Article Text

Abstract
Background and aims: A number of Helicobacter pylori outer membrane proteins (OMPs) undergo phase variations. This study examined the relation between OMP phase variations and clinical outcome.
Methods: Expression of H pylori BabA, BabB, SabA, and OipA proteins was determined by immunoblot. Multiple regression analysis was performed to determine the relation among OMP expression, clinical outcome, and mucosal histology.
Results:H pylori were cultured from 200 patients (80 with gastritis, 80 with duodenal ulcer (DU), and 40 with gastric cancer). The most reliable results were obtained using cultures from single colonies of low passage number. Stability of expression with passage varied with OipA > BabA > BabB > SabA. OipA positive status was significantly associated with the presence of DU and gastric cancer, high H pylori density, and severe neutrophil infiltration. SabA positive status was associated with gastric cancer, intestinal metaplasia, and corpus atrophy, and negatively associated with DU and neutrophil infiltration. The Sydney system underestimated the prevalence of intestinal metaplasia/atrophy compared with systems using proximal and distal corpus biopsies. SabA expression dramatically decreased following exposure of H pylori to pH 5.0 for two hours.
Conclusions: SabA expression frequently switched on or off, suggesting that SabA expression can rapidly respond to changing conditions in the stomach or in different regions of the stomach. SabA positive status was inversely related to the ability of the stomach to secrete acid, suggesting that its expression may be regulated by changes in acid secretion and/or in antigens expressed by the atrophic mucosa.
- BHI, brain heart infusion
- bp, base pair
- DU, duodenal ulcer
- FBS, fetal bovine serum
- IM, intestinal metaplasia
- Le, Lewis antigen
- OMP, outer membrane protein
- OR, odds ratio
- PCR, polymerase chain reaction
- Helicobacter pylori
- outer membrane proteins
- SabA
- OipA
- BabA
Statistics from Altmetric.com
- BHI, brain heart infusion
- bp, base pair
- DU, duodenal ulcer
- FBS, fetal bovine serum
- IM, intestinal metaplasia
- Le, Lewis antigen
- OMP, outer membrane protein
- OR, odds ratio
- PCR, polymerase chain reaction
Adherence of Helicobacter pylori to the gastric mucosa is believed to play an important role in the inflammatory response to the organism.1 At least 32 H pylori outer membrane proteins (OMPs) have been identified, many of which are involved in bacterial adherence.2 A number of the genes encoding these OMPs undergo phase variations in the 5′ region such that not all strains produce functional proteins. Five genes encoding OMPs (oipA, sabA, sabB, babB, and hopZ) are thought to be regulated by the slipped strand mispairing based on the number of CT dinucleotide repeats in the 5′ region of the genes such that when inframe the gene is “on” and when out of frame it is “off”.2,3
Fucosylated ABO blood group antigens and the related sialyl-Lewis x and sialyl-Lewis a antigens (sLex and sLea) have been identified as functional receptors for H pylori adherence.4,5 ABO antigens are recognised by the blood group antigen binding adhesin BabA6 whereas sLex/a is recognised by the sialic acid binding adhesin SabA.5 The outer inflammatory protein OipA also has been suggested to have a role in binding but the cognate receptor structure remains unknown.7
Recent studies have suggested H pylori expressing BabA and OipA are associated with more severe mucosal cellular inflammation and an increased risk of clinical outcomes such as peptic ulcer disease and gastric cancer.8,9,10,11,12,13,14 However, not all studies agree.14–16 Most of the available studies determined OMP functional status indirectly by examining genomic variations using mismatch polymerase chain reaction (PCR) (for the babA2 gene)8,9,10,14,15 and PCR based sequencing for detecting the number of CT repeats (for the oipA, sabA, and hopZ genes).12,13,15,16 However, the high degree of genetic diversity among H pylori complicates interpretation of PCR based methods and may result in an underestimation of the frequency of the functional status of the gene (that is, silent genes may be scored as expressed and chimeric H pylori OMPs as non-expressed).17–19 For example, it has been demonstrated that many Leb non-binding strains possess silent babA gene sequences which may become activated by recombination into the babB locus forming chimeric babB/A genes.17 Recombination events between two babA genes may also result in a change in ABO antigen receptor specificity.18 In the present study, we used immunoblotting to directly evaluate the protein expression status of OipA, BabA, BabB, and SabA in relation to clinical outcomes and to gastric histology.
