Article Text

Abstract
Background—Concentrations of pro-inflammatory cytokines are increased in the intestinal mucosa of patients with active inflammatory bowel disease (IBD). Polymorphonuclear neutrophil granulocytes (PMN) are the most abundant cell type in intestinal lesions in IBD. Interleukin 10 (IL-10) is an important contra-inflammatory cytokine which induces downregulation of pro-inflammatory cytokines.
Aims—To investigate whether PMN from patients with IBD or infectious colitis, respectively, secrete increased amounts of pro-inflammatory cytokines and can be regulated by IL-10.
Methods—Secretion (ELISA) as well as corresponding mRNA levels (semiquantitative RT-PCR) of pro-inflammatory cytokines (IL-1β, TNF-α) and of IL-1 receptor antagonist were assessed in peripheral PMN.
Results—PMN from patients with IBD are primed to secrete enhanced amounts of pro-inflammatory cytokines accompanied by detection of corresponding mRNAs in comparison with normal controls. This finding is not specific for IBD but rather reflects intestinal inflammation in general. IL-10 markedly inhibited pro-inflammatory cytokine secretion as well as corresponding mRNA concentrations.
Conclusions—PMN are an important source of pro-inflammatory cytokines in patients with intestinal inflammation and can be downregulated by IL-10.
- granulocytes
- interleukin 1β
- interleukin 10
- inflammatory bowel disease
- intestinal immunity
- inflammation
- neutrophils
- tumour necrosis factor α
Statistics from Altmetric.com
- granulocytes
- interleukin 1β
- interleukin 10
- inflammatory bowel disease
- intestinal immunity
- inflammation
- neutrophils
- tumour necrosis factor α
Polymorphonuclear neutrophils (PMN) are the first cells to migrate into tissue sites as part of the host defence system. Their extensive homing into areas of mucosal inflammation led to the development of imaging techniques in which radiolabelled PMN are used for semiquantitative assessment of clinical activity in inflammatory bowel disease (IBD).1-4 The homing process is thought to be mediated by specific adhesion molecules expressed on endothelial cells as well as on PMN.5-7 The studies by Saverymuttuet al also suggested that neutrophils which have entered the gut wall will transmigrate into the bowel lumen and will then be excreted with stools.3 During their migration through the intestinal mucosa PMN are thought to unload a host of mostly non-specific inflammatory mediators including superoxide anions, lipid mediators, and proteases.8 ,9
Recent studies have demonstrated the ability of peripheral monocytes as well as intestinal macrophages from patients with IBD to secrete enhanced amounts of pro-inflammatory cytokines.10-18Interleukin 1 (IL-1) and tumour necrosis factor α (TNF-α) may be particularly important for inducing and sustaining intestinal inflammation in IBD.10-13 16-19 Increased levels of TNF-α have also been reported in the stools of children with active Crohn’s disease.20 The increased production of pro-inflammatory cytokines in the intestinal mucosa is thought to be a pivotal factor in the pathogenesis of intestinal inflammation in IBD.9 ,16-19
It has been shown that PMN from patients with active IBD are primed for activation.21-23 The earlier reports of a decreased capacity to secrete superoxide anions in vitro may be attributed to the influence of separation techniques24 or to the fact that circulating stimulatory substances in the serum samples of these patients have already partially exhausted their capacity for superoxide anion secretion.25 26 Moreover, it has been demonstrated that elastase, which is a marker enzyme for activated PMN, is increased in the stools and intestinal mucosa of patients with IBD.27 ,28
It has been established that normal PMN have the capacity to secrete pro-inflammatory cytokines in vitro29-31 which can serve as specific mediators in enhancing immune activation both in a feed back loop to the phagocyte level and by contributing to T and B lymphocyte activation.30 We therefore investigated the capacity of peripheral PMN of patients with IBD as well as infectious colitis to secrete IL-1β, TNF-α, and IL-1ra. We found an increased capacity to secrete pro-inflammatory cytokines, whereas in vitro secretion of IL-1ra was not deficient in PMN from patients with bowel inflammation in comparison with normal cells.
The contra-inflammatory cytokine interleukin 10 (IL-10) is a potent mediator for downregulation of monocyte/macrophage activation and for the induction of lymphocyte differentiation.32-37 Recent studies indicate that IL-10 can suppress the release of IL-1β and TNF-α by PMN and induces secretion of IL-1ra.33 ,34 ,37 ,38 Moreover, IL-10 may be an important novel drug for anti-inflammatory therapy in IBD.37 ,39 We found that IL-10 effectively suppresses the release of both IL-1β and TNF-α, as well as corresponding cytokine mRNA levels, and induces IL-1ra secretion by PMN without a difference between IBD and normal control cells in a dose dependent manner.
