Article Text

Abstract
Background—It is well established that glutamine supplemented elemental diets result in less severe intestinal damage in experimental colitis. However, few studies have examined the mode of action of glutamine in reducing intestinal damage.
Aims—To examine the effects of glutamine supplemented elemental diets on the potent inflammatory cytokines interleukin 8 (IL-8) and tumour necrosis factor α (TNF-α) in trinitrobenzene sulphonic acid (TNBS) induced colitis which presents with both acute and chronic features of ulcerative colitis.
Methods—Sprague-Dawley rats were randomised into three dietary groups and fed 20% casein (controls), or 20% casein supplemented with either 2% glutamine (2% Gln) or 4% glutamine (4% Gln). After two weeks they received intracolonic TNBS to induce colitis.
Results—Both Gln groups of rats gained more weight than the control group (p<0.05) which had progressive weight loss. Colon weight, macroscopic, and microscopic damage scores for the Gln groups were lower than in the control group (p<0.05). IL-8 and TNF-α concentrations in inflamed colonic tissues were lower in the Gln groups than in the control group (p<0.05), and correlated well with disease severity. Bacterial translocation was lower both in incidence (p<0.05) and in the number of colony forming units (p<0.05) for the Gln groups, than in the control group. With respect to all indices studied, the 4% Gln group performed better than did the 2% Gln group.
Conclusion—Prophylactic glutamine supplementation modulates the inflammatory activities of IL-8 and TNF-α in TNBS induced colitis.
- glutamine
- trinitrobenzene sulphonic acid
- inflammatory bowel disease
- rats
- interleukin 8
- tumour necrosis factor α
Statistics from Altmetric.com
- glutamine
- trinitrobenzene sulphonic acid
- inflammatory bowel disease
- rats
- interleukin 8
- tumour necrosis factor α
The aetiology of inflammatory bowel disease (IBD), encompassing Crohn’s disease (CD) and ulcerative colitis (UC), remains unknown. Although the exact pathogenesis is poorly understood, there is evidence that it involves interactions between the immune system, genetic susceptibility, and the environment, most notably the bacterial flora.1 Other evidence indicates infectious, psychogeneic, and allergic aetiologies as well as involvement of the vascular system.2 Food is also a major factor affecting the intestinal environment, and diet has therefore been implicated in the pathogenesis of these diseases.3 ,4 However, the mechanisms underlying initiation, progression, and chronicity may well be distinct for each implicated factor.1
The role of nutrition in the management of IBD has received increasing attention. Consequently various dietary measures have been recommended as adjunctive, if not primary therapies.3-6 Recent studies have indicated that glutamine, the principal respiratory substrate for enterocytes,7 is important for intestinal metabolism especially following stress.8 Glutamine fortified parenteral and enteral diets significantly improve intestinal morphology and function.9-12 Animal models of colitis have indicated that glutamine supplemented elemental diets resulted in less severe intestinal damage, less weight loss, and improved nitrogen balance,13 and reduced intestinal bacterial translocation.14 ,15
Many inflammatory mediators have been implicated in IBD, the most potent of which appear to be interleukin 8 (IL-8) and tumour necrosis factor α (TNF-α). IL-8 differs from the others in that it is mainly produced by monocytes and macrophages,16-18 is highly selective for neutrophils,16-20 and is known to persist in its active form for long periods in tissues; its action is thus likely to be protracted.19 ,21 Greater quantities of IL-8 have been found in active UC than in inactive disease or in active CD.22 ,23 We therefore investigated the effect of glutamine supplementation on IL-8 and TNF-α levels and on disease outcome.
Although experimental animal models of colitis are of value in eliciting some of the mechanisms of IBD, the different models differ in their relative use.1 Most animal models reported lack certain characteristics of IBD and model acute events rather than the typical chronic features of IBD. We therefore utilised trinitrobenzene sulphonic acid (TNBS) induced colitis, which has the dual advantage of chronicity and acuteness1 so often seen in patients with IBD. Other advantages of this model of colitis are its reasonable reproducibility,1 and its value in assessing the efficacy of therapeutic agents commonly used in the treatment of colitis.24 ,25
Methods
ANIMALS
Specific pathogen free Sprague-Dawley rats were obtained from Kyudo Breeding Laboratories (Kumamoto, Japan) and used for the experiments. Rats were kept in a room of constant temperature (25±2°C) and humidity (50–70%) with a 12 hour light period. Animal care was in compliance with applicable guidelines from the Ryukyus University Policy on Animal Care and Use. The rats were allowed to adapt to our laboratory environment for one week before the onset of the experiment, during which period they were maintained on a commercial non-purified diet (Nihon clear, Osaka, Japan). This standard diet contains 25.5% protein and 4.3% fat by weight.