MATERIALS AND METHODS
Bacterial strains
We examined H pylori isolates cultured from patients from South America (Colombia) and the USA. Presentations included simple H pylori gastritis, duodenal ulcer (DU) disease, and advanced non-cardiac gastric adenocarcinoma. DU was defined endoscopically; patients with ulcer scars were excluded as were those with DU and gastric ulcer. Patients who used non-steroidal anti-inflammatory drugs were also excluded. Gastritis was defined as histological gastritis only. None of the strains had been published in our prior studies of H pylori virulence factors. Additional selection criteria included at least eight gastric mucosal biopsies for Colombian patients and more than five for US patients. We excluded gastric cancer cases from the USA due to the small numbers available.
Gastric histology
Gastric mucosal biopsy specimens were stained with Genta20 or El-Zimaity triple stains.21 Sections were examined by the pathologist who was blinded to the patient’s clinical diagnosis or the characteristics of the H pylori strain. We examined up to five biopsy specimens from the antrum (A1 from the antral lesser curvature within 2 cm of the pylorus, A2 from the mid lesser curvature of the antrum, A3 from the gastric angle, A4 from the antral greater curvature within 2 cm of the pylorus, and A5 from the mid greater curvature of the antrum) and up to six specimens from the corpus (B1 from the greater curvature proximal to the antral-corpus border, B2 from the greater curvature at least 4 cm proximal to the antral-corpus border, B3 from the proximal greater curvature, B4 from the proximal lesser curvature, B5 from the lesser curvature within 5 cm of the anglus, and B6 from the greater curvature in the mid corpus). Each specimen was scored for H pylori density, neutrophil infiltration, intestinal metaplasia (IM), and atrophy. All variables were scored using a visual analogue scale graded from 0 (absent/normal) to 5 (maximal intensity), as described previously.22 Scores in each site were averaged both in the antrum and corpus.
H pylori phenotyping
Antral specimens (from A4 or A5) were obtained for isolation of H pylori using standard culture methods.23,24 In order to minimise the risk of phase variations, H pylori samples were obtained by expansion of a single colony. To reduce the likelihood of a change in OMP phase status during in vitro passage, we used H pylori with in vitro passage numbers less than 4 (including the expansion from a single colony). We compared the results using cultures grown from multiple colonies with those grown from single colonies. We also evaluated the effect of multiple in vitro passages (10 and 20 times) and compared the results using cultures obtained from antral and corporal biopsy specimens (B6). Unless stated otherwise, experiments were done with low passage number H pylori grown from single colonies obtained from antral biopsies.
We used actively replicating organisms obtained after 20–24 hours of incubation which corresponds to the logarithmic growth phase; immunoblotting was performed using standard methods. Anti-BabA antiserum (AK277),25 anti-BabB antiserum (AK276),25 anti-OipA antiserum (AK282),26 and anti-SabA antiserum (AK278)27 were used as the first antibody at a 1:5000 dilution. Horseradish peroxidase conjugated protein A (1:3000) (Bio-Rad Lab, Hercules, California, USA) was used as the second antibody. Proteins were detected by ECL chemiluminescent substrate detection reagents (Amersham Life Science, Arlington Heights, Illinois, USA). The accuracy of the immunoblot for antiserum against BabA, BabB, OipA, and SabA was confirmed previously.25–27 As previous studies reported that oipA status was closely related to cagA status12, we also examined CagA status using commercial anti-CagA antibody (Austral Biologicals, San Ramon, California, USA).
Acid exposure
After overnight culture, H pylori were suspended in brain heart infusion (BHI) broth (pH 5.0) supplemented with 10% fetal bovine serum (FBS). The pH of the media was adjusted using concentrated hydrochloric acid prior to addition of H pylori. After two hours of acid exposure, H pylori were harvested and used for immunoblotting. Controls consisted of cultures grown in pH 7.0 BHI broth for two hours.
Data analysis
For independent univariate analysis, Fisher’s exact test, the Mann-Whitney rank sum test and t test were used, depending on the data set of concern. A multiple logistic regression analysis was performed to determine which putative virulence factor(s) was the most discriminating for clinical outcome. Bacterial factors, as well as age and sex, were used as explanatory variables. Multiple linear regression analyses were used for histological data as the data had a normal distribution. In the analyses, bacterial factors, sex, age, and clinical outcomes were the explanatory variables and mutually adjusted associations with the criterion variables were calculated. A p value of less than 0.05 was accepted as statistically significant. Calculations were carried out using statistical software “HALBAU” (Gendai-sugaku-sha, Kyoto, Japan).