Methods
Fetal calf serum (FCS) was purchased from Gibco (Grand Island, New York, USA) or Sigma (St Louis, Missouri, USA). Cytokine enzyme linked immunosorbent assays (ELISA) were obtained from Genzyme/Virotech (Rüsselsheim, Germany) and from DPC Biermann (Bad Nauheim, Germany). Recombinant IL-10 was purchased from Genzyme. All other chemicals were obtained from Sigma if not specified otherwise.
PATIENTS
A total of 31 patients with active Crohn’s disease (CD) and 38 patients with active ulcerative colitis (UC) participated in the study between April 1994 and November 1996 (figures show the exact number of patients being investigated in the different experiments conducted). All patients with IBD attended the outpatient clinics of the Charité University Hospital in Berlin because of increased clinical activity. Increased activity was documented both by clinical assessment as well as disease activity indices (table 1). At the time blood was drawn 24 of 31 patients with CD and 36 of 38 patients with UC received treatment with oral salicylates (mesalazine (Salofalk, Claversal, or Pentasa); salazosulfapyridine (Azulfidine or Colopleon); olsalazine (Dipentum)). None of the patients received steroids and/or cytotoxic drugs, immunosuppressive agents or antibiotics at the time of the initial examination. Six patients with active IBD (three CD, three UC) who were started on steroids in a standard dosage (60 mg prednisolone, tapered according to clinical response) were followed longitudinally. All patients with IBD underwent sigmoidoscopy or colonoscopy for routine clinical evaluation and only those with at least moderate activity by endoscopic assessment were included. Patients with ulcerative colitis were required to have at least 20 cm of inflamed mucosa by endoscopy. Crohn’s disease patients were required to show inflammatory lesions in the colon (with and without manifestations in the small bowel) (table1). Infection or parasites were excluded by stool cultures, microscopic stool examination, and serology.
Patient characteristics
Patients with IBD kept daily diaries for one week after the examination in which they recorded the occurrence of specific symptoms used to calculate the Crohn’s Disease Activity Index (CDAI) or the Clinical Colitis Activity Index (CAI), respectively.40 ,41 The CDAI provides a calculated composite score which incorporates eight items: number of liquid or very soft stools, general well being, abdominal pain, abdominal mass, haematocrit, body weight, extraintestinal manifestations, and use of opiate receptor stimulating drugs for the control of diarrhoea. Higher scores indicate more disease activity. Patients with a score below 150 are considered to be in clinical remission and scores above 450 reflect severely active Crohn’s disease. The CAI incorporates seven clinical items: number of stools, blood in stools, investigator’s global assessment of symptomatic state, body temperature due to colitis, abdominal pain, extraintestinal manifestations, and erythrocyte sedimentation rate. Patients with a CAI of 4 or less are considered to be in remission. Two patients with CD and one patient with UC did not have active disease with regard to the clinical score and results were excluded from the study analysis.
Blood was drawn after informed consent and the study was granted prior approval by the institutional review board. Normal controls (NC, n = 24) were age and sex matched healthy volunteers. Patients with bacterial infectious colitis (IC, n = 9) who were admitted to hospital because of severe symptoms and who showed mucosal inflammation by sigmoidoscopic assessment were used as an additional control group (table 1).
HUMAN POLYMORPHONUCLEAR NEUTROPHILS
Human PMN were separated from blood as previously described.42 ,43 In short, peripheral blood containing 1000 U heparin/10 ml was diluted 1:2 with dextran (Pharmacia, Freiburg, Germany, 3% in 0.85% sodium chloride solution). After sedimentation of cells, PMN were pelleted (400 g × 5 min) and the remaining erythrocytes twice lysed in 0.2% sodium chloride solution. After 20 seconds the sodium chloride concentration was increased to 0.9% by isovolumetric addition of 1.6% sodium chloride. Cells were then pelleted (400 g × 5 min) and layered over Lymphoprep (Nycomed, Oslo, Norway) (SG = 1.077) and were centrifuged for 30 minutes at 500 g. Cells from the interphase were discarded and the PMN pellet resuspended and washed in phosphate buffered saline. Viability was more than 98% (trypan blue) and the cell suspension contained more than 97% granulocytes (as assessed by morphology in Pappenheim’s stain). Monocyte contamination was excluded by double staining for esterases (α-naphthylacetate esterase and chloracetate esterase) and by the random assessment of cell preparations by electron microscopy.44 Moreover, human monocytes secrete interleukin-10 when stimulated with lipopolysaccharide which was not found in PMN culture supernatants.
