Article Text

Abstract
Background and aims: Impairment of the gastrointestinal mucosal barrier contributes to progression of HIV infection. The purpose of this study was to investigate the effect of highly active antiretroviral therapy (HAART) on the HIV-induced intestinal barrier defect and to identify underlying mechanisms.
Methods: Epithelial barrier function was characterised by impedance spectroscopy and [3H]mannitol fluxes in duodenal biopsies from 11 untreated and 8 suppressively treated HIV-infected patients, and 9 HIV-seronegative controls. The villus/crypt ratio was determined microscopically. Epithelial apoptoses were analysed by terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick-end labelling (TUNEL) and caspase-3 staining. Tight junction protein expression was quantified by densitometric analysis of immunoblots. Mucosal cytokine production was determined by cytometric bead array.
Results: Only in untreated but not in treated HIV-infected patients, epithelial resistance was reduced (13 (1) vs 23 (2) Ω cm2, p<0.01) and mannitol permeability was increased compared with HIV-negative controls (19 (3) vs 9 (1) nm/s, p<0.05). As structural correlates, epithelial apoptoses and expression of the pore-forming claudin-2 were increased while expression of the sealing claudin-1 was reduced in untreated compared with treated patients and HIV-negative controls. Furthermore, villous atrophy was evident and mucosal production of interleukin 2 (IL2), IL4 and tumour necrosis factor α (TNFα) was increased in untreated but not in treated HIV-infected patients. Incubation with IL2, IL4, TNFα and IL13 reduced the transepithelial resistance of rat jejunal mucosa.
Conclusions: Suppressive HAART abrogates HIV-induced intestinal barrier defect and villous atrophy. The HIV-induced barrier defect is due to altered tight junction protein composition and elevated epithelial apoptoses. Mucosal cytokines are mediators of the HIV-induced mucosal barrier defect and villous atrophy.
Statistics from Altmetric.com
As a major compartment of CD4+ T cell depletion and viral replication,1–5 the gastrointestinal mucosa is a main target of HIV infection. Recent data indicate that the intestinal epithelial barrier function may play a central role in disease progression in HIV infection. The finding of raised plasma levels of lipopolysaccharides (LPS) associated with LPS stimulation of monocytes6 gave rise to the hypothesis that HIV-induced disruption of the epithelial barrier leads to enhanced translocation of luminal antigens such as LPS across the intestinal epithelium. After saturation of counteracting and neutralising mechanisms this antigen translocation is thought to elicit the chronic immune activation,7 which is known as a main driving force for the progressive immune failure in chronic HIV infection.8–10
In several studies, untreated HIV infection has indeed been shown to be associated with increased permeability of the intestinal epithelium.11–15 This barrier defect occurs independently of opportunistic infections11 12 14 16 and therefore reflects the impact of HIV infection and its immunological sequelae on epithelial integrity. Structurally, chronic HIV infection causes a transformation of the intestinal mucosa with hyporegeneration and dysmaturation of intestinal epithelial cells, resulting in partial villous atrophy.17–21 Both mucosal transformation and the epithelial barrier defect are thought to be a consequence of mucosal T cell activation with increased production of inflammatory cytokines.15 18 22
With respect to these pieces of evidence, a model can be conceptualised whereby HIV replication leads to mucosal T cell activation, which, by local secretion of inflammatory cytokines, causes impairment of the epithelial intestinal barrier function. The resulting epithelial barrier defect in turn further fuels mucosal as well as systemic immune activation. As restoration of epithelial integrity by direct intervention on the epithelial level is not feasible, the only way to interrupt the self-perpetuating cycle of mucosal immune activation would be to reduce immune activation by suppression of HIV replication. In line with this reasoning, the decrease of serum LPS levels observed in HIV-infected patients treated by highly active antiretroviral therapy (HAART) was interpreted as a result of reconstitution of the gastrointestinal epithelial barrier.6 However, the effect of HAART on the epithelial barrier function of HIV-infected patients has not been investigated so far. Furthermore, there is a paucity of data regarding the underlying mechanism of the HIV-induced mucosal barrier defect.
In this study, we investigated barrier function and morphology of duodenal mucosa specimens obtained from HIV-infected patients who were either untreated or suppressively treated by HAART. In treated patients we observed normalisation of both the mucosal barrier defect and villous atrophy. Suppressive HAART also normalised elevated mucosal cytokines, implicating a close inter-relationship between viral replication, mucosal cytokine activation and the HIV-induced mucosal barrier defect.
