Article Text

Abstract
Background—Two different types of secretory phospholipase A2 (PLA2), pancreatic group I (PLA2-I) and non-pancreatic group II (PLA2-II), have been identified and postulated to be associated with the pathogenesis of various diseases, such as acute pancreatitis, septic shock, and multiple organ failure.
Aims—To investigate the type of secretory PLA2 responsible for its catalytic activity found in plasma and ascites of experimental acute pancreatitis.
Methods—Acute pancreatitis of differing severity was induced by the injection of different concentrations (1% or 10%) of sodium deoxycholate (DCA) into the common biliopancreatic duct in rats, and catalytic PLA2 activity in plasma and ascites were differentiated by anti-PLA2-I antibody and specific inhibitor of PLA2-II. Survival rate and plasma amylase, aspartate aminotransferase (AST), and alanine aminotransferase (ALT) were also measured.
Results—In 1% and 10% DCA induced acute pancreatitis, plasma amylase values as well as PLA2activity in ascites were greatly increased. PLA2 activity in plasma was also notably increased in 10% DCA induced acute pancreatitis, but not in 1% DCA induced acute pancreatitis. PLA2-I specific polyclonal antibody significantly inhibited PLA2 activity in ascites but not that in plasma. In contrast, plasma PLA2 activity was completely suppressed by PLA2-II specific inhibitor. In addition, a high mortality (93% at five hours) and a significant increase in plasma AST and ALT were noted in 10% DCA induced pancreatitis.
Conclusion—Ascites PLA2 activity is mainly derived from PLA2-I, whereas plasma PLA2activity is mostly derived from PLA2-II in severe acute pancreatitis, suggesting that increased plasma PLA2-II activity might be implicated in hepatic failure arising after severe acute pancreatitis.
- acute pancreatitis
- phospholipase A2
- sodium deoxycholate pancreatitis
- hepatic failure
Statistics from Altmetric.com
Phospholipase A2 (PLA2) consists of a diverse family of enzymes that hydrolyse the sn-2 fatty acyl ester bond of phosphoglycerides liberating free fatty acid, predominantly in the form of arachidonic acid, and lysophospholipids. Once formed, the two products, free arachidonic acid and lysophospholipid, are widely implicated in the production of potent inflammatory mediators such as prostaglandins, leukotrienes, and platelet activating factor.1 PLA2 exists in both extracellular and intracellular forms.2 The most widely studied extracellular PLA2, the secretory PLA2, consists of 14 kDa calcium dependent enzymes. Secretory PLA2 can be classified into two types, group I (PLA2-I) and group II (PLA2-II), based on their primary structures.3 Mammalian PLA2-II is found in inflammatory regions, such as casein induced peritoneal fluid in rats,4 carrageenan induced pleural exudate in rats,5 and synovial fluid of patients with rheumatoid arthritis.6 In addition, some inflammatory cytokines and lipopolysaccharides dramatically increase PLA2-II secretion in several rat tissues through enhancement of gene transcription.7 ,8 These findings strongly implicate mammalian PLA2-II as one of the factors which promote inflammatory processes.9 Some studies have in fact shown the proinflammatory activities of PLA2-II.10-12
The other type, mammalian PLA2-I, occurs abundantly in the pancreas and has long been thought to act as a digestive enzyme. PLA2-I is synthesised and secreted by pancreatic acinar cells as an enzymatically inactive pro-PLA2-I, which is normally activated by trypsin in the lumen of the duodenum.13 Furthermore, it has been postulated that PLA2-I is associated with the pathology of acute pancreatitis,14 in which PLA2 activity in serum is increased, especially in the severe necrotic form of the disease.15 Experimental studies on acute pancreatitis have shown signs of hydrolysis of phospholipids in necrotic pancreatitis.16
Pancreatitis induced by bile acids, such as sodium taurocholate in the rat, is a frequently used experimental model for evaluating therapeutic regimens in this disease,17 and all the signs of acute pancreatitis are similar to those seen in humans: oedema and pancreatic necrosis, increased serum amylase activity, and decreased pancreatic enzyme content.18 Although we have observed an increase in PLA2 activity in serum and ascites in sodium deoxycholate (DCA) induced pancreatitis,19 the type of PLA2responsible for its catalytic activity is still unknown.