RESULTS
Strains from 200 patients were examined, including 120 from Colombia (40 with gastritis, 40 with DU, and 40 with gastric cancer) and 80 from the USA (40 with gastritis and 40 with DU). Colombian patients comprised 71 men and 49 women (mean age 52.5 years). US patients comprised 68 men and 12 women (mean age 54.2 years). Mean age of gastric cancer patients (57.7 years) was significantly higher than that of DU (51.3 years) or gastritis patients (48.5 years) (p<0.01 for each). There were no significant differences in sex or age distribution among the different clinical outcomes.
H pylori OMPs status and clinical outcomes
Independent univariate analysis using Fisher’s exact test showed that OipA status was significantly associated with DU (table 1). OipA was present more often in strains from DU patients (88%) than in those from gastritis patients (61%) (p<0.001), confirming previous studies evaluating OipA status by genotyping.11 SabA positive status was less prevalent in strains from DU patients (44%) compared with those from gastritis patients (66%) (p<0.01) or from gastric cancer patients (70%) (p<0.05).
Univariate analysis of relationship between Helicobacter pylori genotyping and clinical outcome
Multiple logistic regression analysis among OMPs showed that only the presence of OipA and SabA were related to clinical outcome (table 2). Only OipA status was an independent determinant predictor of gastric cancer versus gastritis (adjusted odds ratio (OR) 4.8 (95% confidence interval (CI) 1.4–16.8)) and DU versus gastritis (OR 4.0 (95% CI 1.6–10.2)). Only SabA status was a predictor of gastric cancer versus DU (OR 2.8 (95% CI 1.2–6.7)). The presence of SabA was associated with a reduced risk of DU compared with gastritis although the differences did not reach statistical significance (OR 0.52 (95% CI 0.27–1.0)).
Multiple logistic regression analysis: Helicobacter pylori factors associated with clinical presentation
When we added CagA status as an explanatory variable for the analyses, calculations could not be converged as OipA and CagA status were generally identical. In fact, 188 of 200 (94%) samples showed identical CagA/OipA phenotype (correlation coefficient 0.83) (p<0.001) (table 3). OipA, CagA, and BabA protein status were significantly correlated with one another in agreement with previous studies at the genomic level.6,8,12,15 When we added CagA status and eliminated OipA status, CagA status was a determinant predictor of gastric cancer versus gastritis (OR 6.5 (95% CI 1.4–29.3)); however, it was not a predictor of DU versus gastritis (OR 2.4 (95% CI 0.9–6.2)).
Relationship among the Helicobacter pylori genotypes
H pylori OMPs status and gastric mucosal histology
Histological analyses were performed in gastritis and DU cases. Independent univariate analysis using the Mann-Whitney rank sum test indicated that BabA, SabA, OipA, and CagA status were closely related to changes in gastric histology (table 4). As BabA, OipA, and CagA status were not independent of one another (as described above), we chose to look at all predictor variables at the same time and performed a backward stepwise multiple linear regression analysis. For this analysis, we used H pylori factors, including CagA status as well as country and clinical outcome, as explanatory variables. OipA positive status was significantly associated with high H pylori density and grade of neutrophil infiltration (table 5). In the analyses, the partial regression coefficient 0.71 of OipA for H pylori density was interpreted as showing that the H pylori density score with OipA positive strains would be expected to be 0.71 points greater than with OipA negative strains. SabA positive status was closely associated with decreased neutrophil infiltration (antrum and corpus) and with severe IM and gastric atrophy (antrum) (table 5). SabA positive status also remained in the final model for corporal IM and gastric atrophy although the differences did not reach statistical significance. BabA positive status was also significantly associated with severe IM and gastric atrophy in the antrum (table 5). BabB positive status remained in the final model for decreased neutrophil infiltration; however, the differences did not reach statistical significance. As scores for corporal IM were relatively low in most cases, we also categorised IM as positive or negative (IM at no sites). In agreement with multiple linear regression analysis, only SabA status was independently related to corporal IM (adjusted OR 4.6 (95% CI 1.5–14.2)).
Univariate analysis of relationship between Helicobacter pylori genotyping and histology
Final model using multiple linear regression analysis: Helicobacter pylori factors associated with histology
In the present study, we examined 11 biopsy specimens (five from the antrum and six from the corpus). Many investigators have used the updated Sydney system which recommends five biopsy specimens for histological analyses (two from the antrum, two from the mid to proximal corpus, and one from the gastric angle). We therefore re-evaluated our samples according to that system (using biopsy sites A1, A3, A4, B4, and B6). Results using five biopsy specimens and 11 biopsy specimens differed with respect to corporal IM and atrophy (table 5). Using only two corporal biopsy specimens, no factors remained in the final model for corporal IM and different factors (CagA) remained in the final model for corporal atrophy.
