Article Text

Abstract
Background—bcl-2and bax belong to thebcl-2-related gene family, which marks a new class of genes that influence apoptosis. Thebcl-2 oncogene acts as a broad antiapoptotic factor and extends both normal and tumour cell survival. In contrast, the bax gene is a promoter of apoptosis.
Aims—To analyse the expression of bcl-2 andbax in pancreatic cancer and correlate the results with clinical parameters.
Patients—Pancreatic cancer tissue samples were obtained from 28 female and 32 male patients (median age 63, range 43–79 years) having surgery for pancreatic cancer. Normal pancreatic tissues obtained from 18 previously healthy organ donors served as controls.
Methods—The levels ofbcl-2 and baxmRNA expression were analysed by northern blot and the exact site of mRNA transcription was determined by in situ hybridisation. The presence of the corresponding proteins was determined by immunohistochemistry.
Results—Northern blot analysis indicated that, in comparison with the normal pancreas,bcl-2 mRNA was overexpressed in 30% andbax mRNA in 61% of the pancreatic cancer samples. Concomitant overexpression ofbcl-2 and bax was present in 26% of the cancer samples. Pancreatic adenocarcinomas exhibited 3.7-fold and 5.4-fold increases (p<0.001) inbcl-2 and baxmRNA levels respectively. In situ hybridisation showed that bothbcl-2 and baxmRNA were expressed in the cancer cells. Immunohistochemical analysis showed positive Bcl-2 and Bax immunostaining in 28 and 83% of the cancer samples respectively. In multivariate analysis (Cox regression model), bax expression was found to be a strong indicator of survival (p<0.001). Patients whose tumours exhibited Bax immunostaining lived significantly longer (12 months) than those whose tumours were Bax negative (five months) (p<0.039). In contrast, no relation was found between Bcl-2 and survival time.
Conclusions—The data indicate that genes that are involved in the regulation of apoptosis are upregulated in human pancreatic cancer cells. Prolonged survival times in patients in whom apoptosis promoting factors are upregulated indicate that apoptotic pathways are of biological significance in pancreatic cancer.
- pancreatic cancer
- apoptosis
- immunohistochemistry
- northern blot analysis
- in situ hybridisation
Statistics from Altmetric.com
In western industrialised countries, pancreatic cancer is at present the fourth or fifth leading cause of cancer death.1 This high death rate is due to the difficulty of achieving early diagnosis, rapid tumour progression, and unresponsiveness to chemotherapy, radiotherapy, immunotherapy, and antihormonal treatment.2 3All these parameters together result in low resectability rates after diagnosis, early recurrence after resection, and an overall poor survival rate.1
Recent molecular studies have identified a variety of growth factors and growth factor receptors which are overexpressed in human pancreatic cancer in vivo and in vitro.4-8 Furthermore, mutations of oncogenes (K-ras) and tumour suppressor genes (p53) are often present in pancreatic cancers.8-10It is believed that the combination of overexpression of growth promoting factors and the alteration or loss of inhibiting pathways may give pancreatic cancer cells a major growth advantage which clinically results in rapid tumour progression, formation of metastases, and subsequently the early death of the patient. In this study, we analysed for the first time the potential role of apoptosis in pancreatic cancer to evaluate whether alterations in this physiological pathway of cell elimination and tissue homoeostasis may be a further mechanism giving pancreatic cancer cells a survival advantage.
The bcl-2 gene family consists of several members, including bcl-2,bcl-xL, mcl-1, and bax. They function as blockers or promoters of apoptosis. Bcl-2, which functions as an antiapoptotic factor, is located on chromosome 18q21, which consists of three exons separated by large introns.11 Its antiapoptotic functions extend cell survival in both normal and tumour cells by inhibiting different cell death mechanisms, such as those induced by ionising radiation and anticancer drugs.12 In addition, transgenic mice overexpressing bcl-2 develop tumours, underlining the gene’s potential importance in tumour development and progression.13 14 In contrast with the apoptosis inhibitor bcl-2, thebax gene is a promoter of apoptosis. The protein encoded by the bax gene shares 21% amino acid sequence homology withbcl-2—mainly within two highly conserved regions called the bcl-2 homology 1 and 2.