PMN CULTURES AND CYTOKINE ASSAYS
PMN were cultured at a concentration of 1 × 106cells/ml in RPMI 1640 (supplemented with 10% FCS, 1% pyruvate, 100 U/ml penicillin and 50 μg/ml gentamicin). Cells were cultured in 24 well plates (Nunc, Roskilde, Denmark) with or without lipopolysaccharide (10 μg/ml) in the presence of different concentrations of IL-10 (if applicable). After 24 hours supernatants were separated from cells by centrifugation, snap frozen and stored at –70°C until determination of cytokine levels.
Supernatant concentrations of TNF-α, IL-1β and IL-1ra were assessed using a specific sandwich enzyme linked immunosorbent assay (ELISA). All samples were analysed in duplicate or triplicate. The amount of cross reactivity was assessed by comparison with the concentration yielding a 50% inhibition of binding. Specificity of the ELISAs was demonstrated by the finding that up to 100 pg/ml of IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12, G-CSF, GM-CSF and TNF-α showed no significant cross reactivity or interference. Sensitivity levels were between 2.0 pg/ml (IL-1β) and 4.0 pg /ml (TNF-α), intra- and inter-assay precision variability was below 3%.45Repeated thawing and freezing of the cultured cells, which would release cytokines stored in the cytoplasm, did not increase the cytokine levels significantly. Therefore all cytokines synthesised were secreted into the supernatants.
REVERSE TRANSCRIPTION POLYMERASE CHAIN REACTION (RT-PCR)
RNA was extracted by a modified guanidium-thiocyanate-phenol-chloroform extraction.46 In short, after removal of culture supernatants and washing, cells were lysed in guanidium solution (4 M guanidium-isothiocyanate, 25 mM sodium citrate, 0.5% sarcosyl, 0.1 M 2-mercaptoethanol, pH 7.0). After removal of cellular DNA 0.1 volumes of 2 M sodium acetate (pH 4.0) and one volume of H2O saturated phenol as well as 0.2 volumes of chloroform:isoamyl alcohol (49:1) were added sequentially. The mixture was incubated on ice (15 minutes) and then centrifuged (20 minutes × 12 000 g). The watery phase contained RNA which was precipitated overnight with one volume of 2-propanol (–20°C). After centrifugation (20 minutes × 12 000g) the pellet was again dissolved in 300 μl guanidium solution (composed as described above) and precipitated with 2.5 volumes of 100% ethanol. Following centrifugation the pellet was washed with 70% ethanol and RNA was resuspended in sterile water which had been treated with 0.1% diethyl pyrocarbonate.
For semiquantitative RT-PCR a cRNA standard (Perkin-Elmer Cetus) was used which contained numerous primer binding sites including those for TNF-α and IL-1β. The weight of approximately 300 bp allows easy discrimination between standard and sample derived DNA. Cellular RNA, prepared as described above, containing wild type (authentic) cytokine mRNA was mixed at constant concentrations with a series of samples containing increasing concentrations of synthetic cRNA standard. Aliquots of RNA mixture were reverse transcribed and amplified according to the method described by Wang et al.47 Preliminary experiments were carried out to ensure that equimolar concentrations of authentic and synthetic cytokine RNA are amplified with identical efficiency.37These studies of amplification kinetics were repeated at different concentrations and for each cytokine. It was decided to use 25 cycles for amplification of both cytokines because linear amplification and equal amplification efficiency could be achieved over the relevant concentration range from 24–27 cycles (TNF-α) and 23–27 cycles (IL-1β), respectively.37 The following primer pairs (Perkin Elmer Cetus) were used:
TNF-α: 5′-CAGAGGGAAGAGTTCCCC AG-3′ and 5′-CCTTGGTCTGGTAGGAG ACG-3′; IL-1β: 5′-AAACAGATGAAGTGCT CCTTCCAGG-3′ and 5′-TGGAGAACACC ACTTGTTGCTCCA-3′.
At the 5′ end the upstream primer was coupled to a fluorescein molecule. The PCR product was diluted in formamide (with 50 mM EDTA, 1:3) and analysed by denaturing 6% polyacrylamide gel electrophoresis.
