Article Text

Abstract
Background and aims: Progression of chronic cholestatic disorders towards ductopenia results from the dysregulation of cholangiocyte survival, with cell death by apoptosis prevailing over compensatory proliferation. Currently, no therapy is available to sustain cholangiocyte survival in the course of those disorders. It was recently shown that cholangiocytes express the glucagon-like peptide-1 receptor (GLP-1R); its activation results in enhanced proliferative reaction to cholestasis. The GLP-1R selective agonist exendin-4 sustains pancreatic β cell proliferation and prevents cell death by apoptosis. Exendin-4 is now employed in humans as a novel therapy for diabetes. The aim of the present study was to verify whether exendin-4 is effective in preventing cholangiocyte apoptosis.
Methods: In vitro, tests were carried out to determine if exendin-4 is able to prevent apoptosis of cholangiocytes isolated from normal rats induced by glycochenodeoxycholic acid (GCDCA); in vivo, animals subjected to 1 week of bile duct ligation and to a single intraperitoneal injection of CCl4 were treated with exendin-4 for 3 days.
Results: Exendin-4 prevented GCDCA-induced Bax mitochondrial translocation, cytochrome c release and an increase in caspase 3 activity. Phosphatidylinositol 3-kinase, but not cAMP/protein kinase A or Ca2+/calmodulin-dependent protein kinase inhibitors, neutralised the effects of exendin-4. In vivo, exendin-4 administration prevented the increase in TUNEL (terminal deoxynucleotidyl transferase-mediated triphosphate end-labelling)-positive cholangiocytes and the loss of bile ducts observed in bile duct-ligated rats treated with CCl4.
Conclusion: Exendin-4 prevents cholangiocyte apoptosis both in vitro and in vivo; such an effect is due to the ability of exendin-4 to counteract the activation of the mitochondrial pathway of apoptosis. These findings support the hypothesis that exendin-4 may be effective in slowing down the progression of cholangiopathies to ductopenia.
Statistics from Altmetric.com
Cholangiopathies are a wide array of congenital or acquired disorders that share the feature of being chronic cholestatic conditions leading to liver failure.1 Despite the different aetiology, cholangiopathies share the common feature of primarily targeting cholangiocytes, the epithelial cells lining the intrahepatic biliary tree.1 These diseases are characterised by the progressive vanishing of bile ducts (eg, ductopenia) that results from an abnormal cholangiocyte homeostasis.1 It is thought that ductopenia results from excessive cell death by apoptosis that prevails over the ineffective cholangiocyte compensatory proliferative response.1 2
Cholangiopathies are a challenge for clinicians: 20% of liver transplants among adults and 50% of those among paediatric patients are due to these disorders.3 This is associated, at least in part, with the fact that there is no therapy effective in maintaining an adequate cholangiocyte survival.1 The factors that regulate the balance between proliferation and death of cholangiocytes are undefined; such a lack of pathophysiological knowledge may contribute to slowing down the development of effective therapies for cholangiopathies.
Exendin-4 is a “long-lasting” analogue of glucagon-like peptide-1 (GLP-1).4 GLP-1 is secreted by enteroendocrine L cells and modulates the biology of a number of cells, by interacting with a specific G-protein-coupled receptor (GLP-1R).4–6 GLP-1 modulates glucose homeostasis, by preventing pancreatic β cell death by apoptosis and by eliciting their proliferation.4–6 As a consequence, exendin-4 is now successfully employed in humans as a novel antidiabetic treatment.7
GLP-1 serum levels are increased in the course of cholestasis.8 We have recently shown that cholangiocytes are susceptible to the action of GLP-1. They express the GLP-1R, the activation of which (by GLP-1 or exendin-4) results in a marked increase in cell growth, both in vitro and in vivo.9 GLP-1 action on cholangiocytes is required for their proliferative response to cholestasis.9
Therefore, the aim of this study was to verify if the GLP-1 analogue exendin-4 is also able to prevent cholangiocyte cell death by apoptosis. Thus, we asked the following questions. (1) Does exendin-4 prevent cholangiocyte apoptosis in vitro? (2) Which are the intracellular pathways that mediate the antiapoptotic effects of exendin-4 in cholangiocytes? (3) Is exendin-4 effective in sustaining cholangiocyte survival in vivo?
