Article Text

Abstract
Introduction: Impaired activity of natural killer (NK) cells has been proposed as a mechanism contributing to viral persistence in hepatitis C virus (HCV) infection. As the function of NK cells is primarily regulated by NK cell receptors (NKR), we analysed whether decreased NK cell function in hepatitis C may be related to dysregulated NKR expression.
Patients and methods: Expression of NK cell was analysed by flow cytometry on lymphocytes from HCV(+) subjects (n = 30), patients who became HCV(−) after antiviral therapy (n = 10), healthy individuals (n = 10), and hepatitis B virus (HBV) infected patients (n = 9). Cytolytic function of lymphocytes was studied in a redirected lysis assay and in a standard 51chromium release cytotoxicity assay, respectively.
Results: In patients with chronic hepatitis C, we found a significantly reduced proportion of NKp46 and NKp30 expressing NK cells compared with healthy and HBV infected subjects. Low expression of natural cytotoxicity receptor (NCR) was also confirmed in in vitro activated NK cell populations derived from HCV patients compared with uninfected donors. In contrast, patients who cleared HCV under antiviral therapy showed normal expression of NKp44, NKp30, and NKp46. Reduced NCR expression in chronic hepatitis C was associated with a parallel decrease in NCR mediated target cell killing. Furthermore, we found a significantly increased proportion of NKG2A expressing NK cells and CD8+ T cells in HCV positive patients, resulting in a reduced cytolytic activity against cells incubated with the HLA-E stabilising peptide HCV core35–44.
Conclusion: The present study indicates that defective expression of NKR represents a novel mechanism contributing to impaired function of NK cells and CD8+ T cells in chronic hepatitis C.
- HCV, hepatitis C virus
- HBV, hepatitis B virus
- NK, natural killer
- NKR, NK cell receptors
- NCR, natural cytotoxicity receptors
- HIV, immunodeficiency virus
- MHC, major histocompatibility complex class
- TCR, T cell receptor
- rIL-2, recombinant interleukin 2
- TGF-β, transforming growth factor β
- hepatitis C virus
- natural killer cell receptors
- HLA-E
- natural killer cells
- CD8+ T cells
Statistics from Altmetric.com
- HCV, hepatitis C virus
- HBV, hepatitis B virus
- NK, natural killer
- NKR, NK cell receptors
- NCR, natural cytotoxicity receptors
- HIV, immunodeficiency virus
- MHC, major histocompatibility complex class
- TCR, T cell receptor
- rIL-2, recombinant interleukin 2
- TGF-β, transforming growth factor β
Impaired activity of CD8+ T lymphocytes1–3 as well as dysfunction of natural killer (NK) cells4–8 has been suggested to be critically involved in the establishment of chronic hepatitis C virus (HCV) infection. However, the mechanisms leading to suppressed function of NK cells and CD8+ T cells are only poorly understood.
The function of NK cells is regulated by multiple cell surface NK cell receptors (NKR)9 which can be divided into activating and inhibitory NKR. Furthermore, recent reports have demonstrated that NKR are also importantly involved in the regulation of immune functions elicited by CD8+ T cells.10–12 Thus altered expression of NKR is likely to result in altered functions of NK cells and CD8+ T cells. With respect to hepatitis C, Khakoo et al showed that genes encoding the inhibitory NK cell receptor KIR2DL3 and its human leucocyte antigen C group 1 (HLA-C1) ligand directly influence resolution of HCV infection, supporting the importance of NKR for the course of hepatitis C.13
The diversity of NK cell receptors observed in different individuals is in part genetically determined.14 On the other hand, microbial infections may modify the NKR repertoire as increased proportions of CD8+ T cells bearing inhibitory NKG2A have been observed in mice infected by different viruses12,15,16 as well as in human immunodeficiency virus (HIV),17 and HCV infected patients.18
Inhibitory NKG2A as well as activating NKG2C belong to the C-type lectin NKR and both specifically recognise the non-classical major histocompatibility complex class (MHC) I molecule HLA-E, which presents peptides commonly derived from the signal sequences of other MHC class I molecules.19–21 Of note, we recently demonstrated enhanced expression of HLA-E on a variety of intrahepatic cells in chronic hepatitis C. Furthermore, we showed that the hepatitis C virus polyprotein gives rise to at least one peptide (HCV core aa35–44; YLLPRRGPRL) that stabilised surface expression of HLA-E and thus inhibited NK cell function.22
Activation of NK cells is primarily mediated by the natural cytotoxicity receptors (NCR) NKp46, NKp30, and NKp44.23 The cell surface ligand(s) of these NCR is still unknown.24 However, NK cell mediated cytolytic activity has been shown to be strictly correlated with NCR density,25,26 and reduced expression of NKR in HIV RNA(+) patients has been shown to be associated with reduced lytic function of NK cells.27
Apart from modulating NK cell function, NKR are also importantly involved in tuning T cell responses. Activation of T cells is primarily induced by signals generated by direct interaction of the T cell receptor (TCR) with antigen, presented via MHC I molecules on antigen presenting cells.28 In addition to positive signals via the TCR, the magnitude of CD8+ T cell activation also depends on signals mediated by NKR expressed on T cells,10–12,28 including CD94/NKG2A/C and NKG2D, another member of the C-type lectin family.10 There is increasing evidence that inhibitory NKRs such as NKG2A are importantly involved in regulation of CTL functionality.10–12 For instance, Speiser et al demonstrated that the weak cytolytic activity of in vitro expanded NKG2A positive melanoma specific CTL can be restored by blocking CD94/NKG2A with a specific antibody.10,11,29
Here we determined expression of activating (NKp30, NKp46, NKp44, NKG2C, and NKG2D) and inhibitory (NKG2A) NKR on NK cells and CD8+ T cells of patients chronically infected with HCV and correlated changes in expression pattern to altered cytolytic functions of NK and CD8+ T cells.