In the current study we induced different severities of acute pancreatitis by injecting rats with different concentrations of DCA, and investigated the type of secretory PLA2 responsible for its activity in plasma and ascites by using a PLA2-I specific polyclonal antibody and a PLA2-II specific inhibitor. Furthermore we attempted to correlate the increased PLA2-II activity with hepatic failure and mortality following the acute pancreatitis.
Materials and Methods
MATERIALS
[9,10-3H]Oleic acid (5 Ci/mmol) was purchased from Amersham (Little Chalfont, UK). Sodium deoxycholate (DCA) was purchased from Difco (Detroit, Michigan, USA). Thielocin A1β was prepared as previously reported.20 21 Rat PLA2-I was isolated from rat pancreas homogenate according to the method of Onoet al.22 Rat PLA2-II was purified from rat platelets as described previously.23 Both purified PLA2 types showed a single band of approximately 14 kDa by Coomassie brilliant blue staining on sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE). Rabbit antirat PLA2-I antibody was prepared at Shionogi Research Laboratories. All other reagents were of analytical grade or better.
DCA INDUCED PANCREATITIS IN THE RAT
These animal experiments were approved by the Animal Experimentation Committee of Shionogi Research Laboratories, Osaka, Japan. Acute experimental pancreatitis was induced in male eight to nine week old Wistar rats (Shizuoka Laboratory Center, Shizuoka, Japan) with a body weight of 200–250 g.24 After fasting overnight, the animals were anaesthetised with an intraperitoneal injection of pentobarbital (50 mg/kg) and were laparotomised. The duodenal loop was pulled out, and a polypropylene catheter (internal diameter 0.5 mm, external diameter 0.8 mm) was introduced through the intact wall of the duodenum via the papilla of Vater into the common biliopancreatic duct. After obstruction of the common bile duct at the porta hepatis with a clamp, pancreatitis was induced by retrograde infusion of 0.3 ml of 1% or 10% DCA into the common biliopancreatic duct over 10 seconds. The clamp was removed 10 seconds after the infusion, the catheter was withdrawn, and the duodenotomy was closed with surgical adhesive (Aron Alpha A, Sankyo, Japan). The abdomen was then closed with Michel’s clips. The 0% DCA pancreatitis group received an identical amount of saline infusion. Animals in the pancreatitis sham group received only cannulation of the biliopancreatic duct without infusion. At given intervals, the animals were anaesthetised subcutaneously with 50 mg/kg pentobarbital. After performing a relaparotomy, ascites samples were collected with a disposable tube containing EDTA Na (7.7 mM, pH 7.4) and prostaglandin E1 (PGE1) (1.5 μg/ml). Thereafter, plasma samples were obtained from the abdominal aorta using a 21 gauge needle and a trigonal stopcock. Plasma samples (1 ml) for the PLA2 assay were collected with a syringe containing 0.1 ml of EDTA Na (77 mM, pH 7.4) and PGE1 (15 μg/ml). Plasma samples (2.5 ml) for the determination of amylase, aspartate aminotransferase (AST), and alanine aminotransferase (ALT) were then collected via a heparinised syringe. Plasma samples were centrifuged at 2000 g for 15 minutes at 4°C immediately after collection to remove the cells, and the supernatants were stored at –20°C until analysis. Amylase, ALT, and AST activities were determined with a Technicon SMAC III (C9100, Technicon Instruments, Tarrytown, New York, USA).25
PLA2 ASSAY
PLA2 activity was measured using radiolabelledEscherichia coli membranes. This assay is commonly used to measure the activities of secretory PLA2.26 ,27 [3H]Oleic acid labelled E coli membranes were prepared by standard procedures.28 For assaying PLA2 activity, samples were incubated for 20 minutes at 37°C with E coli substrate (10 000 cpm containing approximately 0.05 nmol of phosphatidylethanolamine and phosphatidylglycerol) in a total volume of 250 μl of Tris HCl buffer (100 mM, pH 7.4), containing 3 mM Ca2+. The reaction was stopped by adding 1.25 ml of Dole reagent (2-propanol:heptane:1N H2SO4, 78:20:2, vol/vol/vol) and vortexing. After the addition of distilled water (0.5 