STUDY PROTOCOL
After the period of adaptation, weight matched female rats (200–250 g) were randomised into three groups according to the dietary treatment (15 animals in each group). Each diet was started two weeks before the instillation of TNBS to stabilise the rats, and was continued until sacrifice. After two weeks on the diet each rat received an intracolonic instillation of TNBS (50 mg/0.5 ml). Two weeks later rats were killed and damage to the colon was assessed.
TRINITROBENZENE SULPHONIC ACID
2,4,6-Trinitrobenzene sulphonic acid (Wako Chemical Co., Tokyo, Japan) was dissolved in 50% ethanol to a final concentration of 100 mg/ml, and 0.5 ml was intracolonically instilled into each animal used for the experiment.
DIET
Rats were fed either a 20% casein diet (controls), or 20% casein supplemented with either 2% (w/w) glutamine (2% Gln) or 4% (w/w) glutamine (4% Gln). Glutamine was obtained from Ajinomoto Company, Tokyo, Japan. All diets were fibre free (table 1). The diets were made isonitrogenous and isocaloric by adding appropriate amounts of glycine. Animals in all groups were fed ad libitum and allowed free access to water. Between 0900 and 1000 every morning, animals were weighed and food and water were renewed.
Composition of diets
INDUCTION OF EXPERIMENTAL COLITIS
After a 24 hour fast, rats in each dietary group were sedated by intraperitoneal administration of 50 mg/ml sodium pentobarbital (Wako Chemical Co., Japan; 0.1 ml/100 g body weight). A polypropylene catheter was lubricated with jelly and inserted 8 cm, via the anal canal into the colon of the rat just proximal to the splenic flexure. TNBS (100 mg/ml) dissolved in 50% (vol/vol) ethanol was instilled into the colon (total volume of 0.5 ml per rat). After instillation the rats were supported in the supine position until recovery from anaesthesia to prevent immediate anal leakage of the instillate.
TESTING FOR TRANSLOCATING BACTERIA
Two weeks after the instillation, rats were killed by intraperitoneal administration of sodium pentobarbital. Their abdomens were shaved and cleaned with isodine and opened with sterile instruments. The spleen and mesenteric lymph nodes (MLNs) were aseptically removed and placed in preweighed glass tubes containing sterile brain heart infusion (BHI) broth (Wako Chemical Co., Tokyo, Japan) which were then reweighed. Spleen and lymph nodes were homogenised in sterile BHI in an ice cooled water bath using a previously sterilised homogeniser piston (Ystral, Mitamura Riken Kogyo, Japan). Tenfold serial dilutions were prepared in BHI and 100 μl aliquots of the homogenates were cultured in polymyxin B nalidixic acid agar and McConkey’s agar for detection of Gram positive and Gram negative organisms. Blood drawn from the heart (100 μl) was mixed in 0.9 ml BHI and similarly cultured as above. All plates were incubated at 37°C. After two days colonies were counted and microorganisms were quantified as colony forming units per gram tissue (CFU/g) or per ml blood (CFU/ml) using the fomula:  where N is the number of colonies on the plate, D the dilution inoculated on the plate, C the volume of BHI used for homogenising tissue or mixing blood, and W or Vthe weight of specimen in grams or the volume of blood in ml, respectively; 10 is included for the 0.1 ml inoculum. Representative colonies on both agar plates were identified by standard bacteriological procedures.
where N is the number of colonies on the plate, D the dilution inoculated on the plate, C the volume of BHI used for homogenising tissue or mixing blood, and W or Vthe weight of specimen in grams or the volume of blood in ml, respectively; 10 is included for the 0.1 ml inoculum. Representative colonies on both agar plates were identified by standard bacteriological procedures.