Recent studies indicate that Bcl-2 and Bax form homodimers and/or heterodimers.15 The ratio between Bcl-2 and Bax determines cell survival or death following an apoptotic signal. An excess of Bcl-2 homodimers inhibits apoptosis, whereas an excess of Bax homodimers promotes cell death.15 16 Enhanced expression of bcl-2 has been reported in several lymphoid and non-lymphoid tumours.17 In patients with lymphomas, leukaemia, and prostate cancers, the presence of Bcl-2 in the cancer cells is associated with poorer therapy response, shorter disease free survival, and shorter overall survival.18-21In contrast, bax expression is reduced in breast cancers,22 and reducedbax expression is associated with shorter survival times in women with metastatic disease.21 These observations suggest that upregulation and downregulation of apoptosis-blocking or -stimulating genes have some influence on cancer cell viability and subsequently tumour progression. In this study therefore we analysed the expression and distribution ofbcl-2 and bax in human pancreatic cancers and correlated the molecular and immunohistochemical results with clinicopathological parameters of the patients to determine the significance of apoptosis in this malignancy.
Patients and methods
Normal human pancreatic tissue samples were obtained from 18 previously healthy individuals (five women and 13 men; median age 33, range 8–55 years) through an organ donor programme in which no recipient was available for pancreatic transplantation. Pancreatic cancer tissues were obtained from 60 patients (28 female, 32 male) having surgery for pancreatic cancer. The median age of the pancreatic cancer patients was 63 (range 43–79) years. A partial duodenopancreatectomy (Whipple resection) was carried out in 51 patients and a distal pancreatectomy in nine. According to the tumour, node, metastases (TNM) classification of the International Union Against Cancer,23 there were three patients with stage I, 19 with stage II, 36 with stage III, and two with stage IV disease.
Freshly removed pancreatic tissue samples were immediately fixed in Bouin or paraformaldehyde solution for 12–24 hours and paraffin embedded for routine immunohistochemical and in situ hybridisation analysis. In addition, tissues destined for RNA extraction were frozen in the operating room in liquid nitrogen immediately on surgical removal and maintained at −80°C until use. All the studies were approved by the ethics committee of the University of Bern, and by the Human Subjects Committee, University of California, Irvine, California, USA.
northern blot analysis
Total RNA was extracted by the guanidinium isothiocyanate method, fractionated on 1.2% agarose/1.8 mol/l formaldehyde gels, and stained with ethidium bromide for verification of RNA integrity and loading equivalence.27 The RNA was electrotransferred to Nylon membranes (GeneScreen; DuPont, Boston, Massachusetts, USA) and cross linked by UV irradiation. The filters were then prehybridised, hybridised, and washed under conditions appropriate for digoxigenin labelled antisense riboprobes (bcl-2 andbax) or the 32P labelled cDNA probe (7S) as previously described.27 28 In the case of the digoxigenin-labelled bcl-2 andbax cRNA probes, the filters were prehybridised and hybridised overnight at 68°C (bcl-2) or 65°C (bax) in a buffer containing 50% formamide, 2 × SSC (1 × SSC is 0.15 M NaCl/0.015 M sodium citrate buffer) (bcl-2) or 5 × SSC (bax), 2% blocking reagent (Boehringer Mannheim, Mannheim, Germany), 0.1%N-lauroylsarcosine, and 0.02% sodium dodecyl sulphate (SDS). The filters were then washed in 2 × SSC/0.1% SDS at room temperature, followed by three 15 minute washes at 68°C (bcl-2) or 65°C (bax) in 0.065 × SSC/0.065% SDS/35% formamide (bcl-2) or 0.1 × SSC/0.1% SDS (bax). The filters were then incubated in a blocking buffer (1% blocking reagent in 100 mmol/l maleic acid/150 mmol/l NaCl, pH 7.5) for 30 minutes, and in blocking buffer containing anti-digoxigenin alkaline phosphatase antibody (1:20 000; Boehringer Mannheim) for 30 minutes, washed three times with maleic acid buffer for 15 minutes, and incubated with 25 mmol/l CDP-Star (Boehringer Mannheim).28 The membranes were then exposed tox ray films.