ml) and heptane (0.75 ml), the contents were mixed on a Directmix TS-100 (Thermal Kagaku Sangyou, Tokyo, Japan) for 10 minutes and centrifuged at 1500 g for 10 minutes. An aliquot (0.8 ml) of the upper phase was pipetted into another tube containing 1 ml of heptane; 100 mg of silicic acid powder (Silica gel 60, 70–230 mesh ASTM, Merck) was added, and the tubes were shaken on a Directmix for six minutes. After centrifugation, the heptane phase was transferred into scintillation vials. Pico-fluor 40 (Packard) 6 ml was added and radioactivity was counted in a Packard liquid scintillation counter. The reaction was started by addition of the samples or purified enzymes. Thielocin A1β was added to assay tubes as DMSO solutions (2% of final volume). Control experiments showed that DMSO at this concentration had no effect on PLA2 activity. Anti-PLA2-I antibody (2 mg/ml) was diluted with the above described Tris HCl buffer and added to the reaction mixture. The results were corrected for non-enzymatic hydrolysis (3% or less in all experiments). Inhibition is expressed as the percentage of enzyme or sample control. IC50 values were determined graphically from plots of percentage inhibition versus log concentration of thielocin A1β.
STATISTICAL ANALYSIS
The significance of the difference between the mean of the control and that of the drug treated group was evaluated using Student’st test.
Results
SURVIVAL RATE IN RATS WITH DCA INDUCED ACUTE PANCREATITIS
To clarify the severity of DCA induced pancreatitis in rats, the survival rate was examined (fig 1). In the 1% DCA pancreatitis group, survival rate was 93% (n=28) at eight hours after the induction of pancreatitis. Even 24 and 48 hours after the induction of pancreatitis, the survival rates were relatively high (61% and 32%, respectively). In the 10% DCA pancreatitis group, however, the survival rate was only 7% (n=15) at five hours after the induction of pancreatitis and at eight hours, there were no survivors. Because of the lack of a sufficient number of blood samples for the 10% DCA pancreatitis group, the time course changes of amylase activity in plasma as well as PLA2 activity in plasma and ascites from the 1% or 10% DCA induced pancreatitis groups were observed for up to four hours.

: Survival rate in DCA induced acute pancreatitis in rats. Pancreatitis was induced by 1% DCA (broken line) or 10% DCA (solid line). Each point represents the percentage survival rate of 28 animals with 1% DCA pancreatitis or that of 15 animals with 10% DCA pancreatitis.
TIME COURSE CHANGES OF AMYLASE ACTIVITY IN PLASMA
There was no significant difference in plasma amylase activity between the sham control group and the saline control group for up to four hours. However, in the 1% and 10% DCA pancreatitis groups, plasma amylase activity was notably increased at one hour after the induction of pancreatitis. Thereafter, amylase activity gradually decreased (fig 2). At four hours after the induction of pancreatitis, there was no significant difference in amylase activity between the 1% or 10% DCA pancreatitis group and saline control (0% DCA) group.

: Time course changes of amylase activity in plasma from DCA induced acute pancreatitis in rats. Each point represents mean (SEM) from three to six animals.*p<0.05, **p<0.01 versus 0% DCA pancreatitis group. Open circles, sham controls; closed circles, 0% DCA pancreatitis; closed triangles, 1% DCA pancreatitis; closed squares, 10% DCA pancreatitis.
TIME COURSE CHANGES OF PLA2 ACTIVITY IN PLASMA AND ASCITES
There was no significant difference in plasma PLA2activity between the sham control group and the saline control group for up to four hours. At one hour after the induction of pancreatitis, plasma PLA2 activity was significantly increased in the 1% DCA pancreatitis group. However, there was subsequently no significant difference in plasma PLA2 activity between the 1% DCA pancreatitis group and the saline control group. In the 10% DCA pancreatitis group, PLA2 activity increased greatly up to four hours (fig 3A). Even two hours after the 10% DCA injection, plasma PLA2 activity was elevated to 21 times that of the normal group.