MATERIALS AND METHODS
Study subjects
Eleven treatment-naive and eight HIV-infected patients treated by HAART were included in the study. Nine HIV-negative patients without gastrointestinal pathology served as controls. Clinical details of the patients are given in table 1. In treated patients, HIV replication had been suppressed below the limit of detection (quantitative PCR <50 copies/ml, Roche Amplicor, Roche, Mannheim, Germany) by HAART for a median time of 25 months (range 3–50). Antiretroviral therapy consisted of a standard combination of two nucleoside reverse transcriptase inhibitors together with either a non-nucleoside reverse transcriptase inhibitor or a ritonavir-boosted protease inhibitor. HIV-infected patients underwent endoscopy for symptoms related to the upper gastrointestinal tract, and control patients for exploration of unexplained anaemia. Patients with intestinal infections (excluded as described previously14), diarrhoea, inflammatory bowel disease or a pathological lesion of the duodenum and/or antrum were excluded. The study was approved by the local ethics committee and all patients gave written informed consent to participation in the study.
Tissue preparation
Mucosal specimens were obtained from the lower duodenum with a 3.4 mm forceps. After immediate transport to the laboratory, a plastic ring (inner diameter 2.5 mm) was glued to the serosal side of the biopsy specimens with histoacryl tissue glue (Braun, Melsungen, Germany) with the aid of a dissection microscope. The disc-mounted mucosa was placed into a modified Ussing-type chamber as described.23
Electrophysiological experiments
Impedance spectroscopy was performed as described previously.14 24 Using the capacitative properties of the epithelium, this technique resolves the transmural wall resistance (Rt) into its epithelial (Re) and subepithelial (Rsub) component.14 24 Mannitol permeability, which is confined to paracellular pathways, was assessed from [3H[mannitol mucosal to serosal fluxes as described,25 with 10 mM of unlabelled mannitol present on both sides and an exposed epithelial area of 0.049 cm2. In additional experiments, transepithelial resistance was determined in conventional Ussing chambers on rat jejunal mucosa (tissues obtained from male albino Wistar rats) partially stripped from the underlying submucosal tissue as described.26 For these experiments, rat cytokines (PeproTech GmbH, Hamburg, Germany) were used as indicated. The bathing solution contained (mmol/l): Na+, 140; Cl−, 123.8; K+, 5.4; Ca2+, 1.2; Mg2+, 1.2; HPO42−, 2.4; H2PO4−, 0.6; HCO3−, 21, d(+)-glucose, 10; β-OH-butyrate, 0.5; glutamine, 2.5; and d(+)-mannose, 10. The solution was gassed with 95% O2/5% CO2, its temperature was maintained at 37°C using water-jacketed reservoirs, and the pH was 7.4. Antibiotics (50 mg/l azlocillin and 4 mg/l tobramycin) added to the bathing solution prevented bacterial growth and—as shown in preliminary experiments—had no effect on ion fluxes and resistance in the concentration used.26
Morphological analysis and epithelial apoptosis
Mucosal samples were obtained in close proximity to the biopsy site of the samples used for the electrophysiological experiments. Biopsy specimens were fixed in 4% formalin and embedded in paraffin for quantification of epithelial apoptoses or fixed in ethanol/glacial acetic acid (3:1) and stored in 75% ethanol for morphological analysis. For immunofluorescence detection of epithelial apoptoses, serial sections (3 μm) were dewaxed and cellular DNA was stained with terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick-end labelling (TUNEL) assay (Roche, Mannheim, Germany). Blunt ends of double-stranded DNA exposed by strand breaks were visualised by enzymatic labelling of the free 3′-OH termini with fluorescein/deoxyuridine triphosphate. For immunohistochemical staining of caspase-3 activation, slides were subjected to a heat-induced epitope retrieval step before incubation with the primary polyclonal rabbit antibody against cleaved caspase-3 (Cell Signaling, Danvers, Massachusetts, USA, dilution 1:100). For detection, the streptavidinAP kit (K5005, Dako, Glostrup, Denmark) was used and alkaline phosphatase was revealed by Fast Red as chromogen. Epithelial apoptoses were determined as the percentage of epithelial cells with apoptotic nuclei or caspase-3-positive epithelial cells per visual field (apoptotic ratio). An average of six visual fields (∼150 enterocytes per field) was counted in each sample. Morphological analysis was performed in H&E-stained sections by use of analySIS software (version 3.0, Soft Imaging System, Münster, Germany). If necessary, repeated sections in different axes were surveyed until a section within the villus axis plane was obtained. The mean height and mean depth of a minimum of 10 villi and 10 crypts, respectively, were determined in each specimen. The morphological analysis and the quantification of epithelial apoptosis were performed without knowledge of the respective specimen’s patient status.