MATERIAL AND METHODS
Patients
Forty Caucasian individuals, all from the Bonn area in Germany, were enrolled into this study. Thirty patients had chronic hepatitis C and 10 subjects had cleared the virus after combined antiviral therapy with pegylated interferon α and ribavirin. None of these patients had histological signs of liver cirrhosis. As control groups, we also studied 10 healthy HCV RNA negative donors and nine hepatitis B virus (HBV) infected patients (table 1).
Patient characteristics
NK cells were immunomagnetically purified from peripheral blood mononuclear cells according to the manufacturer’s protocol (Miltenyi, Bergisch Gladbach, Germany). Serum samples were collected and stored at −80°C until use.
The study conformed with the guidelines of the Declaration of Helsinki, as approved by our ethics committee.
FACS analysis
The following antibodies were used for FACS analysis: anti-CD56-FITC, anti-CD8-APC, and anti-CD3-PercP were obtained from BD (BD Bioscience, Heidelberg, Germany). Anti-NKG2A-Pe, anti-NKG2C-Pe, and anti-NKG2D-Pe were purchased by R&D (R&D Systems, Wiesbaden, Germany), and anti-NKp30-PE, anti-NKp44-Pe, and anti-NKp46-Pe were obtained from Beckman Coulter (Heidelberg, Germany). After incubation of the cells with 10 μl of antibody for 20 minutes and washing in phosphate buffered saline, samples were analysed on a FACScalibur using CellQuest software.
Flow cytometric analysis of cells in the liver specimens
Liver biopsy specimens for flow cytometric analysis of intrahepatic cells were obtained from liver biopsies of five HCV RNA(+) patients and four control subjects with hepatic diseases not related to HCV using 1.5 mm diameter disposable biopsy needles. Fresh liver samples were washed twice in fresh medium and shaken gently to avoid blood contamination. Liver specimens were disrupted mechanically into small fragments in RPMI 1640 medium with 10% fetal calf serum FCS using a forceps and scalpel. Then the fragments were homogenised on a cell strainer (BD Labware, Heidelberg, Germany). The resulting cell suspension was washed and resuspended in RPMI 1640 medium. Intrahepatic cells were then directly analysed by flow cytometry.
Cell culture conditions
After immunomagnetic purification, NK cells were cultured in the presence of recombinant interleukin 2 (rIL-2) at 100 U/ml (Proleukin; ChironCorp., Emeryville, California, USA), and 1.5 ng/ml of PHA (Gibco Ltd, Paisley, UK) to obtain activated polyclonal NK cell populations.25,27 The culture medium was RPMI 1640 supplemented with 10% fetal calf serum, L-glutamine (2 mM/l), and penicillin, and NK populations were analysed two weeks after purification/activation.27
Redirected cytotoxicity assay
Fc dependent redirected cytotoxicity was studied as described previously.30 In brief, NCR mediated lytic activity was studied after a four hour incubation of isolated NK cells with 51Cr labelled FcγR+ P815 cells at various E:T ratios. The function of NCR was analysed by adding soluble anti-NKp30 (clone 210847), NKp44 (clone 253415), or anti-NKp46 (clone 195314), all obtained from R&D Systems.. The increase in 51Cr release over baseline (irrelevant monoclonal antibody) reflects the ability of a given monoclonal antibody to trigger the lytic machinery of the cells. As a negative control we used an IgG specific monoclonal antibody (clone A85-1; BD Biosciences).
Accordingly, purified CD8+ T lymphocytes were incubated with anti-CD3 and soluble anti-NKG2A to asses the effect of NKG2A stimulation on TCR mediated lytic activity of CTL. To analyse the functional role of CD94, we added anti-CD94 instead of NKG2A. As a negative control, we used an IgG specific monoclonal antibody (clone A85-1). In further control experiments anti-CD94 or anti-NKG2A were added without anti-CD3.
51Cr release was measured with a β counter. Maximum release was determined by addition of Triton X. Specific lysis was calculated as (release − spontaneous release) : (maximum release − spontaneous release) ×100.
Cytotoxicity assay
A 51chromium release assay was performed as described previously31 using K-562 and HLA-E transfected K-562 HLA-E cells as target (T) cells and purified NK cell populations as effector (E) cells. Prior to performing the assay, 1×104 target cells were incubated overnight at 26°C in the absence or presence of peptide HCV core aa35–44 (100 μM) to load peptides onto HLA-E molecules.22
To identify the type of NK cell receptor involved, we also preincubated NKL cells with either anti-CD94 or anti-NKG2A antibody at 10 μg/ml before performing the cytotoxicity assay. As a negative control, NK cells were incubated with an irrelevant mouse IgG specific antibody (clone A85-1; BD Biosciences). E:T ratios were varied as indicated for the different experiments. Specific lysis was calculated as (release − spontaneous release):(maximum release – spontaneous release) ×100.
Statistical analysis
Statistical analyses (Student and Mann-Whitney U tests, respectively) were performed with the SPSS 9.0 (SPSS, Chicago, Illinois, USA) statistical package. Results were considered significant at the two sided p level of 0.05.
RESULTS
Circulating CD3(−)CD56(+) NK cells as a percentage of total circulating lymphocytes did not differ significantly between the four analysed groups (fig 1A). However, flow cytometric analysis of C-type lectin receptor surface expression on NK cells (fig 1B) revealed an enhanced proportion of NKG2A expressing NK cells in HCV infected patients (52.3 (5.2)%) in comparison with healthy controls (16.7 (3)%; p<0.001). Unlike patients with hepatitis C, HBV infected patients showed NKG2A expression comparable with that of healthy subjects (fig 1C, left diagram). No significant differences were found between healthy controls and patients with chronic viral hepatitis regarding expression of NKG2C and NKG2D (fig 1C).