In the case of the 7S cDNA probe, blots were prehybridised for eight hours at 42°C in a buffer which contained 50% formamide, 1% SDS, 0.75 mol/l NaCl, 5 mmol/l EDTA, 5 × Denhardt’s solution, 100 μg/ml salmon sperm DNA, 10% dextran sulphate, and 50 mmol/l phosphate buffer (pH 7.4). The hybridisation was carried out at 42°C for 18 hours by adding the 32P labelled cDNA probe (1 × 105cpm/ml). The blots were rinsed twice in 2 × SSC at room temperature and washed three times at 55°C in 0.2 × SSC/2% SDS under conditions appropriate for cDNA probes. Blots were then exposed at −80°C to Fuji x ray films with intensifying screens (Du Pont).
For statistical analysis of the northern blot results, the intensity of the radiographic bands was quantified by laser densitometry (Bio-Rad 620; Bio-Rad, Richmond, California, USA), as previously reported.5 28 The ratio between thebcl-2 and the corresponding 7S signal and the ratio between the bax and the corresponding 7S signal was calculated in each sample.
in situ hybridisation
In situ hybridisation was performed as previously reported.28 29 Briefly, using consecutive slides, one slide was incubated with the antisense probe and the other with the corresponding sense probe, and the slides were processed under the same conditions simultaneously. Tissue sections (2 μm) were deparaffinised, rehydrated with 1 × phosphate buffered saline (PBS) and incubated in 0.2 mol/l HCl for 20 minutes at room temperature. After the slides had been rinsed in 2 × SSC, the sections were treated with proteinase K (Boehringer Mannheim) at a concentration of 25 μg/ml for 15 minutes at 37°C. After post-fixation with 4% paraformaldehyde in PBS (five minutes) and washing in 2 × SSC, the samples were prehybridised at 60°C for at least two hours in solution containing 50% (v/v) formamide, 2 × SSC (bcl-2) or 4 × SSC (bax), 2 × Denhardt’s reagent, and 250 μg RNA/ml. Hybridisation was performed overnight at 60°C in solution containing 50% (v/v) formamide, 2 × SSC (bcl-2) or 4 × SSC (bax), 2 × Denhardt’s reagent, 500 μg RNA/ml, and 10% (w/v) dextran sulphate. The final concentrations of the digoxigenin labelled probes were about 0.8 ng/μl. After hybridisation, excess probe was removed by washing the slides in 2 × SSC and by RNase treatment: 100 U/ml RNase T1 (Boehringer Mannheim) and 0.2 μg/ml RNase (DNase free: Boehringer Mannheim) at 37°C for 30 minutes. After a wash in 2 × SSC for 20 minutes at 68°C (bcl-2) or 63°C (bax), and in 0.1 × SSC and 35% formamide at 68°C for 20 minutes (bcl-2) or 0.2 × SSC at 63°C for 20 minutes (bax), the samples were incubated with the anti-digoxigenin antibody conjugated with alkaline phosphatase (Boehringer Mannheim). For the colour reaction, 5-bromo-4-chloro-3-indolyl phosphate and nitro blue tetrazolium (Sigma, Buchs, Switzerland) were used.
probe synthesis for northern blot analysis and in situ hybridisation
Riboprobes of bcl-2 andbax were used for northern blot analysis and in situ hybridisation. A human bcl-2 cDNA fragment obtained from ATCC, Parklawn Drive, Rockville, Maryland, USA30 was used as the template for polymerase chain reaction (PCR), with a specific primer for a shorterbcl-2 fragment (sense: 5’-ACG ACA ACC GGG AGA TAG TGA T-3’; antisense: 5’-CAC ATG ACC CCA CCG AAC TCA A-3’). The resulting PCR product corresponding to nucleotides 1484 to 1932 of human bcl-2 was subcloned into the pCRII vector (Promega, Zürich, Switzerland) containing promoters for DNA dependent SP6 and T7 RNA polymerases. The authenticity of the subclonedbcl-2 fragment was confirmed by sequencing using the dye terminator method (ABI 373A; Perkin-Elmer, Rotkreuz, Switzerland).