METHODS
Materials
Reagents were purchased from Sigma-Aldrich (Milan, Italy) unless otherwise indicated. Antibodies for immunoblotting were purchased from Santa Cruz Biotechnologies (Santa Cruz, California, USA), unless otherwise indicated. The antibody anti-cytokeratin (CK)-19 was purchased from Novocastra (Milan, Italy); the antibody anti-Bax was purchased from Cell-Signalling (Milan, Italy); Exendin-4 was purchased from American Peptide (Sunnyvale, California, USA). PD98059, Rp-cAMPS, KN62 and BAPTA/AM were purchased from Calbiochem (Milan, Italy). APO-ONE Homogeneous Caspase-3/7 Assay was purchased from Promega (Milan, Italy).
Experimental design
In vitro studies
Experiments were performed in cholangiocytes purified from normal rats, after a 1 h incubation at 37°C to regenerate membrane proteins damaged by proteolytic enzymes during the purification.10–12 During incubation, cells were kept in suspension in an RPMI medium,9 13 14 to which 5% fetal bovine serum (FBS) was added to limit the constitutive apoptosis.
To verify whether exendin-4 protects cholangiocytes from cell death by apoptosis, cholangiocytes were incubated for 4 h at 37°C13 with: (1) 0.2% bovine serum albumin (BSA; control); (2) glycochenodeoxycholic acid (GCDCA, 400 μmol/l),15 16 in the absence or presence of a 30 min preincubation with exendin-4 (100 nmol/l).9 To verify the eventual cytoprotective effects of exendin-4 against bile acids with different cytotoxicity, cells were incubated with taurochenodeoxycholic acid (TCDCA, 50 μmol/l),17–19 in the absence or presence of a preincubation with exendin-4, as above described.
To identify the intracellular mechanisms that mediate the antiapoptotic effects of exendin-4, the same experiments were also performed by preincubating cells for 30 min at 37°C with either Rp-cAMPs (100 μmol/l, a cAMP-dependent protein kinase A (PKA) inhibitor)9 13 or wortmannin (100 nmol/l, a phosphatidylinositol 3-kinase (PI3K) inhibitor).9 13 In addition, to establish whether Ca2+ signalling plays any role in mediating the cytoprotective effects of exendin-4, cells were also preincubated witheither BAPTA/AM (an intracellular Ca2+ chelator, 5 μmol/l)9 13 or KN62 (10 μmol/l, a calmodulin-dependent protein kinase II (CamKII) inhibitor).9 13
In vivo studies
Male Fischer rats (150–175 g), purchased from Charles River (Milan, Italy), were maintained in a temperature-controlled environment (20–22°C) with a 12 h light–dark cycle and with free access to drinking water and to standard rat chow.