Expression and function of C-type lectin receptors in chronic hepatitis C virus (HCV) infection. (A) Circulating CD3(−)CD56(+) natural killer (NK) cells as a percentage of total circulating lymphocytes, as determined by flow cytometry. (B) Representative examples of flow cytometric analysis of C-type lectin expression on CD56(+) NK cells. (C) Surface expression of the C-type lectin receptors NKG2A, NKG2C, and NKG2D was analysed by flow cytometry of circulating CD3−/CD56+ NK cells from patients with chronic hepatitis C (n = 28), HCV infected patients who became HCV RNA negative after antiviral therapy (n = 10), HBV infected subjects (n = 9), and healthy control individuals (n = 10). Results are mean (SEM). (D) Lysis of HLA-E transfected K-562 cells pulsed with or without the HLA-E stabilising peptide HCV core aa35–44 (lower histograms) by NK cells from a healthy HCV uninfected and HCV RNA positive subject. Representative data of three experiments. *p<0.05, HCV(+) versus HCV(−).
Next, we addressed the functional consequences of upregulated expression of the inhibitory NK cell receptor NKG2A, which specifically interacts with HLA-E. To this end, we isolated NK cells from peripheral blood of HCV infected patients and healthy volunteers. Purified NK cells were tested in a 51chromium release assay using HLA-E transfected K-562 (K-562 B5) cells as target cells. K-562 B5 cells were pulsed with peptide HCV core aa35–44, which has been shown to stabilise surface expression of HLA-E.22 NK cells from HCV infected patients showed markedly reduced lysis of peptide pulsed B5 cells compared with NK cells from healthy individuals. Blocking with anti-NKG2A confirmed that this reduced lysis of HLA-E expressing cells was due to interactions of NKG2A with HLA-E (fig 1D, and data not shown).
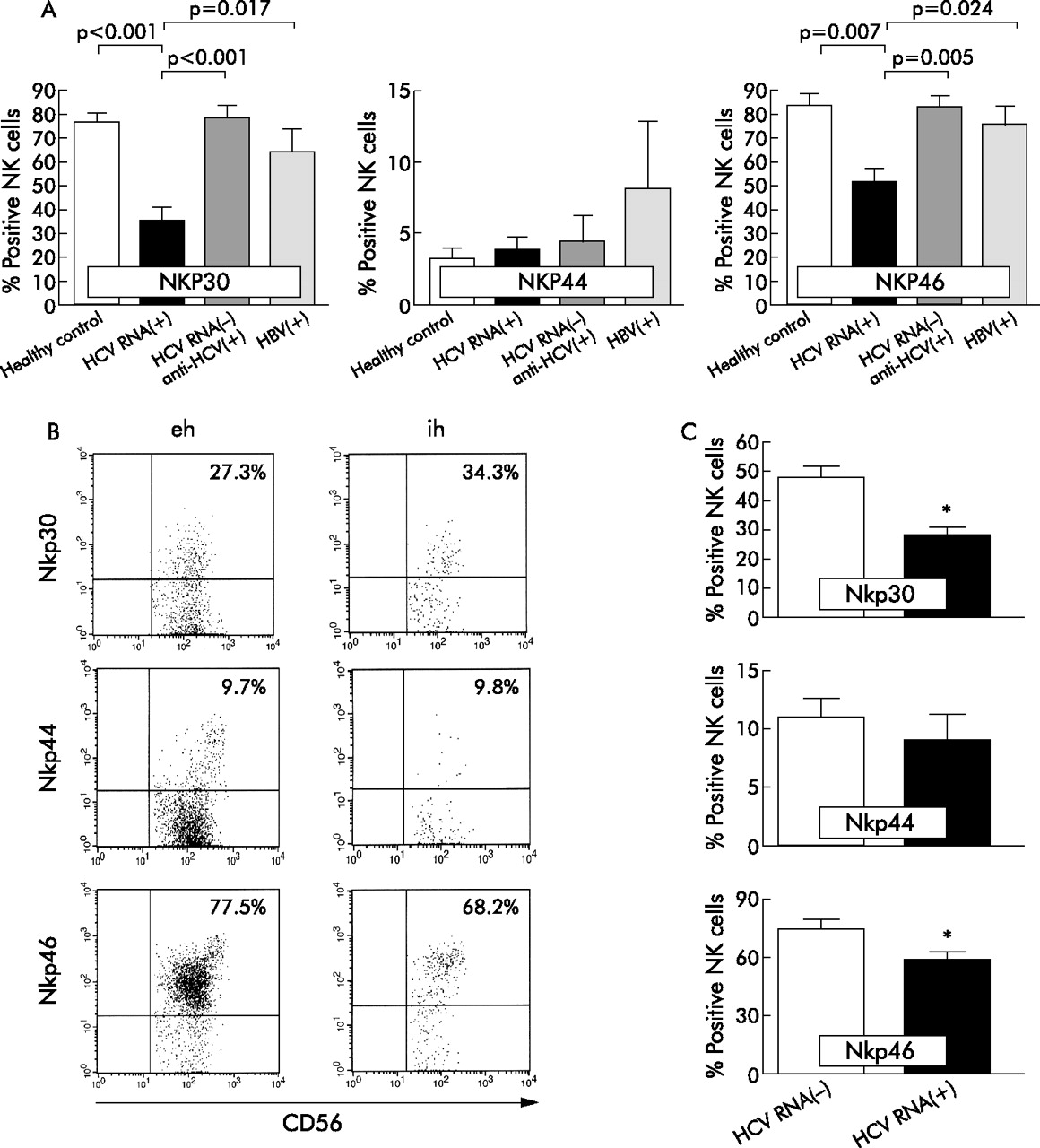
Regarding expression of NCR, we found a significantly lower percentage of NKp30 expressing NK cells in HCV infected than in healthy controls (mean 35.9 (SEM 4.6)% v 77.5 (3.3)%; p<0.001) and HBV patients (60.9 (8.9)%; p = 0.017) (fig 2A). Staining of NK cells from HCV patients with the NKp46 specific antibody was also significantly reduced compared with staining of cells from HCV RNA negative subjects (53.7 (5.6)% v 84.4 (4.6)%; p = 0.015) and patients infected with HBV (77.6 (6.9)%; p = 0.024) (fig 2A). In contrast, no significant differences were found regarding percentage of NKp44 expressing CD56+ NK cells (figs 2A).

