Article Text

Abstract
Background—Although chemokines have been suggested to play an important role in Helicobacter pyloriassociated gastritis, few studies have investigated the role of chemokines other than interleukin 8 (IL-8) in gastric mucosa.
Aims—To investigate the expression and production patterns of various chemokines using gastric biopsy specimens.
Methods—In 192 patients, expression patterns of C-X-C chemokines (IL-8 and growth regulated α (GROα)) and C-C chemokines (regulated on activation, normal T cell expressed and presumably secreted (RANTES), monocyte chemotactic and activating factor (MCAF), macrophage inflammatory protein 1α (MIP-1α), and MIP-1β) were examined using reverse transcription polymerase chain reaction (RT-PCR) and enzyme linked immunosorbent assay (ELISA).cagA gene was identified using PCR.
Results—H pylori infection was associated with increased rates of expression of mRNA for IL-8, GROα, RANTES, and MIP-1α and with increased levels of mucosal IL-8 and GROα. IL-8 and GROα levels correlated with the density of H pylori in both the antrum and corpus. The levels of these chemokines correlated with cellular infiltration in the antrum but not the corpus. cagA gene positive H pyloriinfection was associated with increased rates of expression of mRNA for IL-8 and GROα and with increased levels of these chemokines.
Conclusion—H pylori infection is associated with increased expression rates and production of C-X-C chemokines (IL-8 and GROα), but not with increased production of C-C chemokines. Although H pylori infection is associated with increased C-X-C chemokines in the antrum and corpus, there is a difference in the inflammatory response between these two areas of the stomach.
- Helicobacter pylori
- chemokines
- interleukin 8
- GROα
Statistics from Altmetric.com
Helicobacter pylori is a Gram negative bacterium thought to cause chronic gastritis, peptic ulcer,1 ,2 and gastric cancer.3 ,4 H pylori associated gastritis is fundamentally a bacterial infection of the gastroduodenal mucosal surface and as such is characterised by mucosal infiltration of polymorphonuclear leucocytes (PMNs) and mononuclear cells (MNCs).5 Cure of the infection results in healing of the gastritis and notable reduction in mucosal inflammation. The migration and activation of inflammatory cells into the mucosa is thought to be related to expression of various cytokines.6-15 A number of studies have investigated interleukin 8 (IL-8), which is known to be a neutrophil chemotactic factor,16 ,17 and have shown an association between levels of H pylori infection and IL-8 mRNA11 ,12 ,14 or IL-8 protein.7-10 ,13 ,15
Cytokines showing leucocyte chemotactic activities, including IL-8, are referred to as chemokines.18 ,19 More than 20 members of the chemokine superfamily have been identified and shown to be involved directly or indirectly in inflammatory responses. Chemokines are classified into two major families on the basis of the arrangement of the first two of four conserved cysteine residues. The C-X-C chemokines consist of those members whose first two cysteines (C) are separated by one amino acid (X), while in the C-C chemokines the first two cysteines are adjacent to one another. In general, C-X-C chemokines mainly show chemotactic activities for neutrophils but not monocytes, whereas C-C chemokines show chemotactic activities for monocytes and lymphocytes but have little effect on neutrophils.20 The former group includes IL-8 and growth regulated (GRO), and the latter group includes regulated on activation, normal T cell expressed and presumably secreted (RANTES), monocyte chemotactic and activating factor (MCAF), macrophage inflammatory protein 1α (MIP-1α), and MIP-1β. Although IL-8 is closely associated with H pylori infection, few studies have investigated the relation between H pylori infection and chemokines other than IL-8.
To begin to clarify the relation between H pyloriinfection and chemokines, we examined chemokine expression and production patterns in gastric mucosa by reverse transcription polymerase chain reaction (RT-PCR) and enzyme linked immunosorbent assay (ELISA) using gastric biopsy specimens. In general, gastric mucosal inflammation caused by H pylori is more severe in the antrum than that in the corpus.21 ,22 We therefore investigated the expression of chemokines in the antrum and corpus in patients with and without H pylori infection.