The bax cRNA probe consisted of a 485 bp fragment of mouse bax cDNA, corresponding to nucleotides 1–485 (gift from Dr S J Korsmeyer, Howard Hughes Medical Institute, Washington University School of Medicine, St Louis, Missouri, USA), which shows high sequence homology with humanbax. The fragment was subcloned into the pGEM3Zf vector (Promega Biotechnology, Madison, Wisconsin, USA) carrying promoters for the DNA dependent SP6 and T7 RNA polymerases.
The 7S cDNA probe consisted of a 190 bpBamHI fragment of the mouse 7S cytoplasmic cDNA that cross hybridises with human 7S RNA.4 This probe was used to verify equivalent RNA loading in the northern blot experiments. The 7S cDNA probe was radiolabelled with [α-32P]dCTP (DuPont) using a random primer labelling system (Boehringer Mannheim) as previously reported.27 28
In the case of bcl-2 andbax, the cRNA probes were labelled with digoxigenin.29 After linearisation, the cDNAs were transcribed using the Ribomax System (Promega Biotechnology). The transcription resulted in digoxigenin labelled antisense riboprobes specific for the bcl-2 andbax mRNA. The corresponding sense probes were prepared in an analogous manner. For in situ hybridisation, the sense and antisense probes were shortened to a length of about 150 bases by the procedure of Cox et al 31 and stored in diethylpyrocarbonate treated water at −70°C until further use.
immunohistochemistry
Immunohistochemical analysis was performed with the streptavidin-phosphatase technique as previously reported. After deparaffinisation and hydration, the slides (2 μm) were washed in TBS buffer (10 mmol/l Tris/HCl, 0.85% NaCl, pH 7.4) containing 0.1% bovine serum albumin. In the case of Bcl-2 immunostaining, the sections were treated by immersion in 10 mM citrate buffer (10 mM sodium citrate, pH 6.0) and exposed to microwave irradiation twice for five minutes with a cooling period of four minutes in between the sessions.24 After cooling down to room temperature, the slides were transferred into fresh TBS buffer. In the case of Bax immunostaining, microwave treatment of the slides was not performed. All slides were incubated with 10% normal goat serum at 23°C for 30 minutes to block unspecific binding before the addition of specific monoclonal Bcl-2 or polyclonal Bax antibodies (overnight, 4°C) (Santa Cruz Biotechnology Inc, Santa Cruz, California, USA). The anti-Bcl-2 antibodies were produced by immunising mice against amino acids 41–54 of the human Bcl-2 protein. The anti-Bax antibodies were raised by immunising rabbits against amino acids 11–30 mapping at the amino terminus of Bax protein (p21) of human origin. The specificity of the antibodies was confirmed by western blot analysis. There was no cross reactivity between these antibodies or with other members of thebcl-2 gene family. After a wash with TBS buffer, biotinylated goat anti-mouse (for Bcl-2) and biotinylated goat anti-rabbit (for Bax) IgG secondary antibodies (Kirkegaard & Perry Laboratories, Gaithersburg, Maryland, USA) were added at 23°C for 45 minutes. After a wash in TBS buffer and incubation with the streptavidin/phosphatase complex at 23°C for 30 minutes, the slides were incubated with phosphate ester of 6-bromo-2-hydroxy-3-naphthoic acid (Kirkegaard & Perry Laboratories) as the substrate as previously reported.25 The slides were counterstained with Mayer’s haematoxylin.
To ensure specificity of the immunostaining reactions, consecutive tissue sections were incubated either in the absence of the primary antibody or with a non-immunised IgG antibody. In both cases, no immunostaining was detectable.