Natural cytotoxicity receptor (NCR) expression on natural killer (NK) cells in chronic hepatitis C. (A) Expression of NCR was analysed by flow cytometry on circulating CD3-CD56 NK cells from patients with chronic hepatitis C (n = 28), hepatitis C virus (HCV) infected patients who became HCV RNA negative under antiviral therapy (n = 10), hepatitis B virus (HBV) infected subjects (n = 9), and healthy control individuals (n = 10). Results are mean (SEM). (B) NCR expression on intra- and extrahepatic CD56+NK cells from a representative HCV(+) patient. (C) Percentage of NCR expressing intrahepatic CD56+ NK cells from patients with chronic hepatitis C (n = 5) and four patients with non-HCV related hepatic diseases. Values are mean (SEM). *p<0.05, HCV(+) v HCV(−).
To address the issue of whether reduced expression of NKp30 and NKp46 is associated with replicative HCV infection, we studied surface expression of NCR in patients who had cleared the virus after antiviral therapy with pegylated interferon α and ribavirin. Interestingly, we found that aviraemic patients showed a significantly higher proportion of NKp30 positive (79.5 (4.5)%; p<0.001) and NKp46 positive (84 (4.5)%; p = 0.005) NK cells than HCV RNA positive patients, comparable with expression levels in healthy controls (fig 2A). There was a positive correlation between proportions of NKp30 positive and NKp46 positive NK cells in chronic hepatitis C (fig 3B). However, no positive correlation was found between expression levels of any of the NCR and serum HCV loads (fig 3A).

Expression of natural cytotoxicity receptors and viral load. (A) Correlation between expression levels of NKp30, NKp44, and NKp46, and hepatitis C virus (HCV) serum load (copies/ml) in chronically HCV infected patients. (B) Correlation between proportion of NKp30 and NKp46 expressing natural killer (NK) cells in chronic hepatitis C.
Recently, we demonstrated that expression of NKG2A was upregulated on intrahepatic NK cells in HCV infection and exceeded expression of NKG2C.22 However, nothing is known of expression of NCR on liver infiltrating NK cells. Therefore, we stained intrahepatic CD3(−)CD56(+) lymphocytes for expression of NKp30, NKp44, and NKp46, respectively. We found a comparable proportion of NCR positive NK cells in peripheral blood and in the livers in chronic hepatitis C, although some patients showed higher expression of NKp30 on intrahepatic compared with circulating NK cells (fig 2B). Nevertheless, we observed a significantly lower percentage of NK cells expressing NKp30 and NKp46 in livers from HCV infected patients than in livers from patients with non-HCV associated hepatic diseases (fig 2C).
Next we analysed whether reduced expression of NCR on purified peripheral blood NK cells from HCV RNA positive patients could also be confirmed after NK cells were activated and cultured in vitro. For this purpose, we cultured purified NK cell populations from peripheral blood of HCV infected patients or from healthy uninfected controls in the presence of rIL-2, as described previously.27
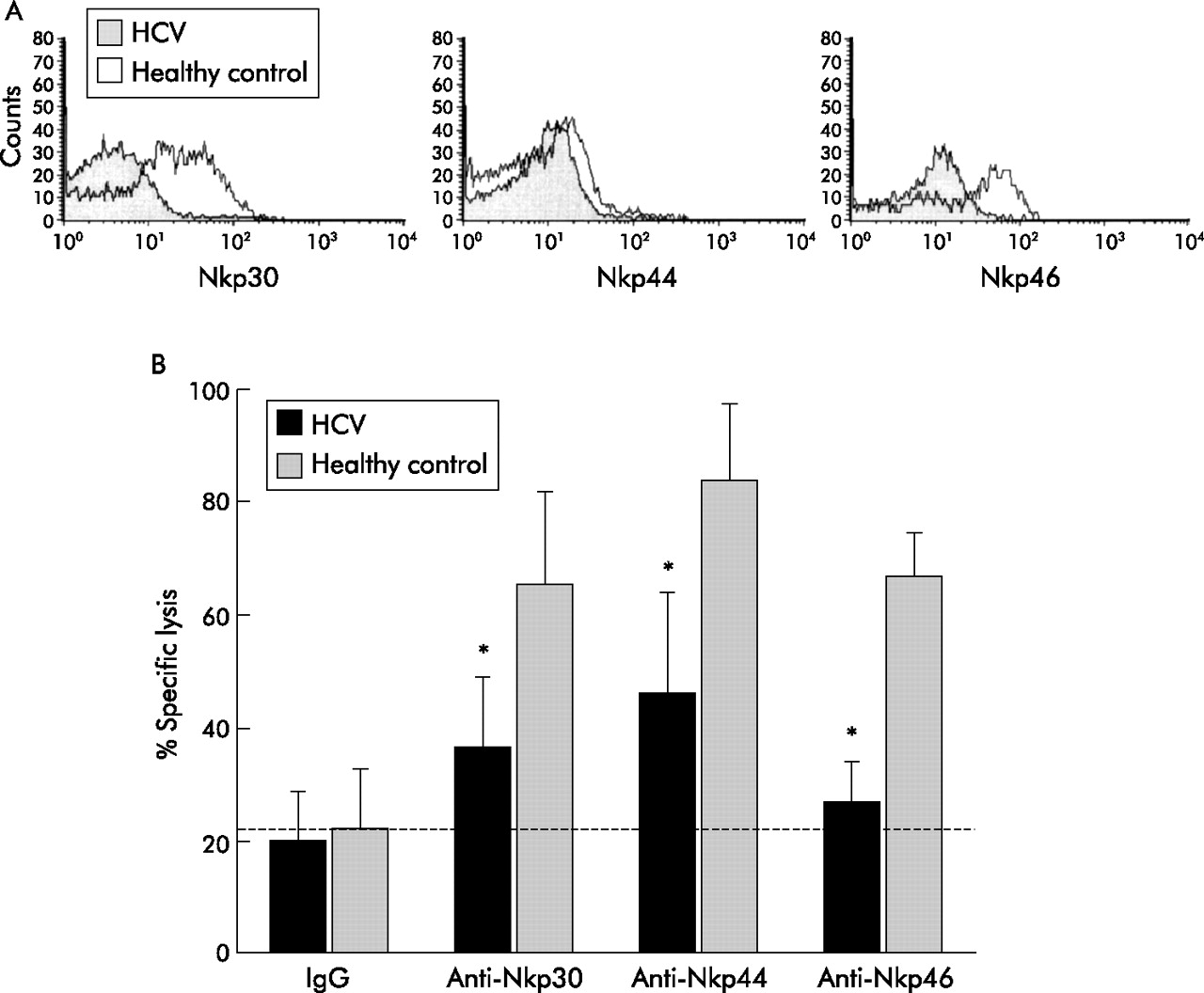
Flow cytometric analysis of NKp30 and NKp46 revealed that these receptors were still expressed at reduced levels in the in vitro activated NK cell cultures derived from HCV RNA positive patients. In addition, expression of NKp44, which is usually upregulated by in vitro culture of NK cells, was also reduced on NK cells isolated from HCV infected patients compared with NK cells from healthy subjects (fig 4A).