Materials and methods
PATIENT POPULATION
A total of 192 patients (110 men and 82 women; age range 21–81 years, mean 54.7 years) were studied; 172 were consecutive symptomatic adults who underwent endoscopy between October and December 1995. In order to examine the relation between H pylori and gastric cancer, 25 patients with gastric cancer, who underwent endoscopy between January and May 1996, also participated in this study. No patient had received non-steroidal anti-inflammatory drugs, proton pump inhibitors, or antibiotics within the previous three months. There was no overlap between subjects in this and in our previous studies. Endoscopic findings included: normal mucosa, 10 patients; gastritis without ulcer, 86 patients; gastric ulcer, 41 patients; duodenal ulcer, 30 patients; and gastric cancer, 25 patients. Ninety four patients were smokers. Informed consent was obtained from all patients and the protocol was approved by the ethics committee of Kyoto Prefectural University of Medicine.
Four biopsy specimens were taken from both the antrum (pyloric gland area) and corpus (fundic gland area); two were used for histological examination, one for H pylori culture and cytokine protein measurement, and one for cytokine mRNA measurement. Biopsy specimens were taken at least 2 cm from ulcers or tumours in the cases of gastric ulcer and gastric cancer.
The presence of H pylori was determined by culture and histological examination. Patients were classified as H pylori positive if at least one of the examinations gave a positive result. Some of the H pylori positive patients who had gastric ulcer or duodenal ulcer were treated with omeprazole 20 mg once daily, amoxycillin 2000 mg twice daily, and clarithromycin 400 mg twice daily, for two weeks. Four weeks after cessation of treatment, the patients were examined endoscopically again and biopsy specimens were taken as before. 13C urea breath test was also performed, and successful eradication of H pylori was determined if all test results were negative.
HISTOLOGY
The biopsy specimens were fixed in 10% buffered formalin, oriented on edge, embedded in paraffin wax, cut in sequential 4 μm sections, and stained with haematoxylin and eosin (H&E) and modified Giemsa stains. We usually cut three to five serial sections for one specimen and multiple high power fields were examined. Virtually all specimens included surface epithelium and muscularis mucosa. Specimens were examined by one experienced histologist blinded to the patient’s clinical diagnosis or the characteristics of the H pyloristrain. The following features were evaluated: MNC infiltration, PMN infiltration, and density of H pylori. All variables were graded using the visual analogue scale graded from 0 to 3 for biopsy specimens from each site according to the Updated Sydney system.23 For analysing the relation between the histological findings and the expression patterns of chemokines, we used the biopsy specimens from a mucosal site adjacent to the site where the biopsy specimens for chemokine measurements were obtained. MNC infiltration was scored as follows: 0 = occasional lymphocytes and plasma cells at a level considered normal; 1 = mild focal increase in MNC numbers; 2 = definite infiltrate with MNCs isolated from each other; and 3 = diffuse MNC infiltrate. PMN infiltration was scored as follows: 0 = no neutrophils; 1 = infiltrate only in the lamina; 2 = infiltrate within the epithelium; and 3 = infiltrate within the foveolar lumen. The density of H pylori was scored based on the average density on the surface and the foveolar epithelium. If areas with widely different scores were present on the same specimen, an average based on the general evaluation of the sample was used by the observer. If intestinal metaplasia was present, only areas without metaplasia were evaluated for the presence of H pylori.
H PYLORI CULTURE
Biopsy specimens were placed in 1.5 ml of phosphate buffered saline (PBS; pH 7.4), and were immediately homogenised using a tissue homogeniser (Kontes, Vineland, New Jersey, USA). Ten ml aliquots of homogenate supernatant were spread on Skirrow agar medium containing 8% horse blood, vancomycin (10 mg/ml), polymyxin B (2.5 U/ml), and trimethoprim (5 mg/ml) and incubated at 37°C under microaerophilic conditions (AnaeroPack Systems; Mitsubishi Gas Chemical Co., Inc., Osaka, Japan) for up to five days. The organisms were identified asH pylori by Gram staining, colony morphology, and positive oxidase, catalase, and urease reactions. The strains were cultured again at 37°C under microaerophilic conditions in brain heart infusion broth (Nissui Co., Ltd., Osaka, Japan) containing 5% horse serum for 24 to 48 hours on a shaker, and aliquots were stored at −80°C in brain heart infusion broth containing 15% horse serum and 15% glycerol.