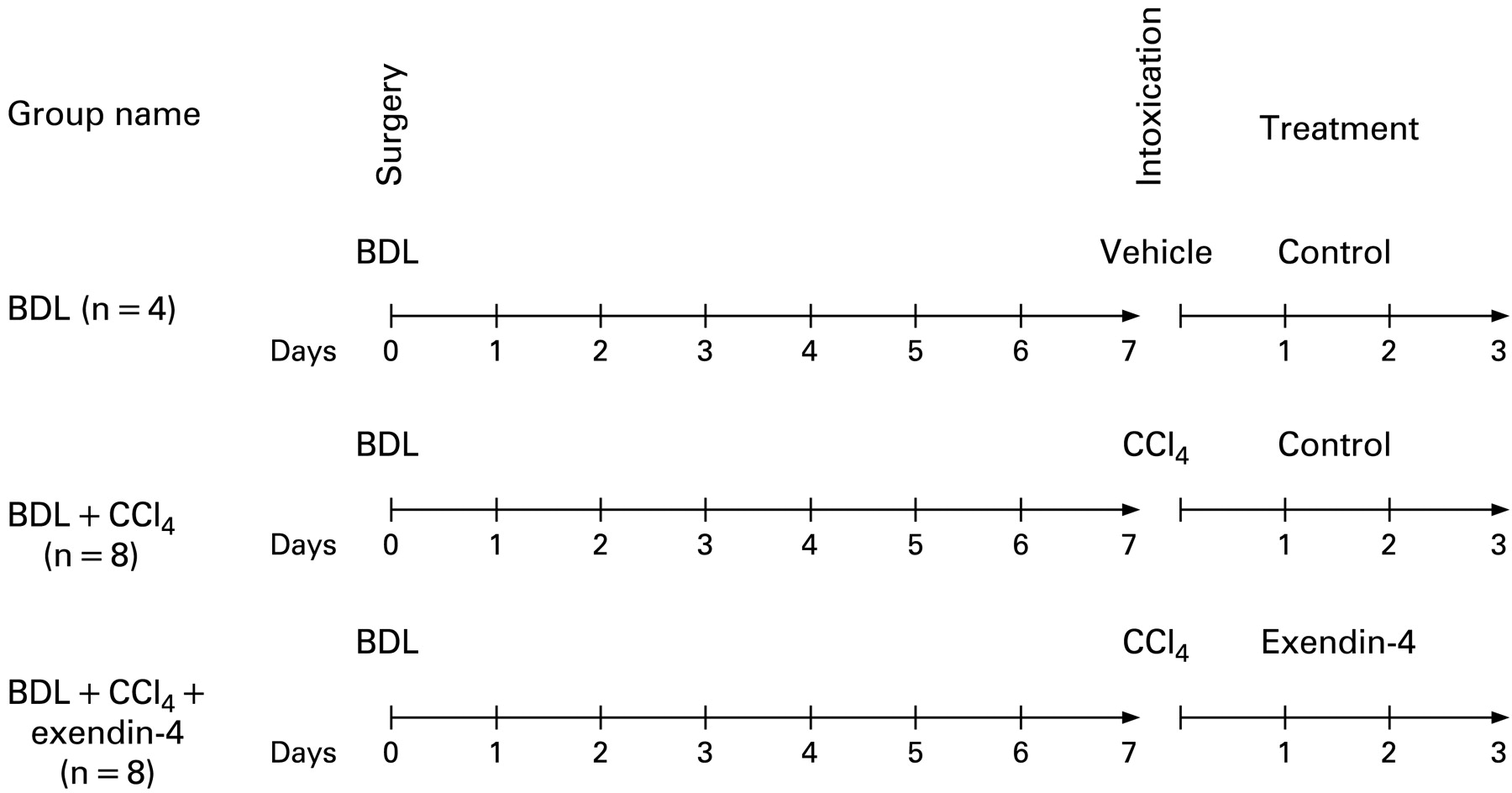
To study the effects of exendin-4 activation on cholangiocyte survival, our studies were performed in rats subjected to bile duct ligation (BDL). After 7 days, animals received vehicle or a single intraperitoneal injection of CCl4, 0.4 ml per 100 g body weight (50% mineral oil:CCl4), which triggers cholangiocyte cell death by apoptosis.20 Subsequently, animals were treated with either (1) exendin-4 (0.1 μg/kg body weight twice a day, intraperitoneally, n = 8) or (2) control injections (n = 8).9 21 Rats subjected to BDL, injected with vehicle and treated with control solution were used as internal controls (n = 4). No death in each experimental group was counted; at the end of the treatment, animals were sacrificed for liver sections. The treatment options and schedule are depicted in fig 1.