Function and expression of natural cytotoxicity receptors (NCR) on in vitro activated natural killer (NK) cells. (A) Single colour flow cytometric analysis of expression of NKp46, NKp30, and NKp44 NCR on activated NK cell populations, cultured in vitro in the presence of interleukin 2, from a representative HCV infected patient and from a representative healthy hepatitis C virus (HCV) RNA(−) donor. Fluorescence indicates reactivity to each given monoclonal antibody. Results are expressed as logarithm of fluorescence intensity (arbitrary units, horizontal axis) versus cell counts (vertical axis). The figure shows data from one of five sets of experiments. (B) Redirected killing assays of in vitro activated NK cell populations derived from HCV infected patients (n = 5) and uninfected donors (n = 3) against FcR+ P815 cells. NK cell effectors were assayed in a four hour 51Cr release assay against P815 cells at 5:1 effector:target cell ratios in the presence of anti-NKp46, anti-NKp30, or anti-NKp44 monoclonal antibody or the irrelevant IgG specific control antibody, respectively. The increase in 51Cr release over baseline (irrelevant monoclonal antibody) reflects the ability of a given monoclonal antibody to trigger the lytic function of NK cells. *p<0.05.
To check whether reduced expression of NCR also has a functional correlate, we analysed the lytic function of NK cells from HCV infected patients in a redirected killing assay. In these experiments, the cytolytic activity of NK cell populations was assessed against the FcR+ P815 murine target cell line, in the absence or presence of monoclonal antibody directed at NKp30, NKp44, and NKp46, as described before.27
We found a significant overall decrease in median NCR mediated lysis of P815 targets in the redirected killing assay. Median lytic activity of NK cell populations derived from HCV RNA positive patients in the presence of NKp46, NKp30, or NKp44 specific monoclonal antibody represented approximately 50% of what was observed for the NK cell populations obtained from healthy controls (fig 4B).
Finally, we studied expression and function of NKR on CD8+ T cells in chronic hepatitis C. No differences were found regarding numbers of circulating CD8+ T lymphocytes (fig 5A) between the groups. Furthermore, we did not find expression of NCR on CD8+ T cells (data not shown), which is in line with previous reports.32–34 However, analysing expression of C-type lectin receptors (fig 5B), we found a significantly higher percentage of CD8+ T lymphocytes expressing the inhibitory NK cell receptor CD94/NKG2A in HCV RNA positive patients (mean 15.4 (SEM 2.9)%) than in healthy individuals (5.6 (0.5)%; p = 0.003) or in patients with chronic hepatitis B (5.42 (1.9)%; p = 0.002). Patients who cleared HCV RNA after combined therapy with interferon and ribavirin displayed NKG2A expression comparable with that of healthy individuals (6.7 (1.9)%) (fig 5C). Expression of NKG2A was closely related to HCV serum levels (r = 0.543, p = 0.045; data not shown), indicating that NKG2A expression was related to HCV replication.

Expression of C-type lectin receptors on CD8(+) T lymphocytes. (A) Circulating CD3(+)CD8(+) T cells as a percentage of total circulating lymphocytes, as determined by flow cytometry. (B) Representative examples of flow cytometric analysis of C-type lectin expression on CD3(+)CD8(+) T lymphocytes. (C) CD3(+)CD8(+) T lymphocytes from healthy individuals (n = 10), hepatitis C virus (HCV) RNA positive patients (n = 20), patients who became HCV RNA negative after antiviral therapy (n = 10), and hepatitis B virus (HBV) infected subjects (n = 9) were analysed by flow cytometry for expression of NKG2A (left), NKG2C (middle), and NKG2D (right), respectively. Results are mean (SEM).