Semiquantitative evaluation of the immunohistochemical results was performed by two independent observers blinded to patient status. The intensity of the immunoreaction and the number of immunoreactive cancer cells were scored as previously reported.26 A final score of ⩾0.8 was taken in the statistical analysis as positive, and a score <0.8 was taken as negative.
statistical analysis
The data are expressed as median and range. Significance was defined as p<0.05. Statistical analysis was carried out by the Department of Mathematics and Statistical Analysis, University of Bern, using the SYSTAT Intelligent Software (Evanston, Illinois, USA) and SAS statistical package (SAS for Windows 6.11; SAS Institute Inc, Cary, North Carolina, USA). For statistical analysis, the Cox regression analysis, χ2 test, Mann-Whitney U test, and log-rank test were used. Survival data were plotted by the method of Kaplan and Meier and compared by the log-rank test and by the Wilcoxon test.32
Results
northern blot analysis
Northern blot analysis using total pancreatic RNA was carried out to determine bcl-2 andbax mRNA expression in the normal and the cancerous pancreas. bcl-2 andbax mRNA expression were present at low levels in all normal pancreatic tissue samples (fig 1). In some normal pancreatic tissues the expression levels for both genes were very faint and were only visible on the original autoradiographs. In contrast, hybridisation signals of bcl-2 andbax mRNA were higher in pancreatic cancer samples compared with the signals in normal samples (fig 1). None of the normal or cancer samples showed any aberrantbcl-2 or bax mRNA transcripts. bcl-2 mRNA was overexpressed in 30% of the cancer samples and bax mRNA was overexpressed in 61% of the cancer samples compared with the normal controls. In 26% of the pancreatic cancer samples both genes were concomitantly overexpressed, and in 35% of the pancreatic cancer samples the expression levels of both genes were comparable with those of normal controls. Densitometric analysis of the northern blots indicated that the bax mRNA levels in all cancer samples were 5.4-fold higher (p<0.001) than in the normal samples. bcl-2 mRNA levels in all cancer samples were 3.7-fold higher (p<0.001) than in the normal pancreatic tissue samples. When densitometric analysis was performed only in pancreatic cancer samples with enhancedbcl-2 or bax mRNA expression, there was a 6.7-fold increase forbcl-2 and a 19.6-fold increase forbax (p<0.001). There was no correlation between the increase above normal betweenbcl-2 and bax(r = 0.12, not significant) in the cancer samples.