PREPARATION OF GNOMIC DNA FROM CLINICAL ISOLATES
The culture medium was centrifuged for five minutes at 10 000g and the bacterial pellets were washed three times with PBS (pH 7.4). The pellets were each suspended in 200 ml of distilled water and were boiled for 10 minutes. The supernatants, obtained by centrifugation (10 000 g for five minutes), were stored at −20°C until use as PCR templates.
PREPARATION OF RNA AND cDNA FOR CHEMOKINE mRNA MEASUREMENT
The biopsy specimens were immediately placed in guanidine isothiocyanate buffer, frozen on dry ice, and stored at −80°C until use. Samples were homogenised using a tissue homogeniser (Kontes), and total RNA was extracted by the acid guanidinium isothiocyanate-phenol-chloroform method as described previously.24 Aliquots (1 mg) of total RNA were incubated at 65°C for five minutes, chilled on ice, and reverse transcribed in a final volume of 10 ml of a solution containing 50 mmol/l Tris-HCl (pH 8.3), 75 mmol/l KCl, 3 mmol/l MgCl2, 10 mmol/l dithiothreitol, 200 mmol/l each of deoxyadenosine triphosphate, deoxycytidine triphosphate, deoxyguanosine triphosphate, and deoxythymidine triphosphate (Pharmacia Biotech AB, Uppsala, Sweden), 1 mmol/l oligo(dT)16 primer, 20 units RNAsin (Ribonuclease inhibitor; Toyobo Co., Ltd, Tokyo, Japan), and 100 units of MoMuLV RNase H− reverse transcriptase (BRL Co., Gaithersburg, Maryland, USA). The mixture was incubated at 43°C for 90 minutes, heated to 95°C for 10 minutes, and stored at −20°C until use.
POLYMERASE CHAIN REACTION
Bacterial DNA and cDNA from biopsy specimens were denatured by heating to 95°C for 10 minutes and cooled on ice for five minutes. Bacterial DNA and cDNA (5 ml) were added to 50 ml reaction mixtures containing 5 ml 10× PCR reaction buffer (500 mmol/l KCl, 100 mmol/l Tris-HCl (pH 8.8), 15 mmol/l MgCl2, 1% Triton X-100, 200 mmol/l each of deoxyadenosine triphosphate, deoxycytidine triphosphate, deoxyguanosine triphosphate, and deoxythymidine triphosphate, 200 nmol/l of each primer, and 1.0 unit Taq DNA polymerase (Boehringer, Mannheim, Germany), and H2O). Amplification was performed using a Takara PCR Thermal Cycler 480 (Takara Biomedicals, Otsu, Japan) for 35 cycles (for chemokine PCRs) or 30 cycles (for 16S rRNA and cagA PCRs), each of which consisted of one minute at 95°C for denaturation, one minute at 60°C (for chemokine PCRs) or 50°C (for 16S rRNA and cagAPCRs) for annealing, and one minute at 72°C for extension. The final cycle included an extension step for seven minutes at 72°C to ensure full extension of the product. Aliquots (10 ml) of each PCR product were analysed by electrophoresis on 1.5% agarose (Dojin Chemical Co., Ltd, Kyoto, Japan) gels containing ethidium bromide, and the bands were examined under ultraviolet light for the presence of the amplified DNA.
Oligonucleotide primers were designed based on previous studies.15 ,25-28 The sense and antisense primers specific for each chemokine were designed to include at least one intron, permitting distinction between amplified cDNA and possible contaminating residual gnomic DNA. Table 1 shows chemokine specific oligonucleotide primers. β-Actin and H pylori 16S rRNA were assayed as positive controls for chemokine measurement and for the presence of H pylori, respectively. In addition,cagA gene negative strains, G21, G50, and 2U (kindly provided by Dr J E Crabtree), were simultaneously assayed to check for contamination.