In contrast, no statistically significant differences were found for expression of NKG2C (5.3 (1.2)% v 6.0 (0.8)%; p>0.05) and NKG2D (94.4 (0.5)% v 83.0 (6.5)%; p>0.05) between HCV RNA positive and healthy subjects. However, NKG2C was significantly lower on CD8 T lymphocytes from patients who became HCV negative after antiviral therapy (2.9 (0.5)%) compared with lymphocytes from both HCV RNA(+) patients (p = 0.037) and healthy controls (p = 0.001). No differences were found regarding expression of NKG2D (fig 5C).
Multicolour analysis of CD3(+)CD8(+) T revealed that most of the CD3+ CD8+ NKG2A+ lymphocytes were CCR7−, CD28−, CD45RA−, and CD45RO+, indicating a memory phenotype (data not shown).
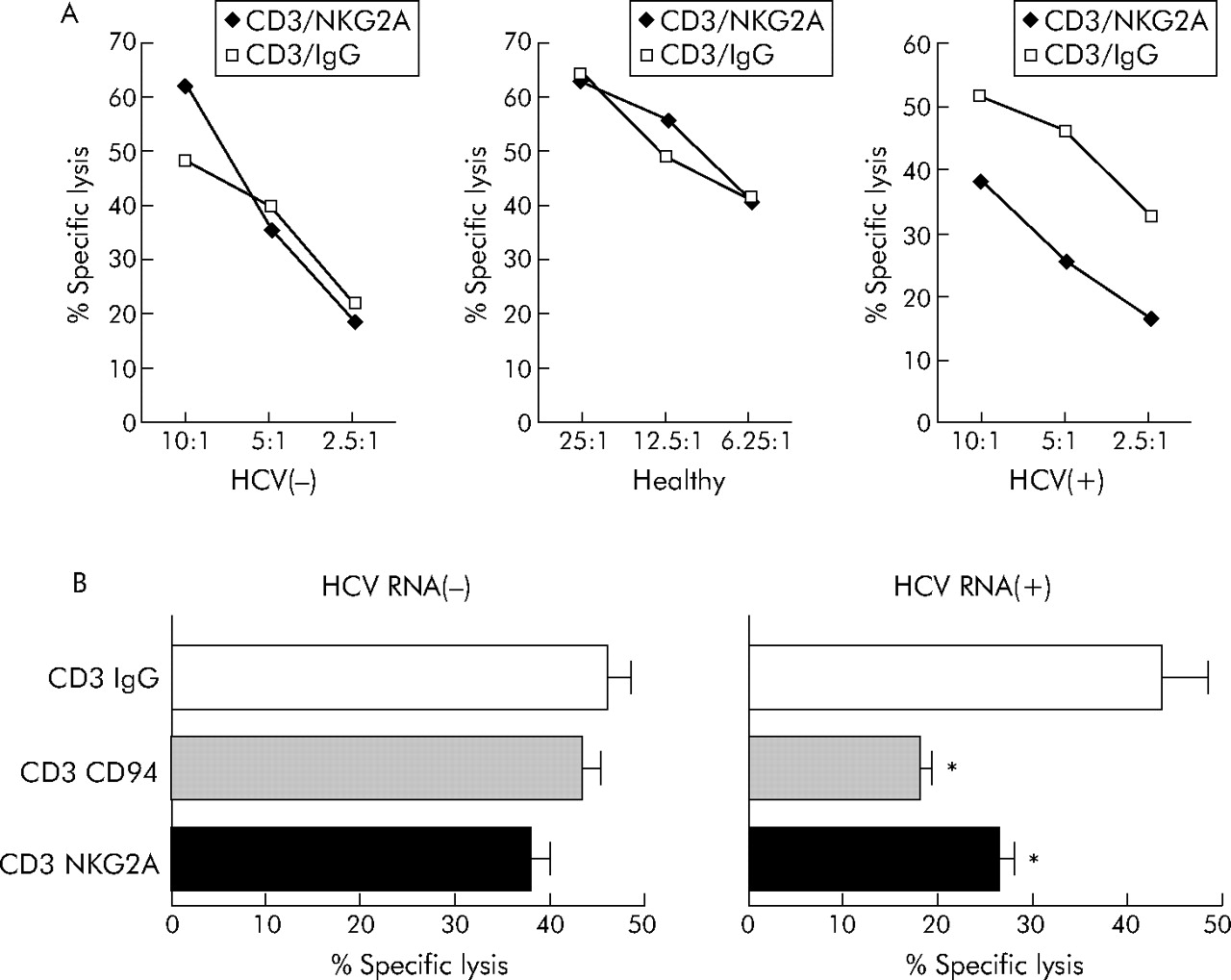
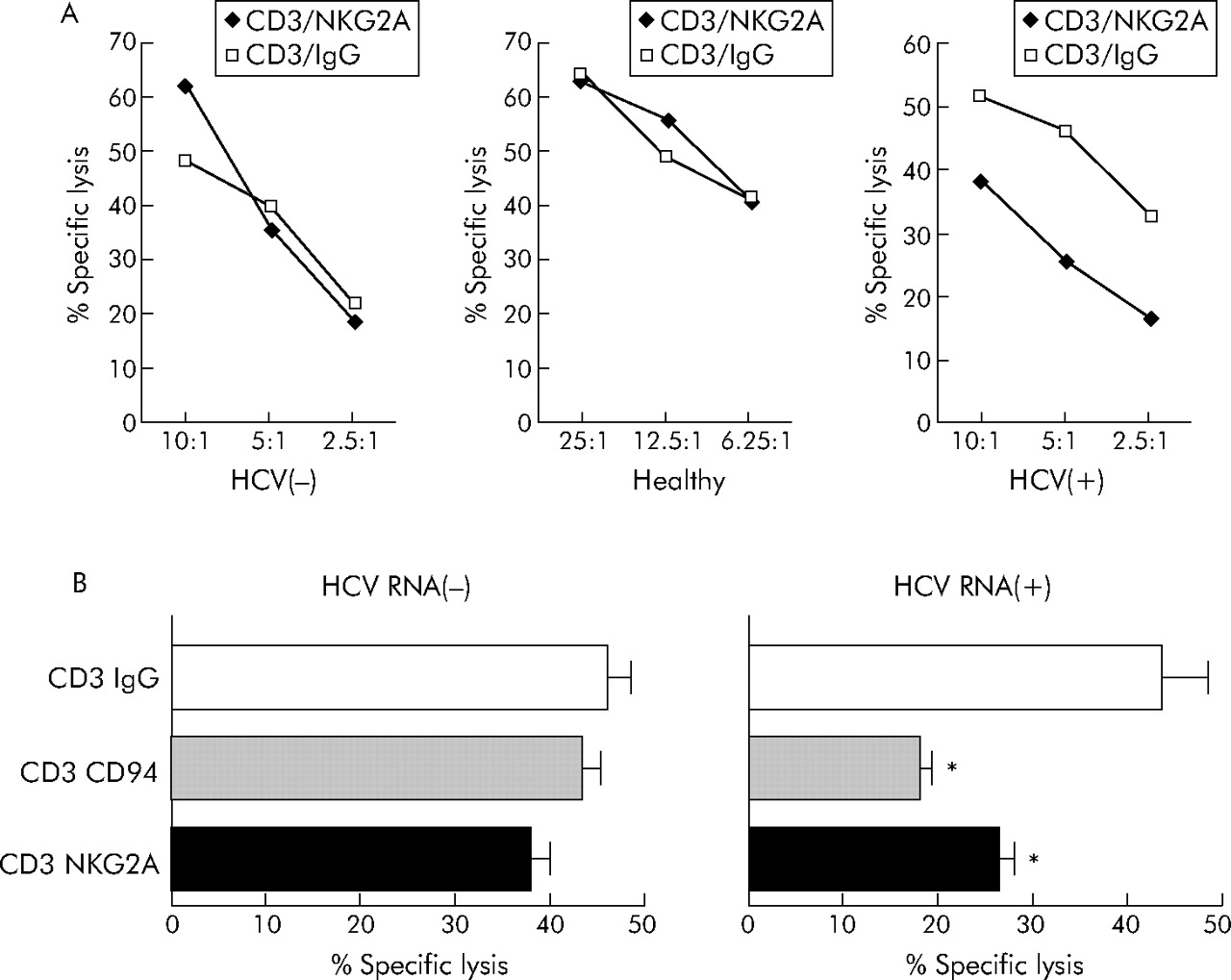
To examine the functional effect of enhanced NKG2A expression in chronic hepatitis C, we used a redirected lysis assay as described previously.30 Stimulation of purified CD3(+)CD8(+) T lymphocytes with a NKG2A specific antibody alone did not induce target lysis (data not shown). However, costimulation of the T cell receptor with anti-CD3 together with NKG2A using anti-NKG2A resulted in markedly reduced TCR mediated cytolysis compared with CD8+ T lymphocytes stimulated with anti-CD3 and an IgG isotype control antibody when T lymphocytes from HCV RNA positive patients were used as effector cells (fig 6A, right panel). In contrast, only small effects were observed when CD3+CD8+ cells from HCV RNA negative or healthy individuals were analysed in TCR/NKG2A costimulation experiments (fig 6A, left and middle panels).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
