eleven of 13 cagA − specimens showed no or scant colonisation by H pylori. Even in such specimens, the mucosal levels of IL-β and IL-8 were significantly higher incagA + than in cagA −specimens (p<0.0001) (fig 6).
Discussion
This study showed that the mucosal levels of IL-1β, IL-6, IL-8, and TNF-α were significantly higher in H pylori positive than in H pylori negative specimens, in agreement with previous studies.2-6 ,16 ,17 In H pyloripositive specimens, the mucosal levels of IL-1β, IL-6, IL-8, and TNF-α were significantly correlated with the infiltration of MNC and PMN, and the mucosal levels of IL-6 and IL-8 were significantly higher in specimens with duodenal ulcer than in those with gastritis without ulcers. These findings suggest that IL-1β, IL-6, IL-8, and TNF-α have some role in gastroduodenal inflammation caused by H pylori infection. However, there was still significant overlap in production of cytokines between ulcer groups and gastritis groups. Therefore, production of cytokines cannot be a single marker of ulceration. Of 44 H pylori negative specimens, there were eight H pylori negative specimens in which at least one of IL-6, IL-8, or TNF-α was detected. Seven of these specimens had MNC infiltration, suggesting that these cytokines play a role in cellular inflammation even in H pylori negative specimens.
In the present study, we also examined the relationship between thecagA gene and the production of various cytokines using quantitative ELISA methods, and found that the mucosal levels of IL-1β and IL-8 were significantly higher incagA + than in cagA −specimens. With regard to IL-8, our results were in agreement with those of Peek et al 18 who reported that the gastric mucosal level of IL-8 was significantly higher in patients infected with cagA + thancagA − strains. In our previous study,6 we also found that IL-8 mRNA expression was significantly more common in cagA + than incagA − specimens. Furthermore, recent in vitro studies indicated that CagA/cagA gene positive strains stimulated greater release of IL-8 than CagA/cagAgene negative strains using gastric epithelial cell lines.10 ,11 However, recent studies have demonstrated that isogenic cagA mutants with disruption of CagA expression also produce IL-8.12 Tummuru et al 13 reported that the picB gene, which is present upstream of the cagA gene, was essential for induction of IL-8 from gastric epithelial cells in vitro. Strains possessing the picB gene, however, have also been reported to express the cagA gene in vivo.13Therefore, the cagA + strains (cagA gene positive and picB gene positive) may have the ability to induce IL-8 from gastric mucosa.
IL-8 possesses potent chemotactic activity for neutrophils19 ,20 and T lymphocytes.21Indeed, we found in the present study thatcagA + specimens showed significantly more severe infiltration of PMN and MNC thancagA − or H pylori negative specimens. Crabtree et al 9 also reported that mucosal immune recognition of CagA protein was strongly associated with epithelial PMN infiltration. PMN enzymatic products such as proteases and elastases are thought to be important mediators of tissue damage.22 A recent study23 indicated that epithelial injury was mediated by the oxi- dant monochloramine, which is produced by the interaction between ammonia and hypochlorous acid (HOCl) and is an important PMN derived microbicidal agent.
Interestingly, in the present study, we found that the mucosal level of IL-1β was also significantly higher in cagA +than in cagA − specimens. There have been few reports about the relationship between thecagA + strains and the expression patterns of cytokines other than IL-8. Peek et al 18 found that levels of expression of IL-1α and IL-1β mRNA were significantly higher in cagA + than incagA − specimens. However, the mucosal level of IL-1α protein was not increased in cagA +compared with cagA − specimens. The number of patients in their study was low and they examined protein concentrations of only two cytokines (IL-8 and IL-1α). In our previous study,6 the levels of expression of IL-1β, IL-6, IL-7, IL-10, and TNF-α mRNA were a little higher incagA + than in cagA −specimens, but the differences were not significant. However, in these studies, which examined cytokine mRNA expression by non-quantitative RT-PCR methods, we could not affirm that cytokines other than IL-8 were related to cagA + strains. We found in the present study that there were definite relations between thecagA + strains and the production of both IL-8 and IL-1β.
Of course, there was still overlap in production of these cytokines between cagA + andcagA − specimens. It has recently been shown that it is possible to isolate cagA + andcagA − strains from a single patient.24 In the present study, multiple colonies on agar were collected together and cultured again in broth and the DNA extracted; therefore, we cannot deny the possibility thatcagA + and cagA −strains coexisted in the same biopsy specimens. It may be that incagA + specimens lacking production of IL-1β and IL-8, cagA + andcagA − strains coexisted.
In the present study, the mucosal levels of IL-1β, IL-6, and IL-8 were significantly higher in H pylori positive specimens with heavy colonisation than in those with scant colonisation, suggesting that the density of H pylori is a pathogenic factor. However, the mucosal levels of IL-1β and IL-8 were significantly higher in cagA + than incagA − specimens even in those without or only scant colonisation by H pylori, although most of thecagA − specimens showed no or scant colonisation by H pylori. These findings suggest that not only the density but also the strain of H pylori is related to the production of IL-1β and IL-8. To our knowledge, this is the first study to have examined the relation between the production of cytokines and the cagA gene, with due regard to the density of H pylori.