EXPRESSION OF DATA
The symbol n refers to the number of experiments. All experiments were carried out three or more times. Normal distribution of data was evaluated by calculating Lilliefors’ probabilities based on the Kolmogorov-Smirnov test.48 ,49 Results are expressed as mean (SE) if distributed normally. Statistical significance of the differences for non-normally distributed data was tested with the Mann-Whitney U test or the Wilcoxon matched pairs test, respectively,49 ,50 and results are given as mean with interquartile percentiles.50
Results
INCREASED SECRETION OF INTERLEUKIN 1β AND TNF-α BY PMN FROM PATIENTS WITH INTESTINAL INFLAMMATION
Spontaneous secretion of IL-1β and TNF-α by PMN was seen only at low levels (table 2). Lipopolysaccharide stimulated secretion of IL-1β (fig 1, table 2) from PMN from patients with Crohn’s disease (p = 0.0024) and ulcerative colitis (p = 0.0025) was higher than that from normal control cells. PMN from patients with infectious colitis also showed increased secretion of IL-1β into culture supernatants in comparison with normal controls (p = 0.015). In a similar fashion secretion of TNF-α (fig 2, table 2) into culture supernatants was increased by PMN from patients with Crohn’s disease (p = 0.0026), ulcerative colitis (p = 0.0093), and infectious colitis (p = 0.027) in comparison with normal controls. Time course experiments demonstrated that more than 70% of the IL-1β and the TNF-α was secreted during the first eight hours of culture and that after 24 hours maximum concentrations had been reached (data not shown).
Secretion of TNF-α, IL-1β and IL-1ra by PMN from patients with IBD, infectious colitis or normal controls
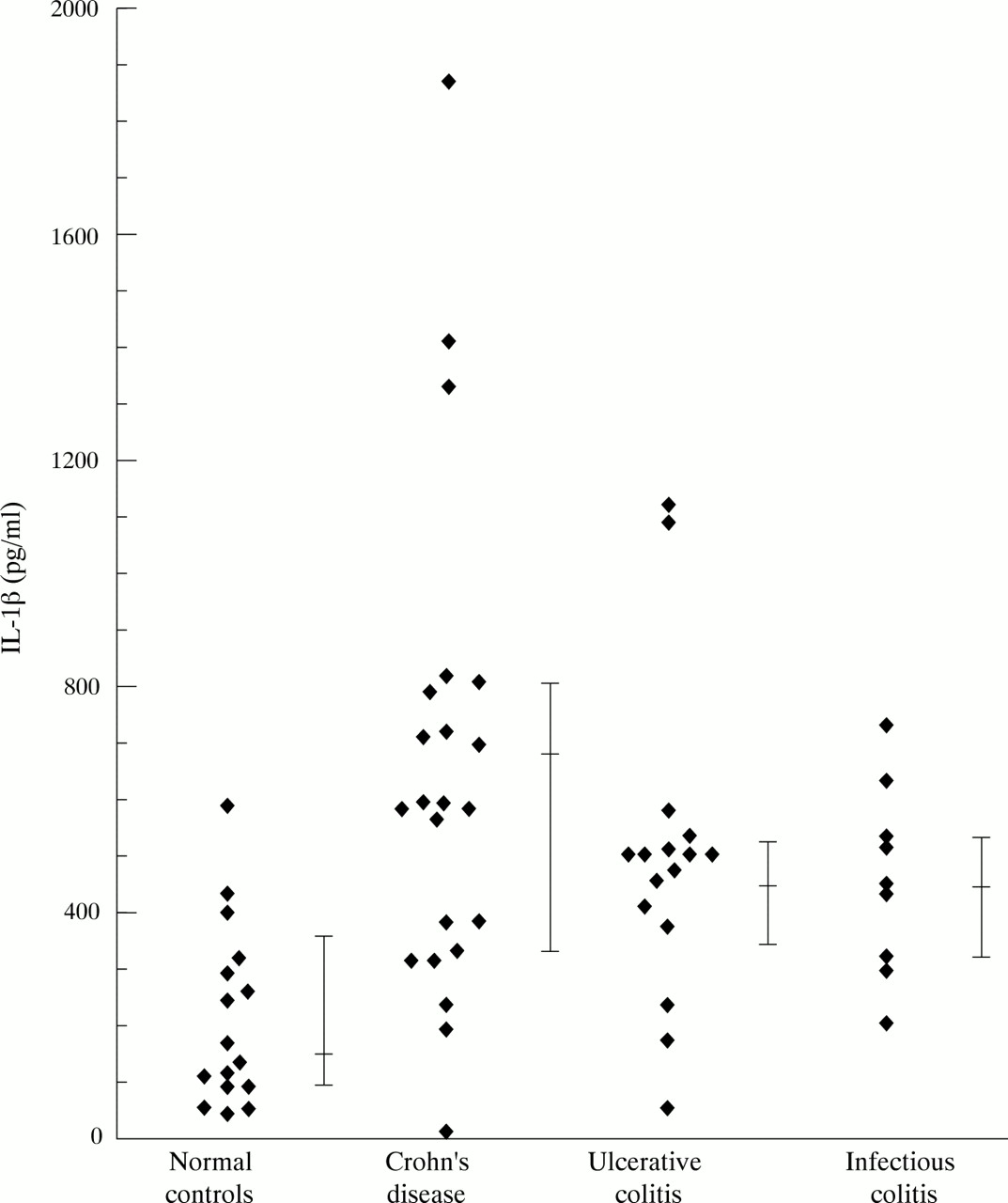
Lipopolysaccharide induced secretion of IL-1β by peripheral blood PMN.