ASSESSMENT OF COLONIC DAMAGE
The distal colon was removed, opened longitudinally, and cleared of faecal material with a gentle spray of 0.9% saline. The freshly opened colonic segments were pinned out on a wax block and examined by two independent observers blinded to the treatment (TM, AA). The extent of mucosal damage was assessed using the colon macroscopic scoring system of Wallace et al.24 After scoring, the detached colon was blotted dry and weighed. The colon weight:body weight ratio was calculated. This ratio has been shown to be a marker of colonic inflammation.26
COLON HISTOLOGY
Samples of the inflamed tissues were removed for histological analysis. The tissues were fixed in 37–40% phosphate buffered formalin (37–40% formaldehyde, 10 ml; sodium phosphate monobasic, 1.86 g; sodium hydroxide, 0.42 g; distilled water, 90 ml) at room temperature overnight. The tissues were then sliced into 4–6 mm pieces, dehydrated in ethanol, embedded in paraffin wax, sectioned, and stained with haematoxylin and eosin. The microscopic slides were reviewed by two histologists (TM and AA) blinded to the experimental groups and the extent of damage; colonic inflammation was assessed using a modification of the histopathological grading system of Macpherson and Pfeiffer27 (table 2). The sections were examined and photographed with an Olympus microscope (Olympus Kogyo Co., Tokyo, Japan).
Histopathological grading scale of chemically induced colitits
COLON CYTOKINE EXTRACTION
Samples of inflamed and non-inflamed colon were weighed and homogenised for one minute in 0.01 M phosphate buffered saline (PBS), pH 7.4, in an ice cooled water bath. The homogenates were then centrifuged at 10 000 g at 4°C for 30 minutes and filter sterilised before the assay. Serum was prepared from cardiac blood and stored frozen at –80°C until analysis. IL-8 and TNF-α were quantified with a commercial rat IL-8 (Panapharm Laboratories Co. Ltd, Kumamoto, Japan) or TNF-α (Genzyme Co., Cambridge, Massachusetts, USA) ELISA kit according to the manufacturer’s instructions. The interassay and intraassay coefficients of variation were less than 10%. The sensitivities of the assays were less than 5 ng/ml and 8 ng/ml for IL-8 and TNF-α respectively.
STATISTICAL ANALYSIS
Statistical analysis of the data was performed by analysis of variance (ANOVA); the Mann-Whitney U test was used for damage scores and Spearman rank correlation was used to correlate tissue cytokine levels with damage scores. Duncan’s multiple range test was used to determine significant differences among means. A value of p<0.05 was considered significant.
Results
GENERAL FINDINGS
One day post-induction, rats in all dietary groups had watery diarrhoea and reduced activity. Towards sacrifice, rats in the control group had difficulty in passing faeces, were increasingly lethargic, and showed piloerection. Rats in the Gln groups occasionally passed soft faeces but exhibited little to no lethargy. Prior to the induction of colitis, weight gain was consistent and similar among all dietary groups. Two weeks after induction, after a slight weight drop, both Gln supplemented dietary groups had an increase in weight to a mean value higher than (but not significantly different from) their preinduction weights while the control group of rats continued to lose weight throughout (data not shown). Both Gln groups of rats had a weight gain that was higher than the control group (p<0.05) (table 3). The 4% Gln group had a weight gain that was higher than that of the 2% Gln group (p<0.05). There were no deaths in any of the groups during the period of investigation.
Body weight gain and colonic inflammation in the different dietary groups
BACTERIAL TRANSLOCATION
Both Gln groups of rats showed a lower incidence of bacterial translocation (p<0.05) and also a lower number of colony forming units compared with the control group (p<0.05) (table 4). The incidence of translocation and the number of colony forming units were lower in the 4% Gln group than in the 2% Gln group. Translocation occurred into at least two of the tissues studied in each dietary group. Translocating bacteria were identified as Escherichia coli, Pseudomonas aeruginosa, Mycobacterium morgani, and Enterococcusspecies.