Recently, the Houston system for scoring corporal atrophy was devised based on the fact that corpus atrophy advances from the antrum into the corpus, extending up the lesser curve followed by the greater curve.28,29 The system consisted of a four point scale ranging from 0 = none to 3 = severe, with 0 = atrophy at no corpus site, 1 = mild or atrophy only at B4, 2 = moderate or atrophy at B2 and B4, and 3 = severe atrophy, including B6. Using this system, SabA status remained in the final model for atrophy and was statistically significant (partial regression coefficient 0.63 (SEM 0.25); p = 0.015), confirming the observation that the Sydney system underestimates the frequency and severity of atrophy/IM.
Immunoblotting for H pylori factors from single colonies and multiple colonies
The above analyses evaluated H pylori factors obtained from single colonies. To confirm the accuracy of the immunoblotting using single colonies, we examined 20 samples with variable production patterns of OMPs (table 6). We re-cultured frozen biopsy specimens from these 20 samples and multiple colonies were harvested en masse. The results for OipA and CagA were identical between multiple colonies and a single colony. In contrast, half of the strains’ negative results for SabA from a single colony (4/8 samples) yielded positive results from multiple colonies (table 6). These differences in phenotypic designation (negative becoming positive results) were also observed with BabA (13%) and BabB (25%). To confirm our results, we chose 20 colonies from each strain where negative immunoblot results for SabA using single colonies produced positive results using multiple colonies (four samples). Immunoblot showed that mixed SabA positive and negative strains were common, where 10–15% of samples from a new single colony yielded positive results despite SabA status having been characterised as negative using with the original single colony (data not shown). Thus one should probably describe the isolates as being of the predominant phenotype. In a preliminary experiment we found that if the samples contained more than approximately 5% OMP positive strains, immunoblot yielded positive results and this held true for all four OMPs evaluated (data not shown). We also showed that the results of single colony experiments usually identify the predominant phenotype (data not shown).
Changing of outer membrane protein (OMP) status by multiple colonies and multiple in vitro passages
Effects of in vitro passages on H pylori phenotype
Previous studies reported that OMPs status may switch over time in vivo and during in vitro laboratory passages.17–19,30 To evaluate the effect of in vitro passages, we selected 20 strains from those used in the above experiments (table 6). We used the original strains grown from a single colony (passage number less than 4) and transferred them 10 and 20 times in vitro. OipA and CagA status did not change during 20 passages. In contrast, during 20 passages, BabA and BabB status changed in 10% and SabA status changed in 25% (table 6). Both changes (negative becoming positive results and positive becoming negative results) were observed in SabA and BabB whereas only the change from negative to positive was observed in BabA. Changes were rare during 10 passages with the only change being one SabA which changed to negative, suggesting that a passage number of 10 or less may best represent in vivo conditions.
Immunoblotting using H pylori from antral and corpus biopsies
We compared results of immunoblotting of strains from the antrum and corpus of the same patient. We selected 20 strains used in the above experiments (table 6) and cultured H pylori from their corresponding corpus biopsy specimens. H pylori proteins were extracted from bacteria grown from single colonies. The results for antral and corpus specimens were identical (data not shown).