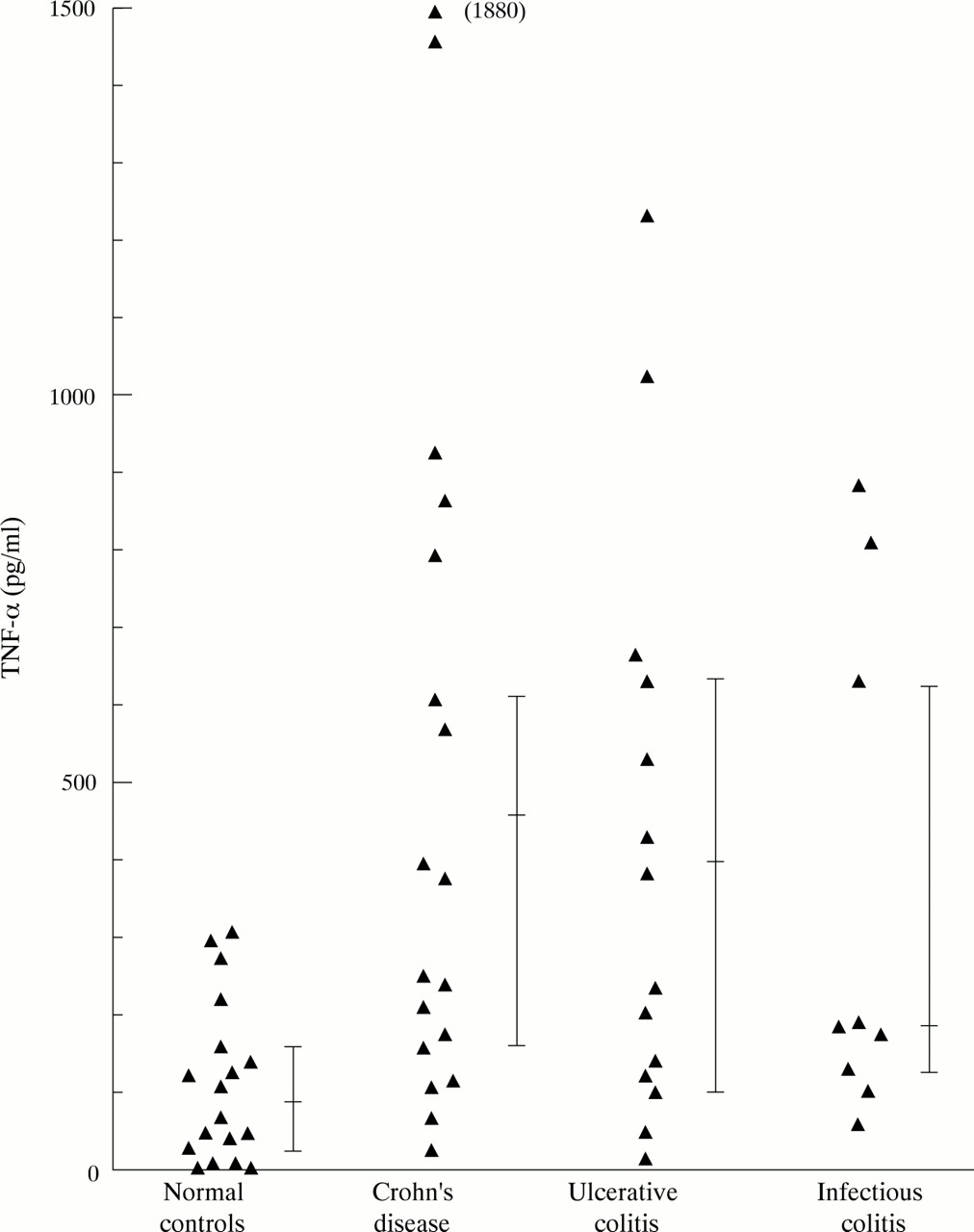
Lipopolysaccharide induced secretion of TNF-α by peripheral blood PMN.
SECRETION OF IL-1RA IN IBD AND NORMAL PMN
Polymorphonuclear neutrophils from patients with IBD and normal controls spontaneously secreted considerable amounts of IL-1ra. Stimulation with lipopolysaccharide greatly increased IL-1ra secretion, with both Crohn’s disease (p = 0.019) and ulcerative colitis PMN (p = 0.004) producing more than normal controls (fig 3, table 2). Secretion of IL-1ra did not differ between PMN from patients with infectious colitis and normal controls. No statistical differences in IL-1β/IL-1ra molar ratios were seen between IBD and normal control PMN (data not shown).