Incidence of bacterial translocation in the different dietary groups
COLONIC INFLAMMATION
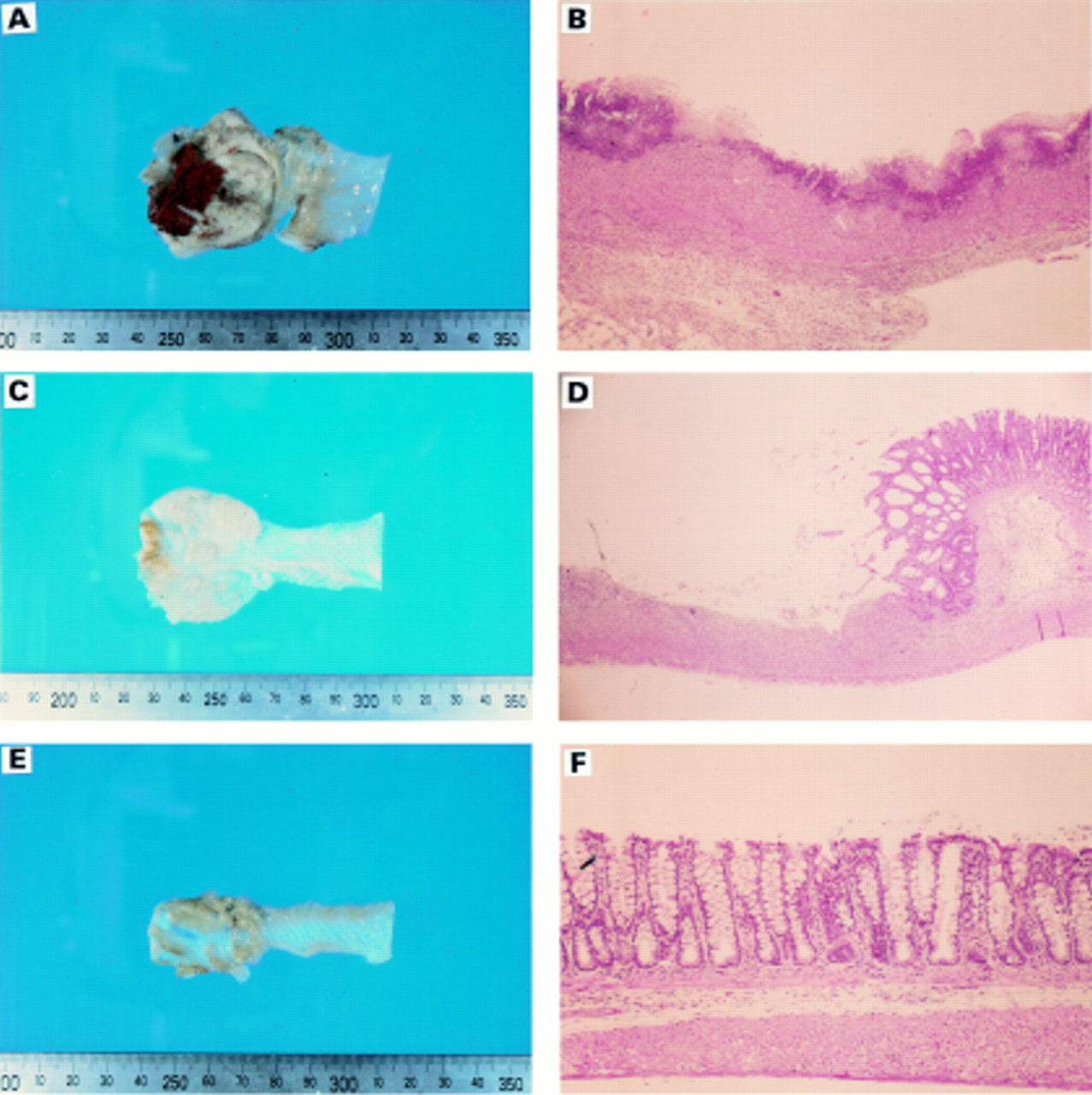
Two weeks post-induction, the control group of rats had severe faecal stasis resulting in partial bowel obstruction; a few rats on both glutamine supplemented diets had smaller and varying degrees of bowel obstruction with the 4% Gln group showing the least obstruction. In the control group, the damage comprised broad mucosal ulcers with a surface layer of necrotic slough, accumulation of mesenteric fat, and fibrinous adhesions to the bowel. Acute colonic damage with haemorrhage and bowel wall thickening were also observed (fig 1). Colon weight and macroscopic damage scores in the 4% Gln group were lower compared with the other groups (p<0.05) (table 3, fig 2A) The 2% Gln group also had a lower score compared with the control group (p<0.05) (fig 2B). Microscopic examination showed infiltration of neutrophils, lymphocytes, macrophages, fibroblasts, and connective tissue mast cells, with extensive crypt distortion and transmural necrosis (fig2A), which was less severe in the 4% Gln group than in 2% Gln group, both of which were lower than the control group (p<0.05) (fig2B).