Western blot analysis
For detection of tight junction protein expression, western blot analysis was performed on membrane extracts of duodenal biopsy specimens as described previously.25 Primary antibodies were rabbit polyclonal immunoglobulin G (IgG) antibodies against claudin-1, claudin-2 and occludin, and primary mouse monoclonal IgG antibodies against claudin-4. Peroxidase-conjugated goat anti-rabbit IgG or goat anti-mouse IgG antibodies and the chemiluminescence detection system Lumi-LightPLUS western blotting kit (Roche, Mannheim, Germany) were used for detection of bound antibodies. Synthesis and specificity of these antibodies (Zymed, San Francisco, California, USA) were described by Rahner et al.27 Chemiluminescence signals were detected using a LAS-1000 imaging system (Fuji, Tokyo, Japan) and densitometrically analysed with the AIDA program package (Raytest, Berlin, Germany).
Mucosal cytokine secretion
Determination of mucosal cytokine secretion was performed as the last of all the substudies, and additional samples from HIV-negative controls had been collected then (n = 13, except for interleukin 12 (IL12) with n = 15 controls). On the other hand, due to limited sample numbers, fewer samples obtained from HIV-infected patients were subjected to cytometric bead array (CBA) (naive, 7; HAART, 6) than to the other studies. Biopsies were cultured for 48 h as described before28 and cytokines produced in situ were quantified in the supernatants with a CBA (Becton Dickinson, Heidelberg, Germany) according to the manufacturer’s protocol.
Mucosal IL13-positive cells
Formalin-fixed and paraffin-embedded 4 μm sections of duodenal tissue were deparaffinised and subjected to epitope retrieval in proteinase K solution (Dako) for 5 min. For IL13 staining, slides were incubated for 45 min with rabbit antibody against human IL13 (Chemicon, Temecula, California, USA) followed by 30 min with donkey anti-rabbit biotin (Dianova, Hamburg, Germany), streptavidin–alkaline phosphatase (Dako), and Dako Red as chromogen. Nuclei were counterstained with haematoxylin. All steps were performed at room temperature.
Statistical analysis
Results are given as means (SEM). In univariate analysis, significance was tested for by the two-tailed Student t test. Multivariate analysis was performed by means of one-way analysis of variance (ANOVA). Bonferroni’s multiple comparison test was used for post hoc analysis. p Values <0.05 were considered significant.
RESULTS
Effect of HAART on duodenal epithelial barrier function
Transepithelial resistance as determined in a conventional Ussing in vitro set-up is composed of the epithelial (Re) and the subepithelial resistance (Rsub). However, the effective diffusion barrier in vivo is represented by Re only.29 Therefore, we performed impedance spectroscopy which allowed specific determination of Re and Rsub.