: Time course changes of (A) plasma PLA2activity and (B) ascites PLA2 activity in DCA induced acute pancreatitis in rats. Each point represents mean (SEM) from three to six animals. †p<0.05 versus sham control group, *p<0.05, **p<0.01, ***p<0.001 versus 0% DCA pancreatitis group. Open circles, sham controls; closed circles, 0% DCA pancreatitis; closed triangles, 1% DCA pancreatitis; closed squares, 10% DCA pancreatitis.
In ascites from the 1% and 10% DCA pancreatitis groups, however, PLA2 activity increased notably for up to four hours (fig3B). PLA2 activity in ascites from the saline control group showed a significant increase in comparison with that of the sham control group at one and four hours after the operation. Furthermore, the volume of ascites in the saline control group was also significantly higher than that of the sham group at one and four hours after the operation (table 1). With 10% DCA pancreatitis, as well as 1% DCA pancreatitis, the volume of ascites was notably higher than that of saline control group at one and four hours after the induction of acute pancreatitis.
Volume of ascites in DCA induced acute pancreatitis in rats
ALT AND AST ACTIVITIES IN PLASMA
There was no significant difference between the sham control group and the saline control group (table 2). In the 10% DCA pancreatitis group, but not the 1% DCA pancreatitis group, both AST and ALT were notably increased at one hour after the induction of pancreatitis. In the sham control group, AST and ALT activities were slightly higher than those of the normal group at one hour after the operation, possibly due to surgical trauma.
ALT and AST activities in plasma from rats with DCA induced acute pancreatitis
INHIBITORY EFFECT OF THIELOCIN A1β ON PLA2 ACTIVITY IN PLASMA AND ASCITES
In order to clarify the type of PLA2 in plasma or ascites from DCA induced acute pancreatitis in rats, the effect of thielocin A1β,5 ,23 a PLA2-II specific inhibitor, was investigated (fig 4). As already described, even one hour after the induction of pancreatitis with 10% DCA, PLA2 and amylase activities reached almost maximum levels. Therefore, we examined ascites and plasma from 10% DCA induced pancreatitis one hour after its induction. Thielocin A1β showed dose dependent inhibitory activity against plasma PLA2activity (IC50 = 0.066 μM). Furthermore, 10 μM of thielocin A1β inhibited 96 (2)% of the plasma PLA2activity. These results indicated that the PLA2 activity in plasma from rat DCA induced acute pancreatitis was mainly PLA2-II. On the other hand, thielocin A1β at 100 μM reduced the PLA2 activity in ascites by 21 (1.5)%. Further investigation was thus done on the PLA2 activity in ascites from DCA induced pancreatitis in rats.

: Inhibitory effect of thielocin A1β on PLA2 activity in plasma (closed circles) and ascites (open circles) from rat 10% DCA induced pancreatitis. PLA2activities of plasma and ascites controls (i.e. 100%) were 463.2 (99.1) and 1264.5 (55.8) pmol/min/ml, respectively. Each point represents mean (SEM) of six samples, each performed in duplicate.