: Sample sections of rat colonic mucosa from each dietary group. Control (A,B); 2% Gln (C,D); 4% Gln (E,F). The varying degrees of necrosis of the intestinal mucosa and infiltration by inflammatory cells among the groups are evident.

: Colon macroscopic (A) and microscopic (B) scores of rats. Values represent mean (SEM). Bars with different letters indicate significant difference (p<0.05).
IL-8 AND TNF-α CONCENTRATION IN INFLAMED AND NON-INFLAMED COLONIC PORTIONS
Compared with the other groups, the 4% Gln group had the lowest concentration of IL-8 and TNF-α (p<0.05) (table 5), in both inflamed and non-inflamed colonic tissues which correlated well with the damage score (IL-8: r=0.894, p<0.05; TNF-α:r=0.744, p<0.05, inflamed colon). The 2% Gln group also had a lower TNF-α concentration (r=0.832, p<0.05, inflamed colon) in both portions of the colon compared with the control group (r=0.893, p<0.05), which also correlated well with the damage score. The IL-8 concentration was also lower in the inflamed portions of the colon in the 2% Gln group (r=0.874, p<0.05) compared with the control group (r=0.965, p<0.05) but concentrations in the non-inflamed portions were similar in the two groups. Generally, higher levels of both cytokines were observed in inflamed tissues than in non-inflamed tissues (p<0.05).
Interleukin 8 (IL-8) and TNF-α concentrations in inflamed and non-inflamed rat colon
SERUM IL-8 AND TNF-α CONCENTRATIONS
The 4% Gln group had the lowest concentration of serum IL-8, compared with the other groups (p<0.05). The 2% Gln group also had a lower concentration of serum IL-8 than did the control group (p<0.05) (fig 3). Interestingly, however, serum TNF-α concentrations in all three dietary groups were negligible.

{kind=link}
{kind=link}
{kind=link}
: Serum IL-8 concentrations in rats. Values represent mean (SEM). Bars with different letters indicate significant difference (p<0.05).
Discussion
Our results confirm other reports that glutamine supplementation of elemental diets may improve the condition of patients with IBD, and specifically that 4% Gln may offer a better response than 2% Gln. A previous study utilising another model of colitis with 2% Gln supplementation reported reduced endotoxaemia, but could not show mucosal regeneration as determined by colon ornithine decarboxylase levels, which might precede the restoration of gut mucosa after injury.14 Although we did not measure colon regeneration by enzymatic studies, both our macroscopic and microscopic examination revealed smaller and more superficial necrotic areas and a less intense accumulation of inflammatory cells in the 4% Gln and the 2% Gln group compared with the more deep seated damage and intense accumulation of inflammatory cells in the control group. In our study we stabilised the rats on the diets for two weeks before inducing colonic damage, whereas in the previous study, glutamine supplementation was started in the post-induction state. This, together with the length of time of supplementation and the model of colitis used, may be responsible for the observed differences.
Fox et al,13 also utilising the dietary prestabilisation mode of feeding in a lethal model of colitis, showed that glutamine supplemented enteral diets significantly improved nutritional status, decreased intestinal injury, decreased bacterial translocation, and resulted in improved survival which is in agreement with our results. In a related study using endotoxin injection, Barberet al,15 working with a defined fomula diet supplemented with either 2% Gln or fibre, showed that while both glutamine and fibre maintained small bowel mass, only glutamine preserved normal jejunal mucosal architecture. Furthermore, even though neither fibre nor glutamine supplementation prevented caecal bacterial overgrowth or bacterial translocation, glutamine was associated with a significantly lower bacteraemia; this also supports our results. In yet another study, similar to the present one, Apteker et al 28 found both macroscopic and microcopic damage scores and also myeloperoxidase activity to be lower in rats pretreated with glutamine enemas before induction of colitis compared with rats treated after the induction of colitis. Klimberg et al 29 also reported that prophylactic glutamine protects the intestinal mucosa from radiation injury, and concluded that glutamine exerts a protective effect on the mucosa by supporting crypt cell proliferation which may accelerate healing of the acutely radiated bowel. Together, these results suggest that pretreatment with glutamine acts as protection against intestinal damage; our results indicate 4% Gln to be more effective than 2% Gln.