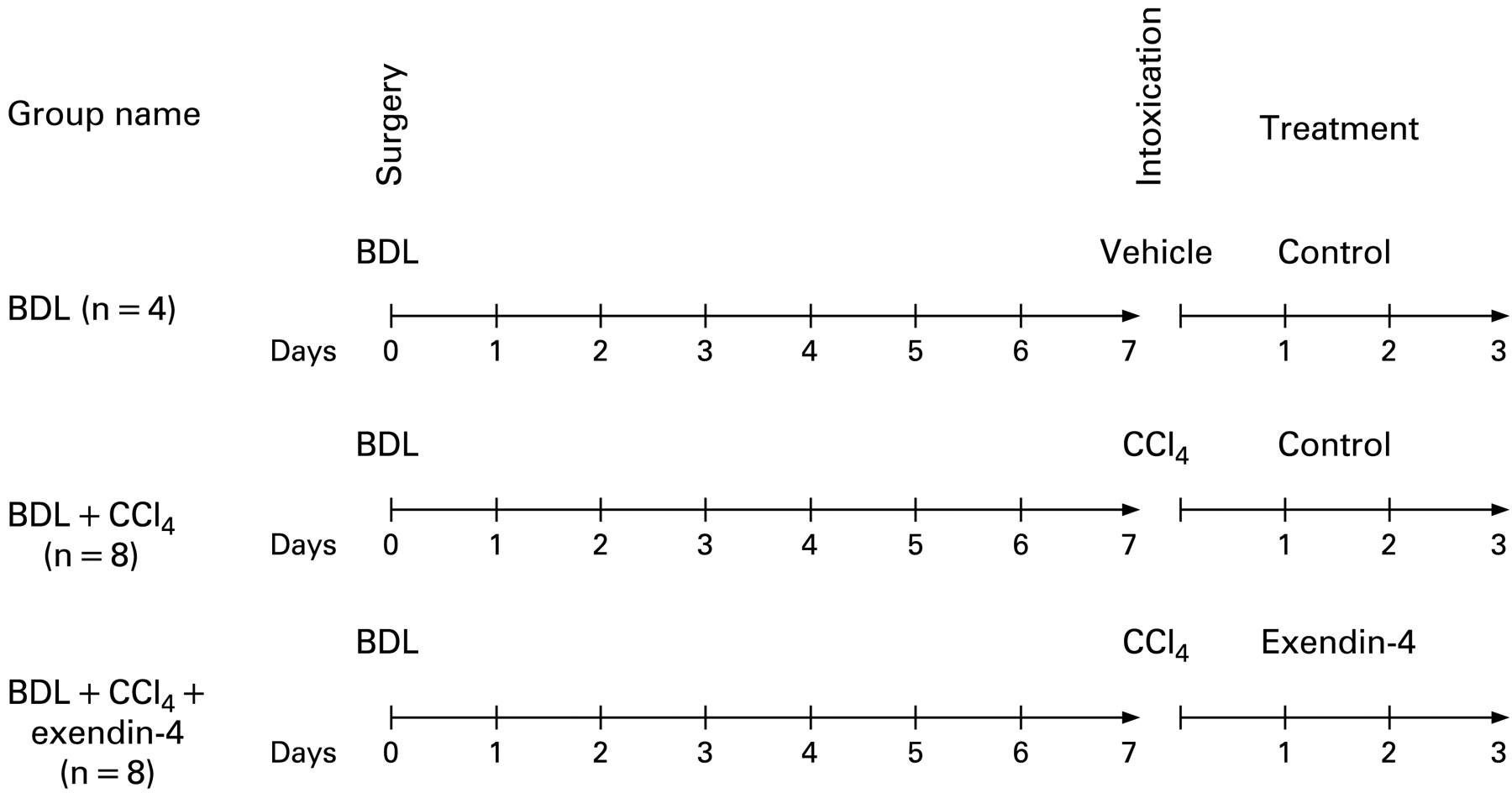
Outline of the treatment protocol of our in vivo model. All the animals were subjected to bild duct ligation (BDL) at day 0. After 7 days, the group named BDL + CCl4 received a single intraperitoneal injection of CCl4 and then received control injection treatment for 3 days, as in previous studies (n = 8).20 The group named BDL + CCl4 + exendin-4 were subjected to the same surgery and CCl4 intoxication, but were treated for 3 days with exendin-4 (0.1 μg/kg body weight twice a day, intraperitoneally, n = 8).9 The group named BDL employed, as in previous studies,20 as a control for CCl4 intoxication, received a single intraperitoneal injection of a control vehicle and, afterwards control injections for 3 days (n = 4).