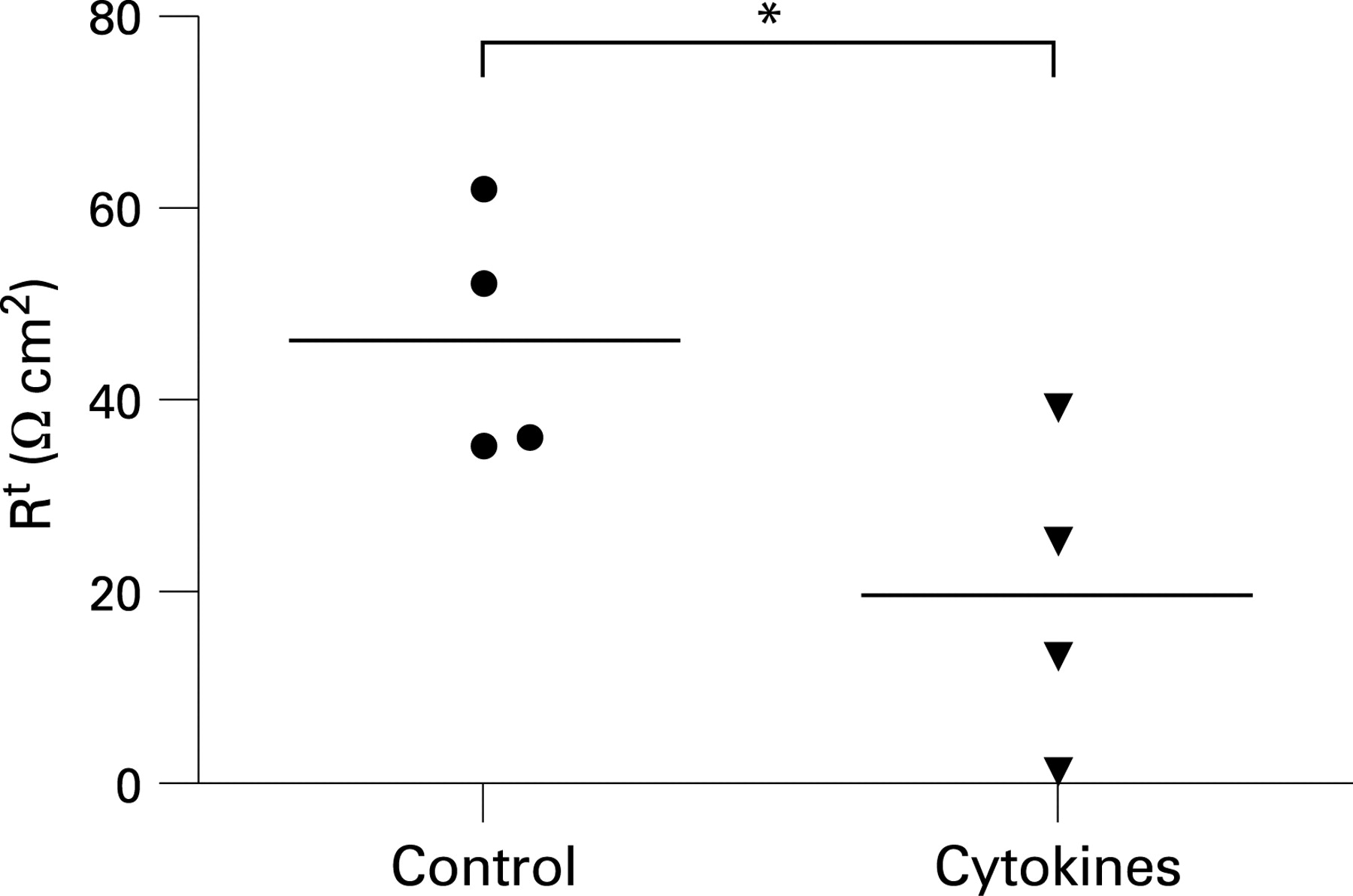
Original impedance locus plots are depicted in fig 1A–C. As shown in fig 1D, the Re of untreated HIV-infected patients was reduced by ∼40%, whereas in treated HIV-infected patients Re was not different from that of controls (p = 0.32). On the other hand, Rsub—roughly reflecting the thickness of subepithelial tissue layers—did not differ between HIV-infected patients and HIV-negative controls (26.6 (2.0) Ω cm2 in untreated HIV-infected patients (p = 0.15), 30.8 (3.2) Ω cm2 in treated HIV-infected patients (p = 0.93) and 31.2 (2.6) Ω cm2 in controls).

In order to characterise the change in Re further, we performed mannitol fluxes to assess paracellular permeability. As shown in fig 1E, there was a >100% increase of the mannitol permeability in untreated patients, whereas mannitol permeability of treated patients was not different from that of controls (p = 0.90). Although there was a trend towards a higher CD4 count in treated compared with untreated patients, no correlation of peripheral CD4 cells with Re or mannitol fluxes was found either in treated or in untreated patients (data not shown). Taken together, electrophysiological as well as flux data indicate that HIV infection causes a pronounced impairment of the intestinal epithelial barrier function. This HIV-induced mucosal barrier defect was fully reversed after HIV replication was suppressed by effective HAART, and therefore seems to be linked to active viral replication.
Morphological changes
In untreated HIV-infected patients, we found a one-third reduction of villus height, whereas crypt depth was not different from that of HIV-negative controls (fig 2). On the other hand, in HIV-infected patients treated by HAART, villus height was not different from that of HIV-negative controls (p = 0.33 and p = 0.07, respectively). Thus, the partial villous atrophy associated with HIV infection is reversed in patients with suppressed viral replication, and the restoration of barrier function following suppressive HAART is paralleled by reconstitution of epithelial morphology.