Human chemokine specific oligonucleotide primers for RT-PCR
CHEMOKINE PROTEIN MEASUREMENT
For chemokine protein measurement, we used the same biopsy specimens as used for H pylori culture. Biopsy specimens were placed in 1.5 ml of PBS (pH 7.4), and were immediately homogenised using a tissue homogeniser as described earlier. Aliquots of homogenate supernatants, obtained by centrifugation (10 000 g for 10 minutes), were stored at −80°C until assay for total proteins by a modified Lowry method and for chemokine protein by ELISA. IL-8, GROα, MIP-1α, and RANTES in biopsy homogenate supernatants were measured by ELISA using commercially available assay kits (Research and Diagnostic Systems, Minneapolis, Minnesota, USA). Assays were performed in duplicate according to the manufacturer’s instructions. In our laboratory, the ELISA sensitivities of IL-8, GROα, MIP-1α, and RANTES were approximately 10 pg/ml.
STATISTICS
The mucosal levels of IL-8, GROα, MIP-1α, and RANTES protein were expressed as pg/mg biopsy protein, and data were expressed as median and range. As histological scores are non-parametric, these scores are presented as median rather than mean values. The two tailed Fisher’s exact probability test was used to compare RT-PCR results between patient populations. This test was also used to compare differences of chemokine mRNA before and after treatment. Wilcoxon’s matched pairs test was used to compare differences of chemokine protein before and after treatment. The relation between histological findings and the production of chemokines was assessed by Spearman’s rank correlation coefficient. This test was also used to assess the relation between the production of IL-8 and GROα. For the remaining statistical analyses, the Mann-Whitney U test for non-parametric data was used. A p value of less than 0.05 was accepted as statistically significant.
Results
H PYLORI INFECTION AND ENDOSCOPIC AND HISTOLOGICAL FINDINGS
Of the 192 patients examined, 131 (68.2%) were positive forH pylori (table 2). Of the 123 patients with positive culture for H pylori, 114 (92.7%) were positive for thecagA gene.
H pylori infection and endoscopic findings
As expected specimens from patients with endoscopic normal mucosa without H pylori had no infiltration of MNCs or PMNs and specimens from those with H pylori infection showed typical changes with infiltration of PMNs and MNCs. There was no significant difference in density of H pylori between the antrum and the corpus. In contrast, both MNC and PMN infiltration was significantly more severe in the antrum than in the corpus in H pylori infection.
H PYLORI INFECTION AND EXPRESSION OF CHEMOKINE mRNA
H pylori infection was associated with increased rates of expression of mRNA for IL-8, GROα, RANTES, and MIP-1α in both the antrum and corpus (table 3). In contrast mRNA for MIP-1β and MCAF was detected in the majority of specimens independent of whetherH pylori infection was present or absent.
Expression of chemokine mRNA and H pylori infection
To reduce the variability because of possible differences in histology related to different diseases, we examined whether expression of these chemokine mRNAs was related to the histological findings in patients with only H pylori gastritis (table 4). As was true for the overall population, in the antrum both MNC and PMN infiltration scores were significantly higher in specimens positive for the expression of IL-8 and GROα mRNA. In the corpus, MNC infiltration score was similar in specimens positive or negative for expression of IL-8 or GROα mRNA. When mRNA for both IL-8 and GROα was expressed, the MNC and PMN infiltration scores were significantly higher in both the antrum and corpus (data not shown).
Relation between histological findings and chemokine profiles in H pylori positive patients with chronic gastritis
In both the antrum and corpus, the MNC infiltration score was significantly higher in specimens positive for the expression of MIP-1α mRNA than in those negative for the expression of this chemokine mRNA. No relation was found in either the antrum or corpus between C-C chemokine mRNA and PMN infiltration. H pyloridensity score was significantly higher in specimens positive for the expression of IL-8, GROα, or MIP-1α mRNA than in those negative for the expression of mRNA for these chemokines both in the antrum and corpus (table 4).
With regard to the relation between the expression of chemokine mRNA and the endoscopic findings in H pylori positive specimens, the positive rate of expression of IL-8 mRNA was significantly higher in the antrum than that in the corpus in patients with duodenal ulcer (86.7% versus 56.7%, p<0.05, n=30) which is consistent with the association of duodenal ulcer with antral predominant gastritis.