Lipopolysaccharide induced secretion of IL-1ra by peripheral blood PMN.
SECRETION OF PRO-INFLAMMATORY CYTOKINES BY PERIPHERAL PMN DURING LONGITUDINAL FOLLOW UP
Six patients with IBD were followed into steroid induced remission. All but one patient showed a substantial decrease in the secretion of TNF-α (fig 4) and IL-1β (data not shown). This patient suffered from Crohn’s disease. Although in clinical remission at the time of examination, he presented two weeks later with signs of high disease activity. Two patients with infectious colitis were re-examined after complete resolution of symptoms. TNF-α and IL-1β secretion levels declined well into the normal range (fig 4).
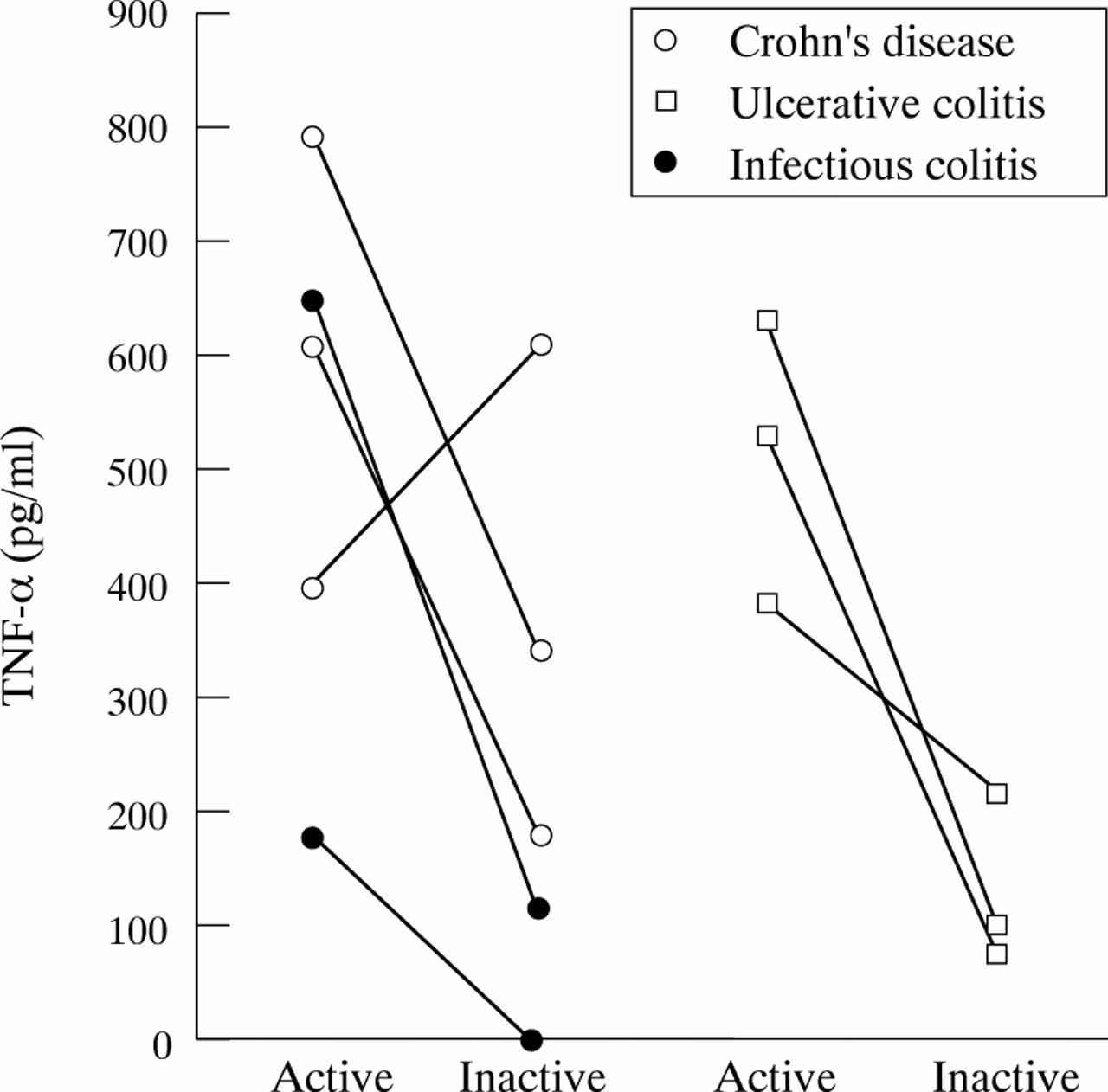
Longitudinal follow up of patients with mucosal inflammation.