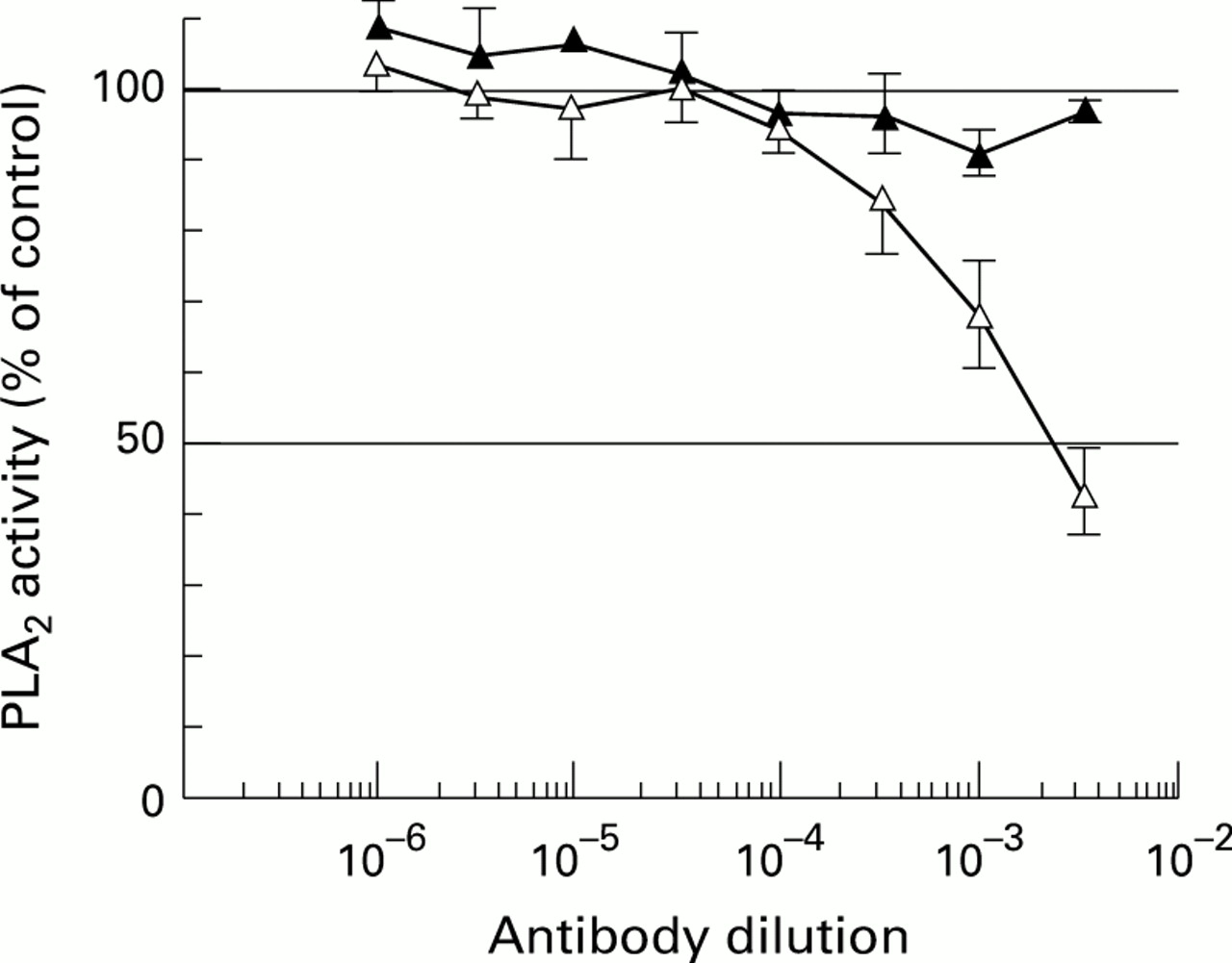
INHIBITION OF PLA2 ACTIVITY IN PLASMA AND ASCITES BY ANTI-PLA2-I ANTIBODY
In order to clarify the type of PLA2 in ascites, we carried out an inhibition study of the PLA2 activity with a specific polyclonal antibody against rat PLA2-I.29 Figure 5 shows that the antibody specifically inhibited the purified rat pancreatic PLA2activity belonging to PLA2-I but did not affect the activity of purified rat platelet PLA2 belonging to PLA2-II. In addition, this antibody inhibited 57 (6)% of the PLA2 activity in the ascites from DCA induced pancreatitis (fig 6). These results indicated that almost half of the PLA2 activity that was found in the ascites from rat DCA induced acute pancreatitis was PLA2-I.

: Inhibition of rat PLA2-I (open circles) and PLA2-II (closed circles) by anti-PLA2-I antibody. Each point represents mean (SEM) of triplicate determinations, each performed in duplicate and expressed as percentage of control (i.e. without the antibody = 100%).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
: Inhibition of PLA2 activity in plasma (closed triangles) and ascites (open triangles) from 10% DCA induced rat acute pancreatitis by anti-PLA2-I antibody. Each point represents mean (SEM) of six samples, each performed in duplicate and expressed as percentage of control (i.e. without the antibody = 100%).