H PYLORI INFECTION AND PRODUCTION OF CHEMOKINES
mRNA levels of IL-8, GROα, RANTES, and MIP-1α were increased in H pylori infection and the level of production of these chemokines was examined at the protein level by ELISA in 100 patients, 76 of whom were infected with H pylori. There were no significant differences in sex or age distribution between these patients and the total patient population. The mucosal levels of IL-8 and GROα protein were significantly increased in H pylori infection in the antrum and corpus (median for IL-8 was 58 pg/mg versus 0 pg/mg, and 60.6 pg/mg versus 0 pg/mg for antrum and corpus, respectively; p<0.0001). For GROα the median was 157.3 pg/mg versus 0 pg/mg and 157.4 pg/mg versus 0 pg/mg for antrum and corpus, respectively (p<0.0001; fig 1). In addition, the mucosal levels of IL-8 and GROα protein were significantly higher in specimens positive for expression of IL-8 and GROα mRNA compared with those in specimens without expression of these transcripts (median for IL-8 was 84.5 pg/mg versus 0 pg/mg, and 88.5 pg/mg versus 0 pg/mg for antrum and corpus, respectively; p<0.0001). For GROα the median was 183.1 pg/mg versus 0 pg/mg and 180 pg/mg versus 96.4 pg/mg for antrum and corpus, respectively (p<0.0001). The mucosal level of GROα protein was significantly higher in cagA positive than in cagA negative specimens in the antrum (fig 1).

H pylori infection and production of GROα. Bars indicate median values for each group.
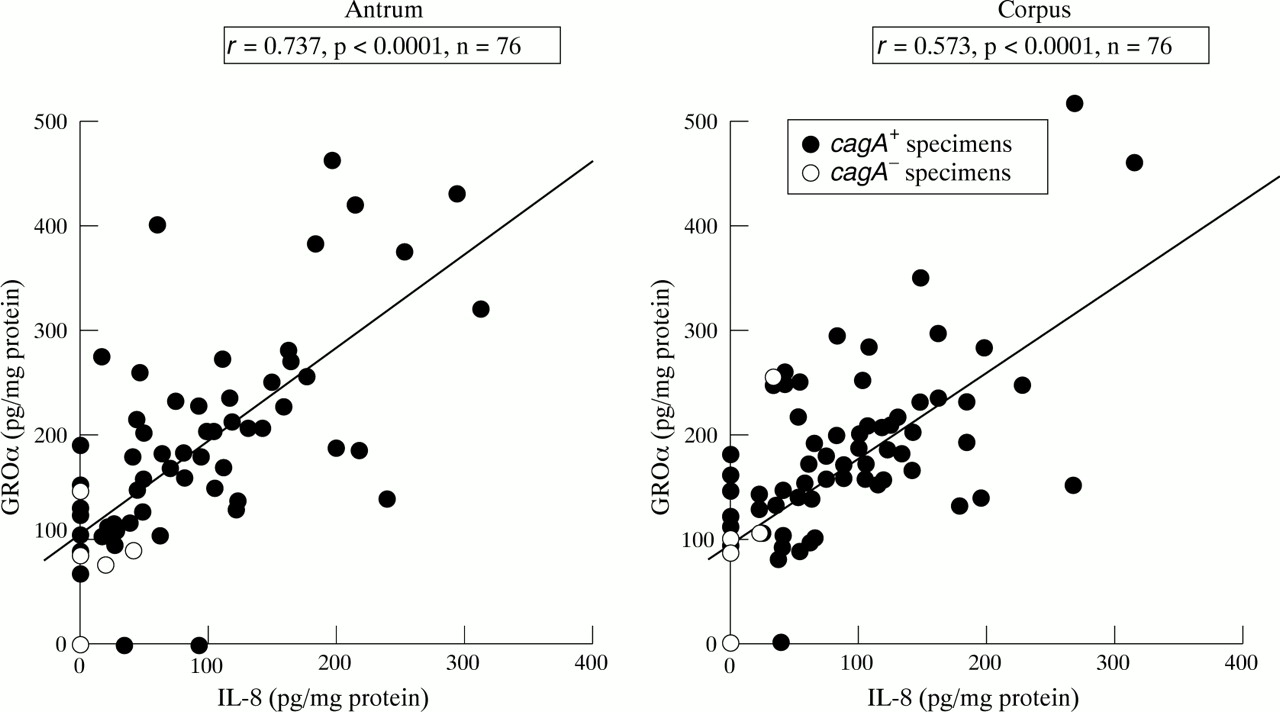
The mucosal level of IL-8 protein was significantly correlated with that of GROα protein both in the antrum and corpus (p<0.0001 by Spearman rank test; fig 2). There was no relation between the level of mucosal RANTES protein betweenH pylori infected and uninfected individuals (median 251.8 pg/mg versus 256.1 pg/mg and 286.4 pg/mg versus 293.3 pg/mg for antrum and corpus, respectively). In the antrum, the RANTES protein level was significantly higher in specimens positive for the expression of RANTES mRNA than in those negative by RANTES RT-PCR (median 282.3 pg/mg versus 228.8 pg/mg, respectively, p<0.05). The mucosal level of MIP-1α protein was below the threshold of sensitivity in all specimens.