Effect of low acid condition on H pylori phenotype
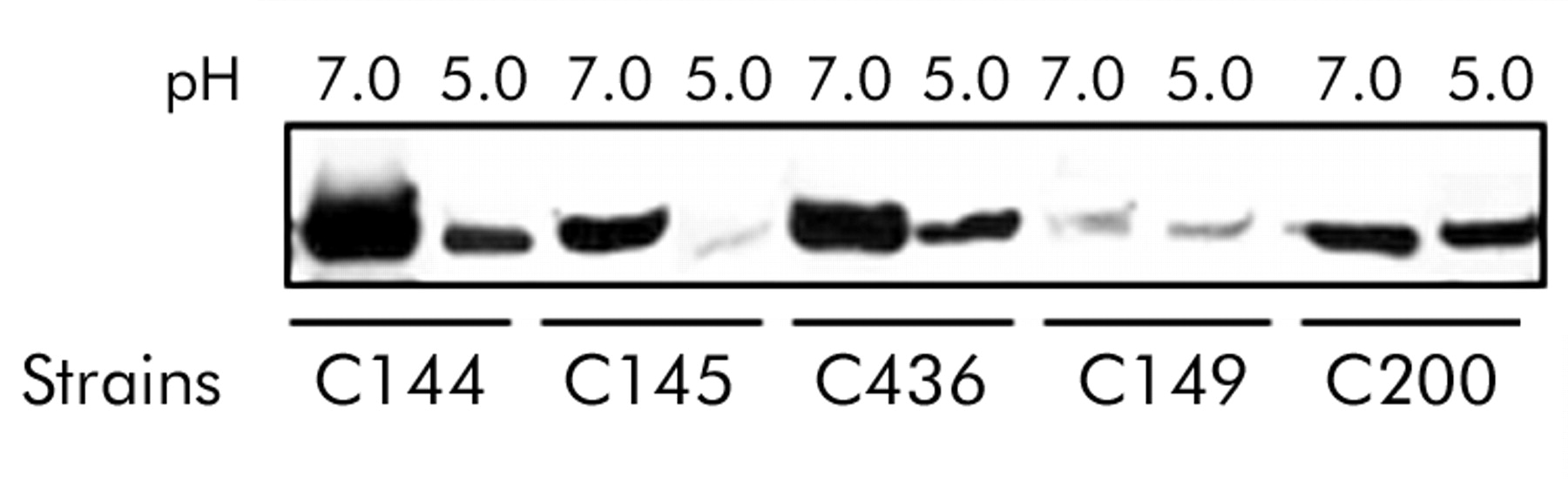
The above in vivo results showing that SabA was associated with gastric cancer and the severity of gastric atrophy/IM suggested that SabA expression may be regulated by changes in the intragastric milieu, such in pH. To test this hypothesis, we compared cultures grown in high acid conditions (pH 5.0) with those grown at pH 7.0. We selected 10 strains (Nos 1–10 in table 6). Our preliminary experiments showed that virtually all H pylori survived the two hours of acid exposure in BHI broth at pH 5.0 (data not shown). Two hours of acid exposure did not result in any change in status for CagA or the OMPs, with the exception of expression of SabA which was dramatically decreased in three of the five stains (fig 1). We therefore further examined these three strains following exposure to pH 5.5 in BHI broth supplemented with 10% FBS for 20–24 hours, with shaking at 200 rpm. We used pH 5.5 as apparent logarithmic growth phase of H pylori was not observed during 24 hours of culture in broth at pH 5.0. SabA status changed from positive to negative in one strain (C436) and expression of SabA decreased in the other two strains (data not shown).

{kind=link}
Effect of low pH on SabA expression by immunoblot analysis. Helicobacter pylori was exposed to pH 7.0 and pH 5.0 broth for two hours. Equal amounts of protein were separated on a 10% polyacrylamide gel, transferred to nitrocellulose membranes, and probed with an anti-SabA antiserum (AK278). SabA expression was decreased in acidic conditions in strains C144, C145, and C436.
DISCUSSION
There has been increasing interest in the role of H pylori OMPs, especially OipA and BabA, in gastroduodenal disease.8,9,10,11,12,13,14 Our studies confirmed previous observations with OipA and BabA and report new findings in relation to SabA and clinical outcome and gastric histology. OipA and CagA appear to be stably expressed. In contrast, BabA and SabA were prone to undergo phase variations. The stomach presents a variety of different microenvironments such that the ability to switch “on” and “off” various adhesins and surface proteins may offer survival advantages to the bacteria. For example, deep in the pits in the antrum the bacteria would be exposed to different populations of cells compared with those on the luminal surface, as well as a much lower fluctuations in pH. The fact that SabA expression was increased in strains obtained from patients with gastric cancer and prevalence was reduced among those with DU suggested that switching may be related to intragastric pH. We showed that SabA expression was dramatically decreased at pH 5. This result is in agreement with a previous study using microarray that expression levels of sabA mRNA decreased in acidic conditions.31 It would be interesting to examine SabA status among patients undergoing long term acid suppression as well as its possible role in proton pump inhibitor associated increased corpus inflammation.32
SabA expression was also increased in patients with intestinal metaplasia and gastric atrophy, suggesting that there may be a relation between SabA and the antigens/receptors expressed by this altered epithelium. In contrast, BabA expression was associated with both high and low acid conditions (for example, gastric cancer and DU),8,9,10,11,12,13,14 suggesting that phase variations of BabA are unlikely to be influenced by intragastric pH and/or the change in gastric mucosal cell types.