Mechanisms of HIV-induced barrier impairment
Increased paracellular permeability can be caused by gross violations of the epithelial continuity (eg, ulcers and/or erosions), by an increased epithelial apoptosis and by an increased permeability of the epithelial tight junctions. Since the duodenal biopsies of our patients did not show any erosive or ulcer-type lesions on histological examination, the first possibility was ruled out.
The apoptotic ratio within the duodenal epithelium was increased twofold in untreated HIV-infected patients as compared with treated HIV-infected patients or HIV-negative controls (fig 3). Thus, the reconstitution of the epithelial barrier function induced by HAART is paralleled by a reduction in the epithelial apoptotic ratio, indicating increased frequency of apoptotic epithelial cells as a mechanism of the HIV-induced intestinal barrier defect.

As a prominent role for IL13 for the mucosal barrier defect in ulcerative colitis has recently been described,30 we immunohistochemically quantified IL13-positive mucosal cells in an additional analysis. Compared with HIV-negative controls (2 (2) cells per 10 high-power field (hpf), n = 3), IL13-positive cells were markedly increased in both untreated and treated HIV-infected patients (48 (3) cells per 10 hpf, p<0.05 vs controls, n = 4, and 73 (12) cells per 10 hpf, p<0.01 vs controls, respectively, n = 3), but the two groups of HIV-infected patients showed no difference from each other (data not shown).
Immunoblots with subsequent densitometric analysis were used for quantitative analysis of the membrane-bound tight junction proteins occludin and the sealing (claudin-1, claudin-4) or pore-forming (claudin-2) claudins (fig 4). In untreated HIV-infected patients, claudin-1 expression was depressed by ∼40% and claudin-2 expression was strongly increased, whereas in treated patients claudin-1 and claudin-2 expression was not significantly different from that of HIV-negative controls (p = 0.57 and p = 0.95, respectively, fig 4A,B). Occludin and claudin-4 expression, on the other hand, was not significantly different from that of HIV-negative controls in both untreated and treated HIV-infected patients (occludin, p = 0.92 and p = 0.06; claudin-4, p = 0.35 and p = 1.0, respectively; fig 4A). The decreased expression of the sealing claudin-1 and increased expression of the pore-forming claudin-2 found in untreated patients with HIV indicate altered composition of the epithelial tight junctions as a second mechanism for the HIV-induced intestinal barrier defect.

Mucosal cytokines and duodenal barrier function
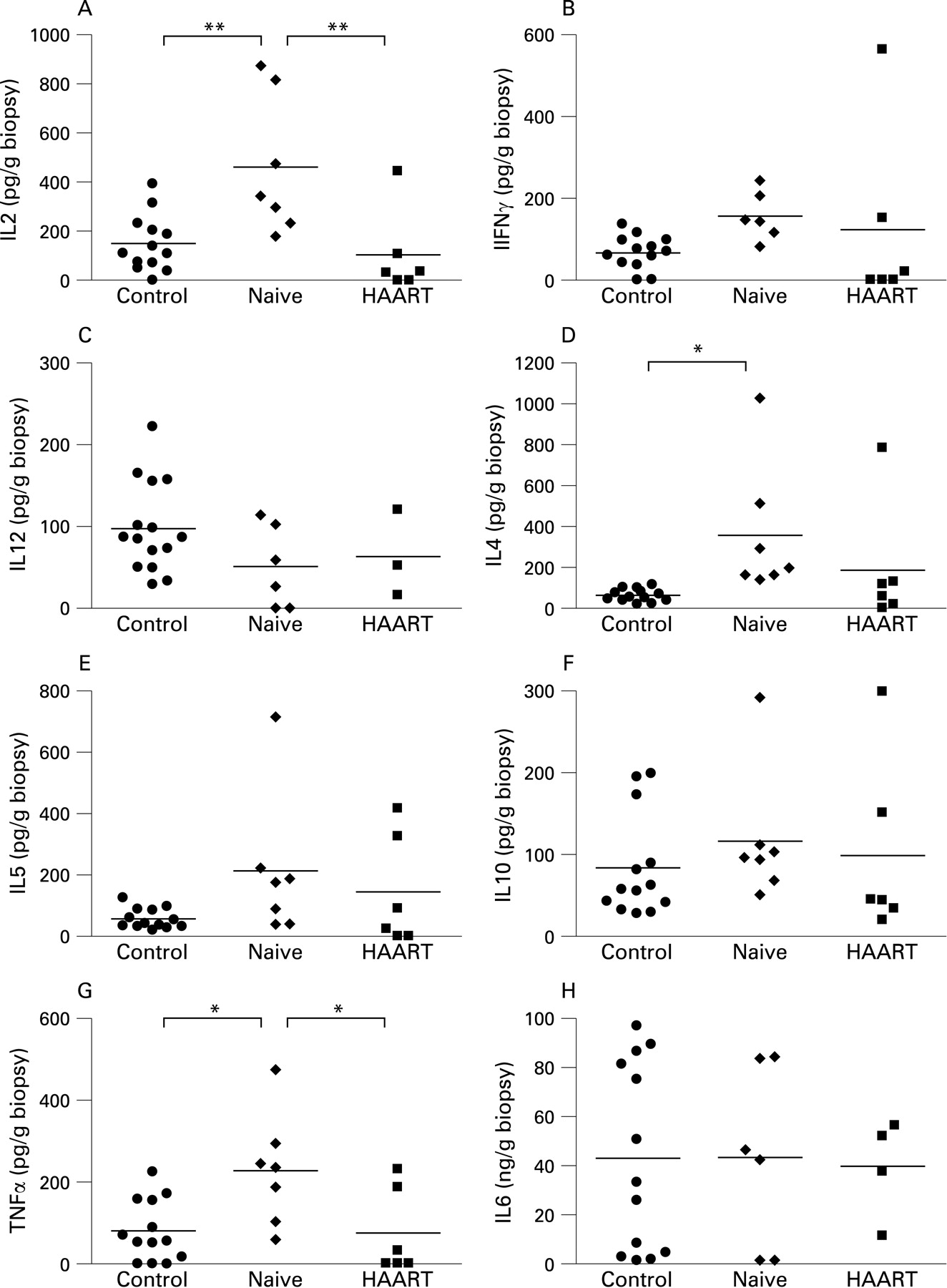
In mucosal samples from untreated HIV-infected patients, production of the Th1 cytokine IL2 (fig 5A), but not of interferon γ (IFNγ) (fig 5B) and IL12 (fig 5C) was elevated over that of HIV-negative controls. In a similar way, production of the Th2 cytokine IL4 (fig 5D), but not of IL5 (fig 5E) and IL10 (fig 5F) was elevated in untreated patients, and also the proinflammatory cytokine tumour necrosis factor α (TNFα) (fig 5G), but not IL6 (fig 5H) was elevated in these patients. Thus, the enhanced mucosal cytokine production observed in untreated HIV-infected patients did not fit into a classic Th1 or Th2 pattern, but rather represents unspecific mucosal cytokine activation. In treated patients, on the other hand, mucosal production of Th1, Th2 or other proinflammatory cytokines was not significantly different from controls (p>0.05; fig 5), indicating normalisation of mucosal cytokine production in suppressively treated patients.