In TNBS induced colitis, there is severe neutrophil infiltration,24 ,30-35 possibly due to local release of neutrophil chemotactic agents. Many lipid derived chemoattractants such as leukotriene B4,36 platelet activating factor (PAF),37 a product of the complementary pathway C5a,38 and also IL-8,16-18 have been implicated in neutrophil infiltration in inflammatory bowel disease. However, the lipid derived chemotactic agents are short lived19 and would not be sufficient explanation for neutrophil emigration.34 ,39 IL-8 differs from the others in that it is mainly produced by monocytes and macrophages,16-18 is highly selective for neutrophils,16-20 and is known to persist in tissue in its active form for long periods; its action is thus likely to be protracted.19 ,21 Furthermore, some studies have found greater quantities of IL-8 in active UC than in inactive disease or in active CD.22 ,23 We therefore considered the effect of glutamine supplementation on IL-8 and TNF-α, which is also a chemoattractant, on disease outcome. As we had previously observed that 4% Gln resulted in a higher survival than 2% Gln in methicillin resistant Staphylococcus aureus infection,4we compared the effects of a two concentrations of glutamine supplementation. The present study showed that while both Gln groups had less damage and bacterial translocation, and also lower concentrations of the potent inflammatory cytokines IL-8 and TNF-α compared with the control group, the 4% Gln group performed better than the 2% Gln group. With the low concentration of both cytokines observed in the Gln groups, it is possible that Gln may have inhibited synthesis, release, and/or action of these inflammatory cytokines, resulting in the improvement in disease outcome. We have no ready explanation for the neglegible serum TNF-α concentration observed. However, it is known that serum levels of this cytokine are not consistently elevated in accordance with disease severity.41
How glutamine reduced bacterial translocation in the prescence of colonic damage is an interesting question. Burke et al 42 found that glutamine supplemented total parenteral nutrition maintained secretory IgA (S-IgA) concentration at normal levels and suggested that glutamine may be important in the gut associated lymphoid tissue (GALT). S-IgA functions primarily by preventing the attachment of bacteria to the mucosal cell.43 It is conceivable that the higher glutamine dose may have further enhanced the efficiency of the GALT in eliminating intestinal bacteria. Some studies have reported a consistent obligate requirement for the presence of normal intestinal flora in the genesis of intestinal inflammation.44-46
Glutamine is a key substrate for rapidly dividing cells such as enterocytes.47 Gut glutamine requirements are increased in catabolic disease states.48 A high rate of glutamine uptake but only partial oxidation is characteristic of rapidly dividing cells such as enterocytes, immunocytes, macrophages, and fibroblasts. This situation provides ideal conditions for the synthesis of key molecules such as glutathione and nucleotides.49 We recently found that dietary supplementation with a nucleoside-nucleotide mixture which is also important for the maintenance of gut integrity, but has proinflammatory properties, worsened colonic mucosal damage in experimental colitis.50Since glutamine is a substrate for nucleotide synthesis, it could be expected that increased supplements may enhance nucleotide synthesis which in turn may aggravate disease outcome. From our results, however, it is obvious that this was not the case, which indicates the preferential use of glutamine for the maintenance of mucosal integrity during damage. Further clarification of the mechanism of action of glutamine supplementation on intestinal damage is necessary.
This model of experimental colitis has been well characterised and has clinical, biochemical, and pathological similarities to colonic CD.29 Animals with TNBS colitis have responded to drugs useful in IBD.24 ,25 This observation, together with the availability of a quantitative scoring system, makes it a useful system for the evaluation of new therapeutic agents.51 Many studies have utilised this model of colitis to assess the therapeutic efficacy of glutamine; to our knowledge, however, our study is the first to assess the effect of glutamine supplementation on the potent inflammatory cytokines IL-8 and TNF-α in TNBS induced colitis. Although it is not possible to extrapolate findings from animal models to the human situation, these results suggest that an increased level of glutamine supplementation may offer a better response in the treatment of IBD.
Acknowledgments
The authors would like to thank Ajinomoto Co., Tokyo, Japan, for the supply of glutamine.