Relation between mucosal levels of IL-8 and GROα protein in H pylori positive patients.
In the corpus, mucosal levels of IL-8 and GROα protein were significantly lower in patients with duodenal ulcer compared with those in patients with gastritis without ulcers (median for IL-8 of 10.1 pg/mg versus 90 pg/mg, and median for GROα of 102.4 pg/mg versus 167 pg/mg, for antrum and corpus respectively; p<0.05). On the other hand, the mucosal levels of IL-8 protein were significantly higher in patients with duodenal ulcer compared with those in patients with gastritis without ulcers in the antrum (median 102.3 pg/mg versus 59.9 pg/mg; p<0.05).
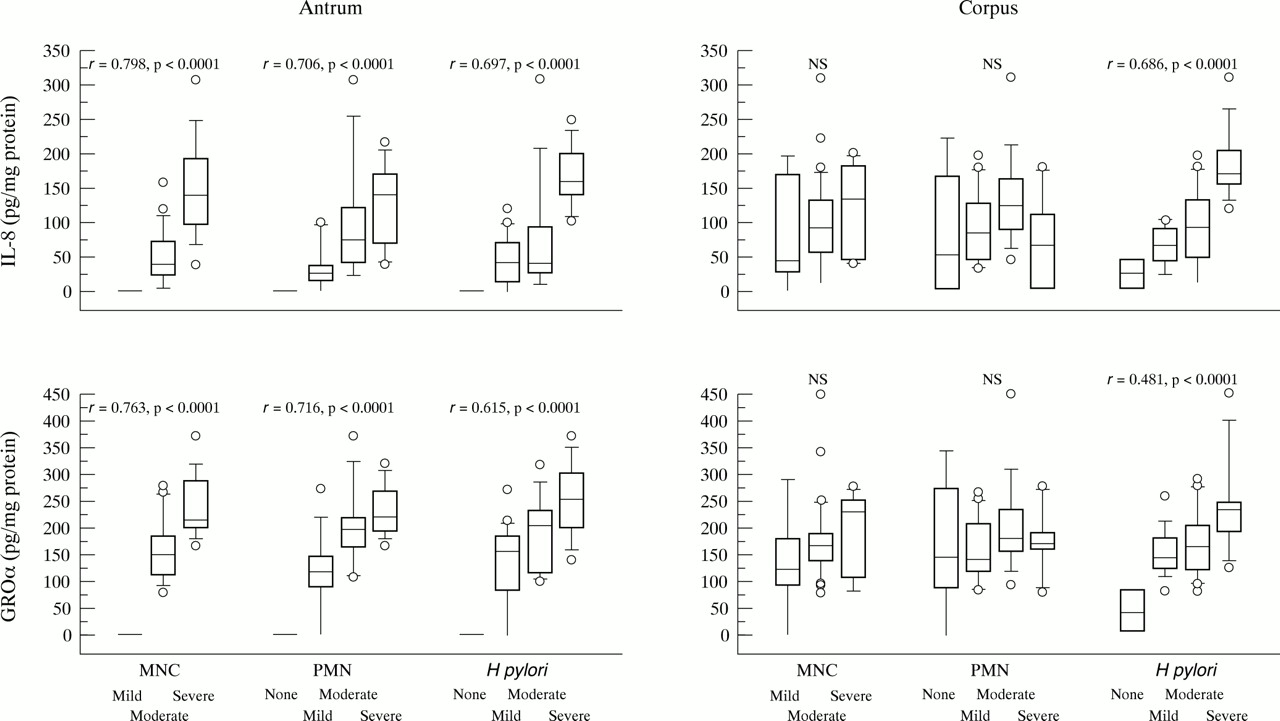
We examined the relation between the production of chemokines and histological findings in H pylori chronic gastritis. The mucosal levels of IL-8 and GROα protein were significantly correlated with infiltration of MNCs and PMNs in the antrum (fig 3). There was no significant difference in the corpus. The mucosal levels of IL-8 and GROα protein were significantly correlated with the density ofH pylori in both the antrum and corpus. H pylori positive specimens with diseases other than chronic gastritis also showed a similar tendency (data not shown).