In order to investigate the barrier effect of the mucosal cytokines, which were increased in our untreated patients, we incubated rat jejunal mucosa with a cytokine cocktail containing IL2, IL4, IL13 and TNFα. As shown in fig 6, the cytokine cocktail caused a drop of the transepithelial resistance in this epithelium indicating induction of a small intestinal mucosal barrier defect by the concerted action of these cytokines.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
An intestinal epithelial barrier defect is considered a key mechanism for immune activation and disease progression in HIV infection,6 but the effect of HAART on barrier function of the gastrointestinal epithelium was unknown prior to our study. Here we report complete resolution of the HIV-induced intestinal mucosal barrier defect as well as reversal of villous atrophy in patients treated by suppressive HAART. Thus, the HIV-induced gastrointestinal barrier defect and mucosal transformation are not irreversible but rather seem to depend on active viral replication. As antiretroviral drugs induce rather than improve malfunction of the mucosal barrier,31 it seems very unlikely that HAART exerts a direct beneficial effect on the mucosal barrier. Our findings therefore not only corroborate the independent aetiological role of HIV replication for mucosal damage suggested in earlier studies,17 19–21 32 33 but also provide a straightforward explanation for the reduction of LPS translocation in HIV-infected patients treated by HAART.6
To perform a mechanistic characterisation of the HIV-induced mucosal barrier defect, we looked for associations between structural alterations, impaired mucosal barrier function and their responses to HAART. In this way, the increased frequency of apoptotic epithelial cells in untreated patients was reduced by treatment with HAART. The intermediately elevated frequency of epithelial apoptotic cells in treated patients could be explained by proapoptotic effects of the antiretroviral therapy,31 although other factors related to HIV infection may also play a role. As even moderate increases of epithelial apoptoses affect epithelial barrier function,34–37 the >100% increased frequency of apoptotic epithelial cells in untreated HIV-infected patients very probably contributes to the mucosal barrier defect. This notion is also supported by recent studies which used high-resolution in vivo imaging to demonstrate formation of transient epithelial gaps during cell shedding from the intestinal mucosa.38 39 Under control conditions, these gaps did not increase the epithelial permeability. Intraperitoneal administration of TNFα, which is a recognised inducer of apoptosis, however, increased both the number of epithelial gaps and epithelial permeability.38 39
Claudins are the main components of the epithelial tight junctions which restrict the movement of solutes, including ions and small antigens, across the gastrointestinal mucosa.40 41 For claudin-1, sealing properties have been demonstrated.42 43 Claudin-2, on the other hand, has been characterised as a paracellular channel for small cations,44 and overexpression of claudin-2 was associated with tight junction strand discontinuation.45 Therefore, the downregulation of claudin-1 combined with increased expression of claudin-2 in untreated patients with HIV implies an increased permeability of the epithelial tight junction contributing to the overall barrier defect observed in these patients.