Northern blot analysis of bcl-2 and bax in the normal pancreas (Normal, n=5) and in pancreatic cancer (Cancer, n=8).
in situ hybridisation
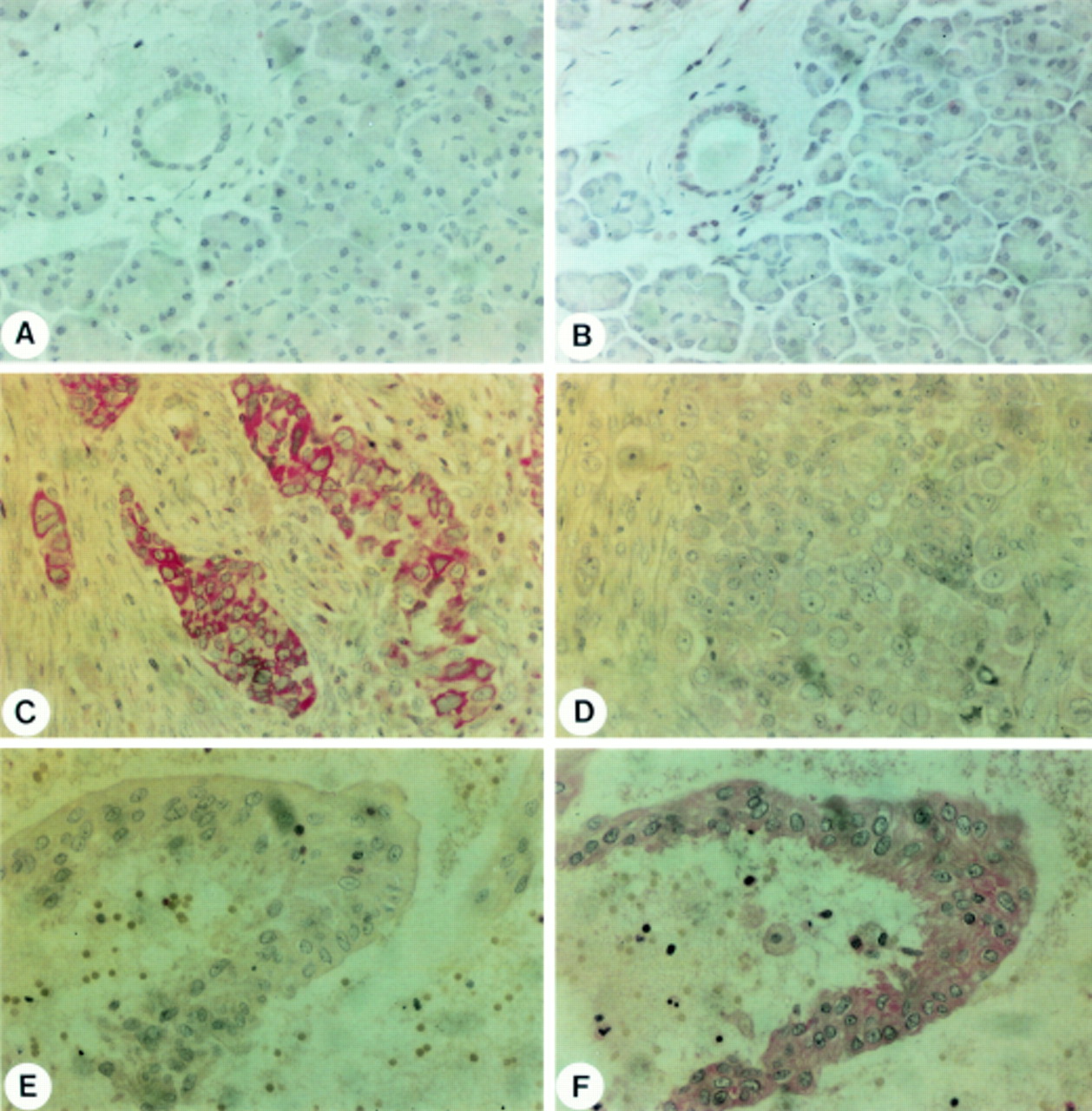
In situ hybridisation analysis was performed to determine the exact site of bcl-2 andbax mRNA synthesis in the normal and cancerous pancreas. In normal pancreatic tissue samples, no or very faint in situ hybridisation signals of bcl-2mRNA were observed (fig 2A). A similar low to undetectable expression pattern was observed for bax mRNA in the normal pancreas (fig 2B). bax mRNA was present only in a few acinar cells and in some ductal cells. In contrast, bcl-2 (fig 2C) andbax mRNA (fig 2D) signals were strongly present in the pancreatic cancer samples which exhibited enhanced levels on northern blot analysis, especially in the cancer cells forming duct-like structures. Some 25 and 56% of the pancreatic cancer samples exhibited moderate to strong bcl-2and bax mRNA staining respectively. Around 25% of the cancer samples exhibiting enhancedbcl-2 mRNA expression also exhibited enhanced bax mRNA expression on in situ hybridisation.

In situ hybridisation of bcl-2 (A, C) and bax (B, D) in the normal pancreas (A, B) and in pancreatic cancer (C-F). Panels C and D are consecutive pancreatic cancer tissue sections showing cancer cells with concomitant bcl-2 (C) and bax (D) mRNA expression. However, not all cancer cells exhibit bax or bcl-2 mRNA expression. Hybridisation of pancreatic cancer sections with the bcl-2 (E) and bax (F) sense probes failed to give a signal. Original magnification × 400.
In some areas with chronic pancreatitis-like lesions adjacent to the cancer mass, faint to moderate bcl-2 andbax mRNA staining was found in the remaining ductal and acinar cells. In contrast, there were nobcl-2 or bax mRNA signals in the connective tissue and in fibroblasts in areas adjacent to the cancer lesions or in the regions with chronic pancreatitis-like morphological changes (data not shown).