The animals were fasted overnight before each experiment.9 13 Before each procedure, animals were anaesthetised with sodium pentobarbital (50 mg/kg intraperitoneally). Study protocols were performed in compliance with the institution guidelines.
Cholangiocyte purification and assessment of cell viability
Purification of cholangiocytes from rat liver was performed using a monoclonal antibody (immunoglobulin M (IgM), kindly provided by Dr R Faris, Brown University, Providence, Rhode Island, USA) against an unidentified membrane antigen expressed by all rat intrahepatic cholangiocytes.22 At the end of each procedure, the purity of the cholangiocytes was assessed by cytochemistry for γ-glutamyltranspeptidase (γ-GT).9 13 23 Cell viability at the end of the purification procedure was determined by trypan blue exclusion and was found to be >96%.
Caspase 3 activity
Changes in caspase 3 activation were measured by APO-ONE Homogeneous Caspase-3/7, according to the manufacturer’s instructions. Briefly, at the end of each experiment, 10 000 cells were resuspended with and incubated in a 1/100 dilution of the substrate Z-DEVD for 1 h. Fluorescence was measured by a 96-multiwell plate reader.
Subcellular fractionation and purification of mitochondria
Cholangiocytes were lysed with a Dounce homogeniser in 10 mmol/l HEPES/KOH (pH 7.6), 10 mmol/l KCl, 1 mmol/l MgCl2, 1 mmol/l dithiothreitol, containing aprotinin, 0.5 mmol/l phenylmethylsulfonyl fluoride and a complete protease inhibitor mixture. Immediately after homogenisation, sucrose was added to 250 mmol/l. Nuclei and unbroken cells were removed by centrifugation at 3000 g for 3 min, and the heavy membrane fraction (mitochondria rich) was sedimented by centrifugation at 9000 g for 20 min.24
Study of changes in Bax and cytochrome c expression and Akt phosphorylation
Proteins obtained from mitochondrial and cytosolic extracts (10 μg/lane) were resolved by sodium dodecylsulfate (SDS)–12% polyacrylamide gel electrophoresis (PAGE) and then transferred onto a nitrocellulose membrane. After blocking, membranes were incubated overnight at 4°C with either anti-Bax or anticytochrome c antibodies, followed by incubation with the corresponding secondary antibody. Similarly, proteins obtained from whole-cell lysates (10 μg/lane) were resolved by SDS–12% PAGE and then transferred onto a nitrocellulose membrane. Membranes were then incubated with antiphospho-Akt and anti-Akt, followed by incubation with the corresponding secondary antibody. Equal loading was evaluated by incubating membranes with the anti-β-actin antibody.25 26 Proteins were visualised using chemiluminescence (ECL Plus kit; Amersham, Milan, Italy). The intensity of the bands was determined by scanning video densitometry using the Chemi Doc imaging system (Bio Rad, Milan, Italy).
Assessment of changes in liver injury and morphology
Serum levels of alanine aminotransferase (ALT), alkaline phosphatase (ALP) and bilirubin were measured in rats from the different experimental groups, using commercially available kits (Sigma), as previously shown.20 27 Changes in liver injury and inflammation were assessed in liver sections after staining with H&E, as previously reported.28
In vivo changes in cell death by apoptosis were assessed by terminal deoxynucleotidyl transferase-mediated triphosphate end-labelling (TUNEL) analysis, as previously described (ApopTag Kit, Oncor, Gaithersburg, Maryland, USA).10 After counterstaining with haematoxylin solution, liver sections (four for each treatment group) were examined by light microscopy. Approximately 100 cells per slide were counted in a coded manner in seven non-overlapping fields.