DOWNREGULATION OF PMN PRO-INFLAMMATORY CYTOKINE SECRETION BY IL-10
Interleukin 10 is a potent contra-inflammatory mediator which downregulated pro-inflammatory cytokine secretion by PMN from both normal controls and patients with IBD in a dose dependent manner (table3, fig 5). No differences between IBD and normal control cells were seen with regard to the inhibitory capacity of IL-10. IL-10 downregulation of lipopolysaccharide induced TNF-α and IL-1β secretion, respectively, was accompanied by inhibition of corresponding mRNA levels (fig 6). IL-10 upregulated secretion of IL-1ra in a dose dependent manner (table 3). Neither IBD nor normal control PMN were secreting any detectable IL-10 (data not shown).
Regulation of TNF-α, IL-1β and IL-1ra secretion by IL-10 in PMN from patients with IBD and normal controls
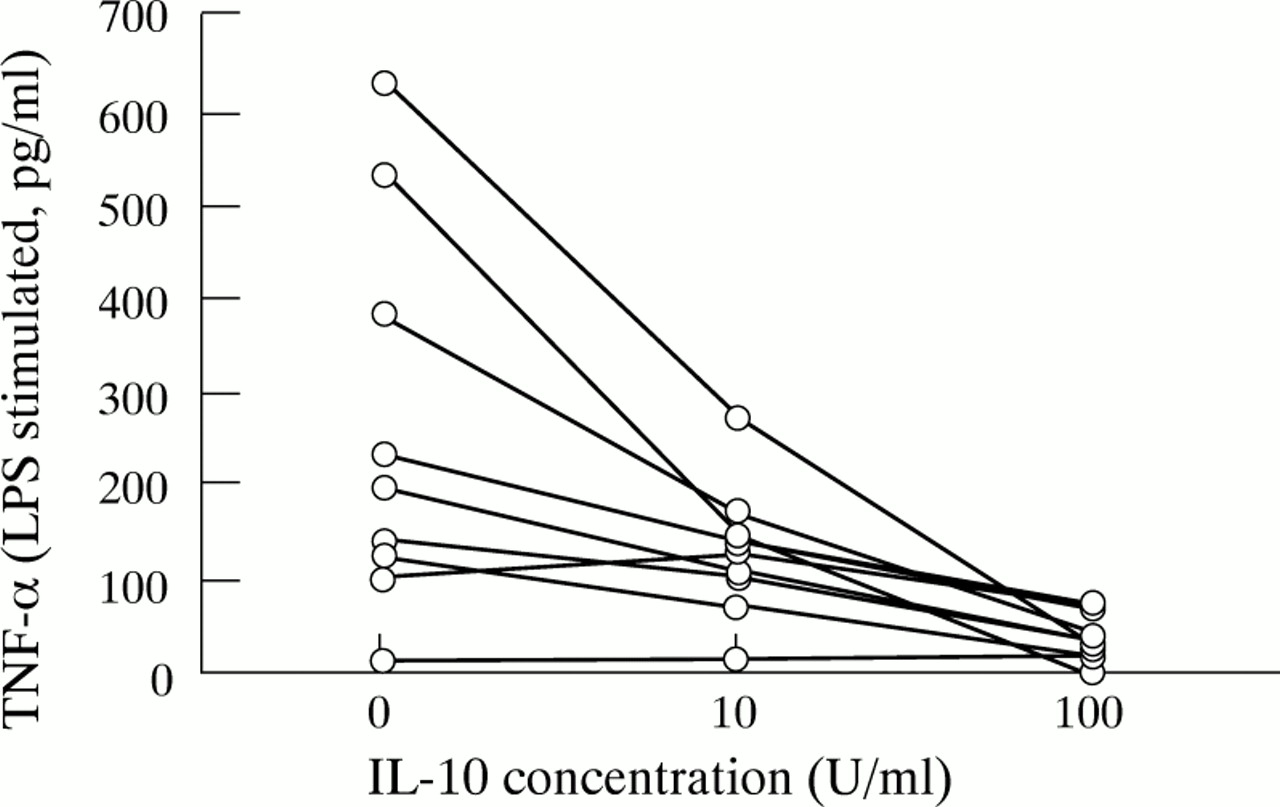
Downregulation of PMN pro-inflammatory cytokine secretion by IL-10.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Pro-inflammatory cytokine mRNA in PMN and regulation by IL-10.