Control slides hybridised with the bcl-2(fig 2E) or the bax (fig 2F) sense probe did not show any specific in situ hybridisation signals.
immunohistochemistry
Immunohistochemical analysis was performed in normal and cancerous pancreatic tissue samples to localise the respective proteins. Weak Bax immunoreactivity was present in a few acinar cells and in many ductal cells of the normal human pancreas (fig 3B). Bcl-2 immunoreactivity was either absent or faintly present in the acinar or ductal cells in normal pancreatic tissue samples (fig 3A). Bcl-2 and Bax immunostaining were not found in the interlobular and periductal stroma in the normal pancreas. In contrast, moderate to intense Bcl-2 immunoreactivity (fig3C) was present in 17 of 60 (28%) of the cancer samples and Bax immunoreactivity (fig 3F) was present in 50 of 60 (83%) of the cancer samples. When Bcl-2 and Bax immunostaining were analysed in consecutive tissue sections, eight of 60 (13%) cancer samples were negative for both factors. However, in 15 of 60 (25%) cancers, moderate to strong Bcl-2 and Bax immunoreactivity was simultaneously present in the cancer cells. Two of 60 (3%) cancers exhibited Bcl-2 (fig 3C) but not Bax immunostaining (fig 3D), whereas 35 of 60 (58%) cancer samples exhibited Bax (fig 3F) but not Bcl-2 immunostaining (fig3E).

{kind=link}
{kind=link}
{kind=link}
Immunohistochemical analysis of Bcl-2 (A, C, E) and Bax (B, D, F) in the normal pancreas (A, B) and in pancreatic cancer (C–F). Panels C and D and E and F are consecutive pancreatic cancer tissue sections. Panels C and D show pancreatic cancer cells that exhibit Bcl-2 (C) immunoreactivity but no Bax (D) immunostaining. Panels E and F show pancreatic cancer cells that do not exhibit Bcl-2 (E) immunoreactivity but do exhibit Bax (F) immunostaining. Original magnification × 400.
association of immunohistochemical findings with clinicopathological parameters
To determine whether the presence of Bcl-2 or Bax immunostaining in pancreatic cancer cells is of clinical significance, the immunohistochemical data were correlated with patient data (sex, age) and clinical data (tumour stage and postoperative survival after tumour resection).
When the survival data were plotted by the method of Kaplan and Meier and statistically analysed, the presence of Bax immunostaining but not of Bcl-2 immunostaining in pancreatic cancer cells was associated with longer postoperative survival. Patients whose tumours exhibited no Bax immunoreactivity in the cancer cells lived for a median (range) five (three to nine) months, whereas those whose tumours exhibited Bax immunostaining in the cancer cells lived for 12 (three to 29) months (p = 0.0398) after surgery.
We found no relation between the presence of Bcl-2 immunoreactivity in pancreatic cancer cells and age, sex or tumour stage. There was also no relation between the presence of Bax immunoreactivity and these parameters. Moreover, with Cox regression analysis, the presence of Bax was found to be the only factor that could strongly predict the risk of dying from the tumour. Its influence on the prognosis was highly significant (p<0.001). The conditional risk ratio for Bax was 0.212, which suggests that during the period of postoperative survival, pancreatic cancer patients with Bax immunoreactivity in the tumour cells will have about a 5 times lower risk of dying than those without Bax immunoreactivity. Other parameters, like Bcl-2 immunostaining, sex, and age, had no significant influence on the risk of dying postoperatively in this analysis.
Discussion
In this study we analysed the concomitant expression ofbcl-2 and bax in normal and cancerous human pancreatic tissue samples. The genes belong to the same family of apoptotic genes. Although the factors are structurally similar, they exert opposite effects on apoptosis. Bcl-2 inhibits apoptosis and contributes to cell survival and the resistance of cells against damaging influences. In contrast, Bax, which is considered to be a central regulator of apoptosis, is a promoter of programmed cell death.
Northern blot analysis showed enhanced bcl-2and bax mRNA expression in 30 and 61% of the cancer samples respectively. By in situ hybridisation both mRNA moieties were predominantly expressed in the cancer cells and were expressed to a minor extent in areas adjacent to the tumour with chronic pancreatitis-like morphological changes. Bcl-2 and Bax immunostaining in the cancer cells was absent in 72 and 17% of the cancer samples respectively. Interestingly, the presence of the apoptosis-promoting bax gene product in the pancreatic cancer cells was associated with longer postoperative survival. In contrast, the presence or absence of Bcl-2 immunostaining in the pancreatic cancer samples had no influence on patient survival or other tumour parameters. These findings suggest that the presence of Bax in pancreatic cancer may activate apoptosis, which subsequently influences the viability of pancreatic cancer cells and thereby influences postoperative survival after tumour resection.