NKG2A mediated inhibition of CTL cytotoxicity. (A) Using a redirected cytolytic assay of P815 cells in the presence of anti-CD3 and anti-NKG2A monoclonal antibodies (mAb) or isotype control (anti-IgG mAb), we studied the function of NKG2A receptors in purified CD8(+) T cells derived from healthy individuals, subjects who became hepatitis C virus (HCV) RNA negative after antiviral therapy, or HCV RNA positive patients without therapy. As illustrated, we found that CD8(+) T cells from patients with replicative hepatitis C showed reduced cytolytic activity when NKG2A was costimulated with CD3. (B) If a CD94 specific mAb was used instead of anti-NKG2A to analyse the effects of NK cell receptors (NKR) on cytolytic function in NKG2A/NKG2C double positive CD8+ T cells derived from HCV RNA negative and HCV RNA positive subjects, simultaneous engagement of NKG2A and NKG2C by stimulation of CD94 in the redirected lysis assay resulted in markedly impaired cytolytic activity of CD8(+) T cells in HCV RNA(+) patients. *p<0.05, HCV(+) versus HCV(−).
As some (<10%) NKG2A positive CD8+ T lymphocytes also expressed the activating NKR NKG2C, which also forms heterodimers with CD94, we repeated the costimulation experiments using anti-CD94 instead of anti-NKG2A. Engagement of CD94 also resulted in reduced lytic function of CTL from HCV RNA positive patients (fig 6B).
DISCUSSION
Natural killer cell receptors are important regulators of NK cell9,27,32–36 and CD8+ T cell functions.10–12,37 Regarding the fact that impaired activities of NK cells4–6,8,22 and CD8(+) T cells1–3 have been reported in chronic HCV infection, we analysed whether dysregulated expression of NKR on these lymphocyte subpopulations might be involved in the inefficient cellular immune responses observed in chronic hepatitis C.