SabA has been suggested to be a sialic acid binding adhesin.5 Other studies have suggested that SabA is responsible for binding of H pylori bacterial cells to gangliosides.33 During persistent infection and chronic inflammation (gastritis), H pylori triggers an alteration in the patterns of glycosylation in the gastric mucosa, including upregulation of the inflammation associated sLex antigens. It has been suggested that SabA performs selectin mimicry by binding the sialyl-(di)-Lex/a glycosphingolipids promoting membrane attachment and apposition. It is possible (or suggested) that at sites of vigorous local inflammatory response, H pylori undergo phase variations and switch off the sLex binding capacity, allowing non-functional SabA to escape intimate contact with sialylated lymphocytes or other defensive cells. Thus SabA should be involved in adaptation of bacterial adherence properties to inflammation pressure.
We also directly examined the relationship between expression of SabA protein and clinical outcome. Previous studies indirectly evaluated SabA status using PCR to assess whether the gene was likely “on” or “off”. Lehours and colleagues15 reported that combination of the sabA “on” status with hopZ “off” status and iceA1 allele was related to an increased risk of mucosa associated lymphoid tissue lymphoma. In contract, de Jonge and colleagues16 evaluated SabA status by the number of CT repeats in a small study (for example, nine gastric cancers) and reported that sabA “on” status had no relation to clinical outcome. In that study, they considered all strains with 7±3 CT repeats as gene “on” status. However, recent reports have shown that the sabA gene can be “on” or “off” depending on the nucleotide sequence after the CT repeats. The presence of sabA “off” status strains has been confirmed among strains with seven CT repeats,15 suggesting that conclusions based on PCR based methods for SabA status should be interpreted with caution. This caution also applies to BabA in that distinguishing babA2 from babA1 based on 10 bp deletion of the signal peptide sequence may provide miscategorisations as regulation by the 10 bp deletion appears to be specific for strain CCUG178756 as no other strain containing two babA genes called 1 and 2 has been reported. Recent data showed that BabA expression was also regulated at the level of transcription and by formation of the chimeric babA-babB gene.17–19 In the present study, BabA status was changeable during in vitro passages. Further studies are necessary to determine the mechanism (for example, differences in the poly(A) tract in the babA promoter, existence of recombination events between babA and babB).
Overall, because of the technical difficulties in evaluating OMPs status by PCR, we suggest that currently the most reliable method is to confirm protein expression by immunoblot. That said, there are important methodological cautions, including the use of single colonies, low in vitro passage number, and confirmation by evaluating H pylori obtained from different regions of the stomach. These cautions are based on the finding of a relatively high frequency of coexistence of H pylori expressing and not expressing the factor of interest. The frequency of mixed expression was highest with SabA, less with BabB and BabA, and very rare with OipA. Our data suggest that the single colony method usually represents the predominant phenotype whereas the multiple colonies method is biased towards the presence of positive strains in the sample. While study of the role of different proportions of H pylori expressing putative virulence factors in relation to outcome are theoretically interesting, current technical limitations restrict analyses to studies based on the predominant phenotype which cannot be reliably determined using results from multiple colonies. We also found that the problem of phase shifting during in vitro passage could be minimised using low in vitro passage number. Finally, we showed that correct categorisation of the status of the gastric mucosa in relation to the presence of IM and atrophy of the corpus was dependent on the number and sites of biopsies obtained. The updated Sydney system recommends two proximal or mid corpus biopsies and may significantly underestimate the presence of intestinal metaplasia/atrophy extending upwards from the antral-corpus border but not yet reaching the sites recommended by the Sydney system. Our results suggest that discovery of the predominant strain being SabA positive might suggest the presence of hypochlorhydria or achlorhydria and prompt a search for corpus atrophy.
Acknowledgments
Olabisi Ojo was supported by the UNESCO BAC short term fellowship and the Macarthur Foundation/University of Ibadan Staff Training Grant. This material is based on work supported in part by the National Institutes of Health grants R01 DK62813 (to YY), by the Office of Research and Development Medical Research Service Department of Veterans Affairs (to DYG), by Deutsche Forschungsgemeinschaft DFG (project OD 21/1-1) (to SO), and by Public Health Service grant DK56338 which funds the Texas Gulf Coast Digestive Diseases Center.
REFERENCES
Footnotes
-
Published online first 1 December 2005
-
↵* Current address: Department of Biochemistry, College of Medicine, University of Ibadan, Ibadan, Nigeria
-
Conflict of interest: None declared.
Linked Articles
- Digest