The exact role of Bcl-2 in human cancers is at present not clear and there is wide variability of Bcl-2 immunopositivity in different human malignancies. In gastric adenocarcinomas and invasive ductal breast carcinomas, about 75% of the tumour samples exhibit enhanced Bcl-2 immunoreactivity in the cancer cells33 34 in comparison with normal tissues. In contrast, in colonic carcinomas, the percentage of Bcl-2 immunoreactivity is much lower and only 5% of the tumour samples show Bcl-2 immunostaining in the cancer cells.35
The presence of Bcl-2 in cancer cells seems to influence clinical outcome differently depending on the underlying malignancy. In non-small cell lung carcinomas, breast carcinoma, and follicular thyroid carcinomas,36-38 the presence of Bcl-2 immunoreactivity in the cancer cells is associated with poor prognosis. However, in axillary node-positive breast cancer patients, multivariate statistical analysis of clinical and molecular data has demonstrated that the presence of bcl-2 expression is independently associated with longer disease free and overall survival.34 Furthermore, in colonic carcinomas both positive and negative influences of Bcl-2 on survival and prognosis have been reported.35 39 These findings indicate that the apoptosis-inhibiting gene bcl-2 is expressed in various frequencies in different cancers, and a general pattern of activation or inactivation of these genes in malignant tumours cannot be defined. Therefore the function of apoptotic genes in different human cancers must be evaluated individually.
In our analysis of 60 pancreatic cancers, increasedbcl-2 immunoreactivity was present in 28% of the cancer samples. However, the presence ofbcl-2 in the cancer cells was not associated with the prognosis and survival time of the patients after tumour resection. These findings suggest that the Bcl-2-mediated antiapoptotic pathway is of minor importance for the growth behaviour and viability of pancreatic cancer cells in vivo. In contrast, we found a significant correlation between the presence of Bax in the cancer cells and patient prognosis: increased Bax immunostaining was present in 83% of the cancer samples, and its presence was associated with significantly longer survival. Inasmuch as Bax stimulates apoptosis, its presence in the pancreatic cancer cells may attenuate their survival potential, resulting in a better clinical outcome. Conversely, in breast tumours, baxexpression was reduced compared with normal controls, and reducedbax expression correlated with shorter overall survival in women with metastatic breast carcinomas.21 Although most pancreatic cancers exhibited enhanced bax expression—whereas in breast cancers bax expression in the cancer cells was below that of normal cells—in both malignancies the presence of Bax in the cancer cells is associated with longer survival compared with tumours in which Bax is absent. These findings are in agreement with the overall concept that Bax is a promoter of apoptosis and that cancer cells with high levels of Bax may be more vulnerable to environmental factors than Bax negative cancer cells.
Bcl-2- and Bax-mediated apoptosis is dependent on a balance between homodimer and/or heterodimer formation of these proteins. In most previous studies on the influence of these proteins on cancer cell behaviour, the factors were analysed separately. However, the concomitant presence of, and the balance between, Bcl-2 and Bax is important for defining whether a cell is more likely to be eliminated by apoptotic mechanisms or to acquire resistance to cell death. Taking this into consideration, we evaluated the simultaneous presence of Bcl-2 and Bax in our study. However, we found no relation between simultaneous bcl-2 andbax mRNA expression levels, or immunohistochemical staining with respect to patient characteristics, tumour parameters, or survival. It is possible that the gene products of other bcl-2 family members also possess the ability to form heterodimers with Bax in a competitive manner.40 Therefore Bcl-2 may not be the only factor to influence Bax mediated apoptotic effects in an additive or diminishing fashion. Therefore additional apoptotic pathways need to be studied to understand cancer cell survival more clearly.
In summary, our present findings suggest that Bax, but not Bcl-2, influences the biological growth behaviour of pancreatic cancers in vivo. In contrast with previously reported molecular alterations in growth factors, growth factor receptors, proto-oncogenes, and tumour suppressor genes, the enhanced presence of Bax is a strong parameter which seems to contribute to a better prognosis in pancreatic cancer patients.
Acknowledgments
This work was supported by a research grant from the Sandoz Stiftung zur Förderung der Medizinischen Forschung.