Numerous studies have demonstrated induction of a mucosal barrier defect by inflammatory cytokines in intestinal model epithelia22 30 36 46–50 as well as in native intestinal mucosa.26 30 34 51 However, conflicting results were reported as to whether or not inflammatory cytokines are actually increased in the intestinal mucosa of untreated HIV-infected patients. Based on tissue RNA quantification, increased mucosal cytokine expression has been described,52–56 but mononuclear cells isolated from the colon of HIV-infected patients did not show increased production of cytokines.57 58 In contrast to the latter findings, we found increased mucosal production of TNFα, IL2 and IL4 in our untreated HIV-infected patients. As we quantified cytokine secretion from intact mucosal biopsies, the failure of isolated mononuclear cells to produce increased amounts of cytokines57 58 may be explained by a lack of co-stimulation “physiologically” provided by the mucosal microenvironment. Finally, a cocktail of the cytokines suspected to cause the barrier defect according to our data evoked a significant barrier defect in rat jejunal mucosa, further corroborating the link between raised inflammatory cytokines and mucosal barrier impairment.
Of the mucosal cytokines found to be increased in our untreated patients, TNFα unequivocally has been shown to affect mucosal barrier function,22 30 34 36 46 49 51 whereas the mucosal action of IL2 and IL4 is less well described. TNFα induced villous atrophy,59 epithelial apoptosis22 30 and removal of claudin-1 from the apical membrane.50 In addition, TNFα decreased the epithelial resistance and reduced claudin-1 but increased claudin-2 expression in intestinal model epithelia.34 48 60 61 Our observation that TNFα and other mucosal cytokines were not increased in patients with suppressed HIV replication further substantiates the hypothesis that villous atrophy, increased epithelial apoptosis and altered tight junction composition in untreated HIV infection are probably maintained by TNFα, with a potential contribution of additional mucosal cytokines such as IL1, IL4 and IL13.
In conclusion, we propose the following model of HIV-induced damage of the gastrointestinal epithelial barrier. Active HIV replication leads to a local increase of inflammatory cytokines within the gastrointestinal mucosa.53 These cytokines induce impairment of the epithelial barrier function by triggering epithelial apoptosis and by alteration of tight junction protein expression. The ensuing barrier defect facilitates translocation of enterobacteria62 63 and bacterial antigens such as LPS.6 Increased antigenic exposure further stimulates mucosal cytokine production, promoting mucosal and possibly systemic immune activation.6 Suppression of HIV replication by HAART interrupts the self-perpetuating cycle of mucosal immune activation and barrier impairment and allows for reconstitution of the epithelial barrier function.
Acknowledgments
The authors wish to express their gratitude for the excellent technical assistance provided by Anja Fromm, Diana Böse, and Simone Spieckermann.
REFERENCES
Footnotes
Funding: This work was supported by the German Research Foundation (DFG KFO grant 104).
Competing interests: None.
Ethics approval: The study was approved by the local ethics committee.
Linked Articles
- Digest