Production of C-X-C chemokines and histological findings in H pylori positive patients with chronic gastritis are expressed as box plots. The ends of the bars indicate the 25th and 75th percentiles. The 50th percentile is indicated with a line, and the 10th and 90th percentiles are indicated with error bars. Data outside the 10th and 90th percentile are displayed using symbols. Correlation coefficients were calculated with the Spearman rank test.
EXPRESSION AND PRODUCTION OF CHEMOKINES AND ERADICATION OFH PYLORI
Twenty patients were treated with eradication therapy. Ten had gastric ulcer and 10 had duodenal ulcer; all patients were infected with H pyloripossessing the cagA gene. H pyloriinfection was successfully cured in 15 patients, seven with gastric ulcer and eight with duodenal ulcer. The positive rates of expression of IL-8, GROα, and MIP-1α mRNA decreased significantly following successful treatment of patients (data not shown). There was no significant change in the expression ofRANTES, MIP-1β, and MCAF mRNA. The mucosal levels of IL-8 and GROα protein fell below the level of detection in all but one and two patients, respectively (fig 4). The mucosal level of RANTES protein was not reduced even in successfully treated patients.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Production of GROα and eradication of H pylori. *p<0.005 by Wilcoxon’s matched pairs test. Before, before eradication therapy; after, after eradication therapy.
EXPRESSION AND PRODUCTION OF CHEMOKINES AND THEcagA GENE
As expected, the positive rate of expression of IL-8 mRNA was significantly higher in cagA gene positive strains (cagA + specimens) than in cagA gene negative strains (cagA − specimens) in the antrum (86.8% versus 44.4%, p<0.005). Similarly, the mucosal levels of IL-8 protein were also significantly increased incagA + compared withcagA − specimens in both the antrum and corpus (median 65.3 pg/mg versus 0 pg/mg and 73.7 pg/mg versus 10 pg/mg for antrum and corpus, respectively; p<0.05). The positive rate for expression of GROα mRNA was significantly higher incagA + than in cagA −specimens in the corpus (83.3% versus 44.4%, p<0.05). The mucosal level of GROα protein was significantly increased incagA + over that incagA − specimens in the antrum (median 173.2 pg/mg versus 83.0 pg/mg, p<0.001; fig 1). Expression of C-C chemokine mRNA did not differ between cagA + andcagA − specimens in either the antrum or corpus.
Discussion
In the present study, H pylori infection was associated with increased expression and production of not only the C-X-C chemokine IL-8 but also GROα. Furthermore, the expression of C-X-C chemokines significantly decreased in patients after successful eradication. These findings are consistent with the C-X-C chemokines playing important roles in H pylori associated gastric inflammation. The degree of cellular infiltration was however lower in the corpus than that in the antrum but there were no differences in expression of C-X-C chemokines between these areas, suggesting that other, as yet unidentified, factors modulate the mucosal response between C-X-C chemokine expression and cellular infiltration. It is of interest that the mucosal levels of C-X-C chemokines correlated significantly with the density of H pylori in both the antrum and corpus, whereas the mucosal levels of C-X-C chemokines were significantly correlated with cellular infiltration only in the antrum. However, even in the corpus, expression of both IL-8 and GROα mRNA was associated with significantly more severe MNC and PMN infiltration, suggesting that IL-8 and GROα may act cooperatively and that their synergistic action causes severe inflammation even in the corpus with low responsiveness to either chemokine alone. There are two types of IL-8 receptor: type I is IL-8 specific and type II has affinity for GROα.29 Further studies will be necessary to investigate whether the numbers of these receptors differ between the antrum and corpus and whether such a difference might be related to the differences in inflammatory response to H pylori between these two areas of the stomach.