IL-1β concentration was significantly correlated with IL-8 concentration in cagA + specimens. As it is well known that IL-1β induces the production of IL-8,19the significant increase in IL-8 in cagA +specimens might be a consequence of IL-1β activity. IL-1β concentration, however, was high even in H pylori negative specimens, although IL-8 was not detected in most H pylorinegative specimens. This result was in agreement with our previous observation that IL-1β mRNA was expressed even in H pylori negative normal mucosa.5 ,6 As there are various stimuli, including dietary components, in the stomach regardless of the presence of H pylori, the production of IL-1β may, to some degree, represent physiological production.
In the present study, all patients with duodenal ulcer were infected with cagA + strains and the mucosal level of IL-8 was significantly higher in specimens from patients with duodenal ulcer than from those with gastritis without ulcers. These results strengthen the hypothesis that the cagA +strains are more pathogenic. However, more than 90% of the patients were infected with cagA + strains, most of whom had gastritis without developing peptic ulcer. Therefore,cagA + strains may not be equivalent to ulcerogenic strains.
The mucosal levels of IL-6 and TNF-α were increased in bothcagA + and cagA −specimens compared with H pylori negative specimens. In gastric mucosa infected with cagA − strains, therefore, IL-6 and TNF-α may play a role in gastric inflammation. There is no doubt that factors other than CagA/cagA gene also induce the production of IL-6 and TNF-α in H pyloriinfection. Furthermore, neither IL-6 nor TNF-α concentrations were correlated with IL-8 concentration. These findings indicate that IL-6 and TNF-α were induced, at least in part, independently of the production of IL-8. Huang et al 25 also reported that IL-8 stimulation by H pylori was independent of IL-6 production using gastric epithelial cell lines. In the present study, IL-6 levels were very low. Previous studies2 ,3have also indicated that the gastric mucosal concentration of IL-8 is about 700 fold higher than that of IL-6. These results were in agreement with our present observation that the concentration of IL-8 was about 120-fold higher than that of IL-6. However, Gionchettiet al 26 reported that the median concentration of IL-6 in H pylori positive specimens was 120 pg/mg protein using the same methods as used in this study. The only difference between our present and their previous study was that they homogenised the specimens with an ultra turrax homogeniser, which was more powerful than that used here. This suggested that the concentration of IL-6 is dependent on homogenisation of specimens. Further studies are required to determine the role of IL-6 in H pylori associated gastritis.
The mucosal level of TNF-α was much lower than that of IL-1β in the present study, although the abilities of IL-1β and TNF-α to induce IL-8 are similar in vitro.20 This may explain the lack of correlation between IL-8 and TNF-α levels, although both IL-1β and TNF-α are known to stimulate IL-8 production. In the present study, the TNF-α concentration was significantly correlated with that of IL-6, in agreement with previous findings.2 Interestingly, in cagA − specimens, even at high concentrations of IL-6 and TNF-α, PMN and MNC infiltration was not severe compared with that in cagA + specimens; IL-6 and TNF-α may therefore not play major roles in gastric inflammation.
With regard to the production patterns of IL-7 and IL-10, the present results were not in agreement with those of our previous studies,5 ,6 in which the levels of mRNA expression of these cytokines were shown to be increased in H pyloripositive relative to H pylori negative specimens using RT-PCR. The discrepancies between protein and mRNA levels may have been due to blockage of the pathway from mRNA to protein expression, degration of mRNA prior to translation into protein, or simply the fact that the amount of mRNA did not exceed the threshold required for protein production.
Furthermore, there were interesting findings in the production of IL-10 and IL-7. In the present study, two specimens with mucosal levels of IL-8 greater than 300.0 pg/mg also showed high levels of IL-10. On the other hand, the mucosal level of IL-10 was higher in H pylori positive specimens with low levels of IL-8 than inH pylori positive specimens overall. This may explained as follows. IL-10 is thought to have a strong down regulatory effect on proinflammatory cytokines such as IL-1β, IL-6, IL-8, and TNF-α.27 When the mucosal levels of these proinflammatory cytokines exceed a certain threshold, the production of IL-10 protein is also increased and the production of these proinflammatory cytokines may be reduced via the down regulatory effects of IL-10 over the course of time. Due to this complicated cytokine network, it may be very difficult to analyse the production of IL-10 by one point examination of biopsy specimens.
IL-7 is thought to be a potent inducer of IL-6, IL-1α, IL-1β, and TNF-α expression, suggesting that IL-7 is an important mediator in inflammation.28 However, there were no relations between the production of IL-7 and cellular inflammation, disease state, or density of H pylori. In the previous study,6IL-7 mRNA was also expressed in a number of specimens from H pylori negative patients, and the expression rate of IL-7 mRNA was not significantly decreased after eradication of H pylori. Therefore, IL-7 may not play an important role in gastric inflammation. However, Ueno et al 29recently demonstrated that IL-7 was expressed only after differention into goblet cells using a human colonic epithelial cell line. Although there was no significant relation between metaplasia and the production of IL-7 in the present study, the specimen with the highest level of IL-7 had severe metaplastic mucosa in the antrum. Further studies are required to determine the role of IL-7 in H pylori associated gastritis.
In conclusion, our findings suggest that the ability to induce cytokines differs among strains of H pylori;cagA + strains induce various kinds of cytokines and may cause severe inflammation, whilecagA − strains have weak ability to induce IL-1β and IL-8 and may cause only mild inflammation. However, most patients infected with the cagA + strains have gastritis, and do not develop peptic ulcers. Therefore, although thecagA + strains are pathogenic, they do not seem to be ulcerogenic.
References
Linked Articles
- COMMENTARY