Concerning C-type lectin receptors, we found enhanced expression of the inhibitory NKG2A receptor on both NK cells and CD8(+) T lymphocytes. The natural ligand for the inhibitory NKG2A receptor is the non-classical MHC I molecule HLA-E.22 Recently, we demonstrated that HCV infected hepatocytes express enhanced levels of HLA-E. Moreover, we showed that the HLA-A2 restricted epitope HCV core aa35–44 stabilises HLA-E expression resulting in impaired cytolysis mediated by NK cells.22 Corroborating these findings, we now demonstrate that enhanced surface expression of NKG2A on NK cells from HCV infected patients is associated with reduced cytotoxic activity against HLA-E expressing cells. In addition to direct impairment of cytotoxic function, inhibition of NK cell activity by upregulated NKG2A may have further consequences. For instance, enhanced expression of NKG2A has also been shown to result in altered NK cell induced modulation of dendritic cell functions in chronic hepatitis C.8
Regarding CD8+ T cells, expression of NKG2A has been suggested to be involved in fine tuning of T cell responses by raising the threshold for TCR triggering,38 thereby serving as a mechanism for peripheral tolerance.10 In line with this concept, the CD94/NKG2A receptor has been shown to inhibit cytokine production and cytolytic activity of antigen specific T cells,39 and the inability of antiviral CD8+ T cells to control virus induced tumours has been attributed to upregulation of CD94/NKG2A receptors on T cells.12 In line with this hypothesis, upregulation of NKG2A due to chronic exposure to a pathogen might be a mechanism to prevent clonal exhaustion by terminal differentiation or to suppress excessive immunopathological responses. On the other hand, a premature shut off of the antiviral CTL response due to induction of NKG2A would facilitate viral persistence and thus could contribute to viral immune evasion.11,12 We found that enhanced NKG2A expression on CD8+ T cells in chronic hepatitis C was associated with impaired cytolytic function of T cells when NKG2A was costimulated with the TCR. In line with the findings of Jabri and colleagues,11 we also found that engagement of CD94 instead of NKG2A, which simultaneously stimulates NKG2A and NKG2C, resulted in TCR inhibition. Thus engagement of inhibitory NKG2A is dominant over engagement of activating NKG2C, which was also present on CD8+ T cells. Similar to what was observed for NK cells, we found reduced cytolytic activity of CD8+ T cells from HCV infected patients against HLA-E expressing cells. Thus simultaneous upregulation of NKG2A and its ligand HLA-E in chronic hepatitis C might represent an effective viral strategy to evade immune surveillance by acting on both NK cells and CD8+ T cells.
In contrast with C-type lectin receptors, the NCR NKp30, NKp44, and NKp46 are selectively expressed on NK cells.32–34 Analysing intrahepatic as well as circulating lymphocyte populations, we observed that chronic hepatitis C was associated with a reduced percentage of NKp30 and NKp46 expressing CD56+ NK cells.
Dysregulated expression of NCR in hepatitis C resembles recent findings by De Maria and Moretta who demonstrated significantly reduced surface expression of NKp30 and NKp46, but not NKp44, in HIV-1 infected patients.27 In line with our results, this group also found that altered NCR expression was associated with a reduced lytic capability of NK cells. Thus downregulated expression of NCR seems to be a mechanism used by several different viruses to counteract NK cell function. However, our findings that HBV infection is not accompanied by reduced expression of NCR suggests that downregulation of NCR is a specific phenomenon linked to distinct viruses. Furthermore, our finding of lower numbers of NCR expressing NK cells in HCV(+) livers than in livers of patients with hepatic diseases not associated with hepatitis C argues against a general reduced expression of NCR due to hepatic inflammation.
The mechanism(s) operative in regulation of NKR is at present only incomplete understood. Recent data suggest that changes in the microenvironment, possibly altered secretion of cytokines, might be involved. Castriconi et al showed that transforming growth factor β (TGF-β) downregulates surface expression of NKp30 but not NKp46 or NKp44 on NK cells36 and in vitro studies proposed cytokine stimulation by IL-15 and TGF-β11,40 as inducing factors of NKG2A expression. Ray et al proposed upregulation of TGF-β in HCV infection and a probable role for this cytokine in the pathogenesis of HCV related chronic liver disease.41 However, we did not find different serum levels of TGF-β and IL-15 between the study groups (data not shown).
Thus it remains to be resolved which other cytokines and signalling pathways are involved in altered NKR regulation in chronic hepatitis C.
The finding that NCR expression levels in patients who had cleared the virus by antiviral therapy were comparable with those observed in healthy individuals together with the fact that a significant correlation existed between NKG2A expression on CD8+ T cells and HCV serum levels supports the concept that reduced expression is linked to HCV replication or to changes in the cellular microenvironment induced by HCV replication.
The activation status of both NK cells and CD8+ T cells depends on the balance between activating and inhibitory signals delivered by surface receptors. Thus our present findings of upregulated expression of inhibitory NKG2A receptor combined with concomitant downregulation of activating NKp30 and NKp46 synergistically leads to the dominant delivery of inhibitory signals to NK cells and NKR positive T cells. This altered NKR expression is reflected by impaired lytic function of both NK cells and CD8(+) T cells. Therefore, dysregulation of NK cell receptor expression provides a plausible explanation for at least one mechanism presumably contributing to defective cellular immune functions in HCV infected patients. Future experimental work is needed to clarify if and to what extent dysregulation of NKR expression facilitates the establishment and maintenance of chronic hepatitis C.
Acknowledgments
This work was supported by the Deutsche Forschungsgemeinschaft, SFB 704, Teilprojekt A6, and by the Kompetenznetz Hepatitis, Teilprojekt 11.3.3.
The authors thank Agathe Iwan and Monika Schulz for excellent technical assistance.
REFERENCES
Footnotes
-
Published online first 1 December 2005
-
Conflict of interest: None declared.