Discussion
It has been suggested that both IL-1β and TNF-α may be important mediators involved in the initiation and perpetuation of intestinal inflammation in IBD. This study demonstrates that enhanced levels of both cytokines are released by purified peripheral polymorphonuclear neutrophils from patients with intestinal inflammation. Our main understanding has been that PMN will enter inflamed sites in the intestine in great numbers,1-4 ,51 ,52 unload mostly non-specific molecules including superoxide anions, lipid mediators and proteases, and then transmigrate through the bowel wall into the lumen.3 ,8This view of the neutrophil as a short lived cell which does not partake in specific immunoregulation was modified by recent studies which showed that: (1) normal PMN are capable of a stimulus dependent release of a host of cytokines in vitro,29-31 ,38 (2) corresponding cytokine mRNAs could be induced in PMN,30 ,31 ,38 and (3) activation of PMN prolongs their life span by preventing apoptosis (RD Lohmann and S Schreiber, unpublished data).
This study now shows that PMN carry the capacity to release increased amounts of IL-1β and TNF-α in vitro in active inflammatory bowel disease and infectious colitis. On stimulation with lipopolysaccharide, pro- inflammatory cytokine secretion is maximised in PMN from patients with intestinal inflammation. Increased amounts of pro-inflammatory cytokines which have been found in both the mucosa9-18and the faeces20 of patients with IBD may therefore be produced to a considerable part by PMN. Increased secretion of pro-inflammatory cytokines does not appear to be specific for IBD because PMN from patients with infectious colitis also showed increased release of IL-1β and TNF-α.
Interleukin 1 receptor antagonist can competitively block IL-1β from binding to its receptor. Secretion of IL-1ra as an anti-inflammatory molecule is therefore regarded as an important negative feedback mechanism to limit the potential pro-inflammatory effects of IL-1β.19 ,53 We did not observe any statistically significant differences in molar ratios of IL-1ra to IL-1β between PMN from normal controls and patients with IBD. Deficient secretion of IL-1ra does not therefore appear to play a pivotal role in PMN mediated immunoregulation.
The immunological activation of neutrophils can be divided into “priming” and “activation”.8 21 25 26 We hypothesise that our in vitro findings show an increased level of priming of circulating neutrophils leads to enhanced secretion of pro-inflammatory cytokines. Only upon activation by a secondary stimulus (lipopolysaccharide) may PMN from patients with intestinal inflammation secrete increased amounts of pro-inflammatory cytokines into the mucosa. As neutrophils are excreted with the faeces in patients with IBD, these cells may contribute to the increased levels of TNF-α which were found in the stools of children with IBD or infective diarrhoea.20 However, little is known about the in vivo penetration of lipopolysaccharide as a stimulus for cytokine release into the inflamed mucosa. Further studies which address the in vivo role of PMN in the inflamed mucosa are therefore necessary.
The detection of pro-inflammatory cytokine mRNA in PMN from both normal controls and patients with IBD indicate that cytokine production may be tightly regulated in PMN. Interleukin 10 is a powerful contra-inflammatory molecule inducing downregulation of macrophage as well as of T (h1) lymphocyte activation.32-37 Moreover, recent studies indicate a potential role for IL-10 as an anti-inflammatory therapeutic agent in inflammatory bowel disease, which inhibits pro-inflammatory cytokine secretion from mononuclear phagocytes in vitro and in vivo.37 ,39 Our observations indicate that IL-10 has the capacity to inhibit both pro-inflammatory cytokine mRNA levels and secretion in polymorphonuclear neutrophils from patients with IBD and induces secretion of IL-1ra.
The detection of cytokine mRNA in PMN already indicated that the inflammatory activation of these cells may be restricted by complex transcriptional controls. Activation of NF-κB may be a pivotal event in pro-inflammatory signal transduction and transcriptional regulation.54 ,55 Future studies will therefore address the role of cytokine transcription factors in PMN and other inflammatory cells in patients with IBD.
We have shown that neutrophils from patients with intestinal inflammation secrete high levels of TNF-α and IL-1β in comparison with normal control PMN after stimulation with lipopolysaccharide. Interleukin 10 is a powerful inhibitor of pro-inflammatory cytokine secretion and transcription by PMN in both patients with IBD and normal controls. As activation prolongs the short life span of PMN,56 we conclude that neutrophils may be important contributors to the initiation and perpetuation of mucosal inflammation and deserve further attention as competent immunoregulatory cells.
Acknowledgments
This work was supported by a grant from the Deutsche Forschungsgemeinschaft (SCH 512/1–2 to SS), by a European Training and Mobility of Research Network Grant (ERB-4061-PL-97–0389), by a stipend from the German Crohn’s and Colitis Foundation (DCCV) to SN, and by development grants from Genzyme GmbH and MFG.