Discussion
PLA2-I is synthesised and secreted by pancreatic acinar cells as an enzymatically inactive pro-PLA2-I, which normally is activated by trypsin in the lumen of the duodenum. Although its trigger mechanism is unknown, PLA2-I has long been considered to be one of the important digestive enzymes that destroys pancreatic tissue, resulting in acute pancreatitis.14 The injection of 0.15 ml of a 10% solution of DCA into the pancreatic duct induces severe, haemorrhagic, and necrotising pancreatitis, with a high mortality rate.19 In the present study, amylase activity in plasma was noticeably increased even one hour after the induction of pancreatitis. Furthermore, the survival rate was only 7% at five hours after the induction of pancreatitis. This exceptionally severe form of experimental pancreatitis was chosen because the corresponding indication in humans would be a severe, life threatening pancreatitis.
A novel finding in the present study is that the plasma PLA2 activity in rat DCA induced acute pancreatitis was equivalent to PLA2-II. These results largely support those obtained by Nevalainen et al 30 who showed that the catalytic activity of PLA2 in serum samples from 36 patients with severe acute pancreatitis is significantly correlated with the concentration of PLA2-II but not with that of PLA2-I. Furthermore, the remarkable increase in PLA2-II activity in plasma in the 10% DCA pancreatitis group but not in the 1% DCA pancreatitis group correlates well with the high mortality observed in the 10% DCA pancreatitis group. These results indicated that a significant increase of PLA2-II activity in plasma may contribute to high mortality in the acute phase of rat DCA induced acute pancreatitis.
The cellular source of PLA2-II circulating in blood plasma in inflammatory diseases is unknown, although a number of interesting hypotheses have been presented. A hepatocyte cell line cultivated in vitro was found to secrete PLA2-II into the culture medium when stimulated with cytokines, and therefore, it was proposed that PLA2-II might represent an acute phase protein secreted by liver cells.31 Furthermore, Nevalainen et al 32 recently reported that PLA2-II circulating in blood plasma originates in hepatocytes. Hepatic complication was therefore suggested to clarify the relationship between high mortality and the remarkably increased plasma PLA2-II activity in 10% DCA induced acute pancreatitis. Indeed, the plasma activities of both ALT and AST were significantly increased at one hour after the induction of 10% DCA pancreatitis. Our results were also supported by a report from Closa et al 33 who showed that rats with sodium taurocholate induced acute pancreatitis showed increases in plasma PLA2activity concomitant with inflammatory lung injury, and a portocaval shunt could prevent the metabolic changes and ameliorate the inflammatory process in the lung, suggesting that the liver plays an active role in the systemic inflammatory response to acute pancreatitis. These observations indicate that the increase in PLA2-II activity in plasma is associated with the development of hepatic complications in acute pancreatitis.
In striking contrast to the plasma PLA2-II activity, 57% of the PLA2 activity in ascites is due to PLA2-I. Furthermore, the remarkable increase in PLA2 activity in ascites one hour after induction of pancreatitis correlated well with the increase in the volume of ascites at one hour in the present study. Furthermore, in piglets with haemorrhagic pancreatitis, early chemical peritonitis was characterised by severe inflammation of the peritoneum with subsequent destruction of the mesothelial cells, leading to denudation of the underlying connective tissue.34 Moreover, PLA2, when instilled into the peritoneal cavity in the presence of pancreatitic ascites, has the ability to increase peritoneal permeability to dextran molecules in healthy as well as in pancreatitic rats.35These findings suggest that early increase in PLA2-I in ascites may contribute to the increased peritoneal permeability observed in the early stage of DCA induced pancreatitis in rats. In contrast, the notable accumulation of ascites at four hours in both the 1% and 10% DCA groups implies that PLA2-II in the circulation is not a critical factor for the increased peritoneal permeability.
In conclusion, increased PLA2-II activity in plasma could be involved in high mortality via hepatic complication, and increased PLA2-I activity in ascites might play a crucial role in the accumulation of ascites due to the augmentation of peritoneal permeability.
Acknowledgments
We wish to thank Dr Kouji Nomura, Shionogi Research Laboratories, for the generous supply of rat PLA2-I specific polyclonal antibody.