With regard to C-C chemokines, the positive rate of expression of MIP-1α mRNA was significantly enhanced in H pyloripositive specimens. Furthermore, the positive rate of expression of MIP-1α mRNA significantly decreased after successful eradication. The MNC infiltration score was significantly higher in specimens positive for the expression of MIP-1α mRNA compared with those negative for its expression in both the antrum and corpus. Expression of MIP-1α mRNA was not associated with PMN infiltration. These data are in agreement with previous observations that MIP-1α has chemotactic activity for MNCs but has little effect on PMNs.20However, as production of MIP-1α protein was below the threshold of sensitivity in all patients, the role of this chemokine cannot be determined from this study.
While the positive rate of expression of RANTES mRNA was significantly higher in H pylori positive infection, there were no differences in protein levels. The mucosal level of RANTES protein was significantly higher in specimens positive for the expression of RANTES mRNA than in those negative for the expression of this mRNA in the antrum. Similar conflicting data have been reported for some other cytokines including IL-1α,13IL-7,14 ,15 and IL-10.14 ,15 These conflicts between protein and mRNA levels are possibly related to disturbances in the pathway from mRNA to protein expression, degradation of mRNA prior to translation into protein, or the fact that the amount of mRNA may not exceed the threshold required for protein production.
We found no correlation between RANTES mRNA and protein expression; the degree of MNC and PMN infiltration and the levels of expression of RANTES were not decreased after successful eradication therapy. These findings suggest that RANTES may not play a role in MNC or PMN infiltration in H pylori infection. RANTES does not show any chemotactic activity on PMNs,30 but shows chemotactic activity on T lymphocytes31 and eosinophils.30 ,32 Further studies will be necessary to investigate the relation between infiltration by these cells and RANTES expression in gastric mucosa.
MIP-1β and MCAF mRNA were detected in the majority of specimens regardless of the presence or absence of H pylori. It has been thought that chemokines are not expressed in normal cells, but as the stomach is constantly exposed to various stimuli including dietary components, these influences may induce expression of chemokine mRNA.
In the antrum, the mucosal levels of IL-8 and GROα protein, the positive rate of expression of IL-8 mRNA, and H pyloridensity were all significantly higher in patients with duodenal ulcer than in those with other diseases. In contrast, in the corpus, the mucosal levels of IL-8 protein and H pylori density were significantly lower in patients with duodenal ulcer than in those with other diseases. It is not clear what relation may exist between these observations and the formation of duodenal ulcer disease in this study.
Finally, we examined the cagA gene. Recent studies usingcagA isogenic mutant strains indicated that thecagA gene was not essential for induction of IL-8, but was a phenotypic marker for H pylori strains capable of inducing IL-8.33 ,34 Genes coexpressed with the cagAgene, present in the upstream sequence of the cagA gene, are thought to contribute to IL-8 induction.34 ,35 Recently, a region of approximately 40 kilobases responsible for virulence that contains the cagA genes was reported and is referred to as the “cag pathogenicity island”.36 cagA positive strains are, therefore, regarded as strains possessing the cag pathogenicity island. In agreement with previous studies,13-15 expression and production of IL-8 were enhanced by cagA positive strains compared withcagA negative strains. In this study the positive rate of expression of GROα mRNA in the corpus and the mucosal level of GROα protein in the antrum was also significantly higher in infection withcagA positive H pylori. There was no association between C-C chemokines and the cagA gene.
In conclusion, H pylori infection is associated with increased expression and production of C-X-C chemokines IL-8 and GROα, but not C-C chemokines. Although H pylori induces C-X-C chemokines both in the antrum and corpus, the association between chemokine expression and inflammation is less in the corpus.
Acknowledgments
We thank Professor David Y Graham, Baylor College of Medicine, for his many helpful discussions and assistance with the manuscript. Part of this work was presented at the Annual Meeting of the American Gastroenterological Association, Washington DC, May 1997, and appeared in abstract form in Gastroenterology1997;112:A336.