Changes in bile duct mass were assessed by the computerised analysis of the immunohistochemistry for CK-19, as previously shown by us.9 13 14 20 After examination of the staining with a microscope, photographs of seven different fields per group (selected in a blinded, random fashion) were taken. The volume per cent of liver occupied by ducts was calculated from the total number of points over hepatic tissue and the number of points over CK-19-positive ducts, as previously reported.9 13 14 20
Statistical analysis
Data are expressed as means (SE). Data obtained from the in vitro experiments are expressed as a percentage of the basal value, with the exception of results of caspase 3 activity that are expressed as arbitrary units. The 95% CI was calculated. Differences between groups were analysed by analysis of variance (ANOVA). Differences between groups were considered significant when the p value was <0.05. We considered as principal indicators of induction of cell death by apoptosis: (1) caspase 3 activity for in vitro studies16 29 and (2) TUNEL staining for the in vivo studies.10 20 29–31 Changes in Bax mitochondrial expression and cytochrome c cytosolic expression were taken as indexes of activation of the “mitochondrial pathway” of induction of apoptosis.32
RESULTS
Exendin-4 protects cholangiocytes from cell death by apoptosis in vitro
Incubation with GCDCA induced apoptosis in cholangiocytes isolated from normal rats. GCDCA markedly increased the activity of caspase 3 compared with control (fig 2A; mean 16 672 (407), 95% CI 15 873 to 17 470). In comparison with untreated cells, GCDCA also enhanced Bax mitochondrial translocation and cytochrome c release, as suggested by increased expression of Bax in mitochondria (fig 2B; mean 257.06% (8.16%), 95% CI 241.04% to 273.07% of basal value) and cytochrome c in cytosol (fig 2C; mean 454.88% (45.87%), 95% CI 364.98% to 544.79% of basal value) fractions, respectively. Preincubation with exendin-4 limited the GCDCA-induced increases in caspase 3 activity (fig 2A; mean 3793 (800), 95% CI 2224 to 5361), Bax mitochondrial expression (fig 2B; mean 112.27% (11.84%), 95% CI 89.05% to 135.49% of basal value) and cytochrome c in cytosol (fig 2C; mean 227.76% (38.83%), 95% CI 151.65% to 303.86% of basal value).

In vitro effects of exendin-4 on cholangiocyte apoptosis. (A) Glycochenodeoxycholic acid (GCDCA; 400 μmol/l) induced a significant increase in caspase 3 activity that was prevented by preincubation with exendin-4 (100 nmol/l). Similarly, expression of Bax in mitochondria (B) and of cytochrome c (Cyt c) in cytosol (C) was markedly enhanced by GCDCA. Both those changes were prevented by preincubation with exendin-4. (D) Taurochenodeoxycholic acid (TCDCA; 50 μmol/l) slightly but not significantly increased caspase 3 activity compared with untreated cells. Data are the mean (SE) of at least three experiments. *p<0.05 vs basal; #p<0.05 vs GCDCA.
In contrast to GCDCA, TCDCA did not induce apoptosis in cholangiocytes, since it did not elicit any significant increase in caspase 3 activity (fig 2D; mean 5590 (1353), 95% CI 2937 to 8243).
The in vitro antiapoptotic effects of exendin-4 are mediated by the PI3K pathway
As shown in fig 3A, only preincubation with wortmannin (a PI3K inhibitor) neutralised the ability of exendin-4 to inhibit the activation of caspase 3 by GCDCA (GCDCA, mean 7047 (389), 95% CI 6285 to 7810; exendin-4+GCDCA, mean 3826 (834), 95% CI 2191 to 5461; wortmannin+exendin-4+GCDCA, mean 6406 (985), 95% CI 4475 to 8337). In contrast, no effects were observed when cells were pre-incubated with a cAMP-dependent PKA inhibitor (Rp-cAMPs), a CamKII inhibitor (KN62) or an intracellular Ca2+ chelator (BAPTA/AM).
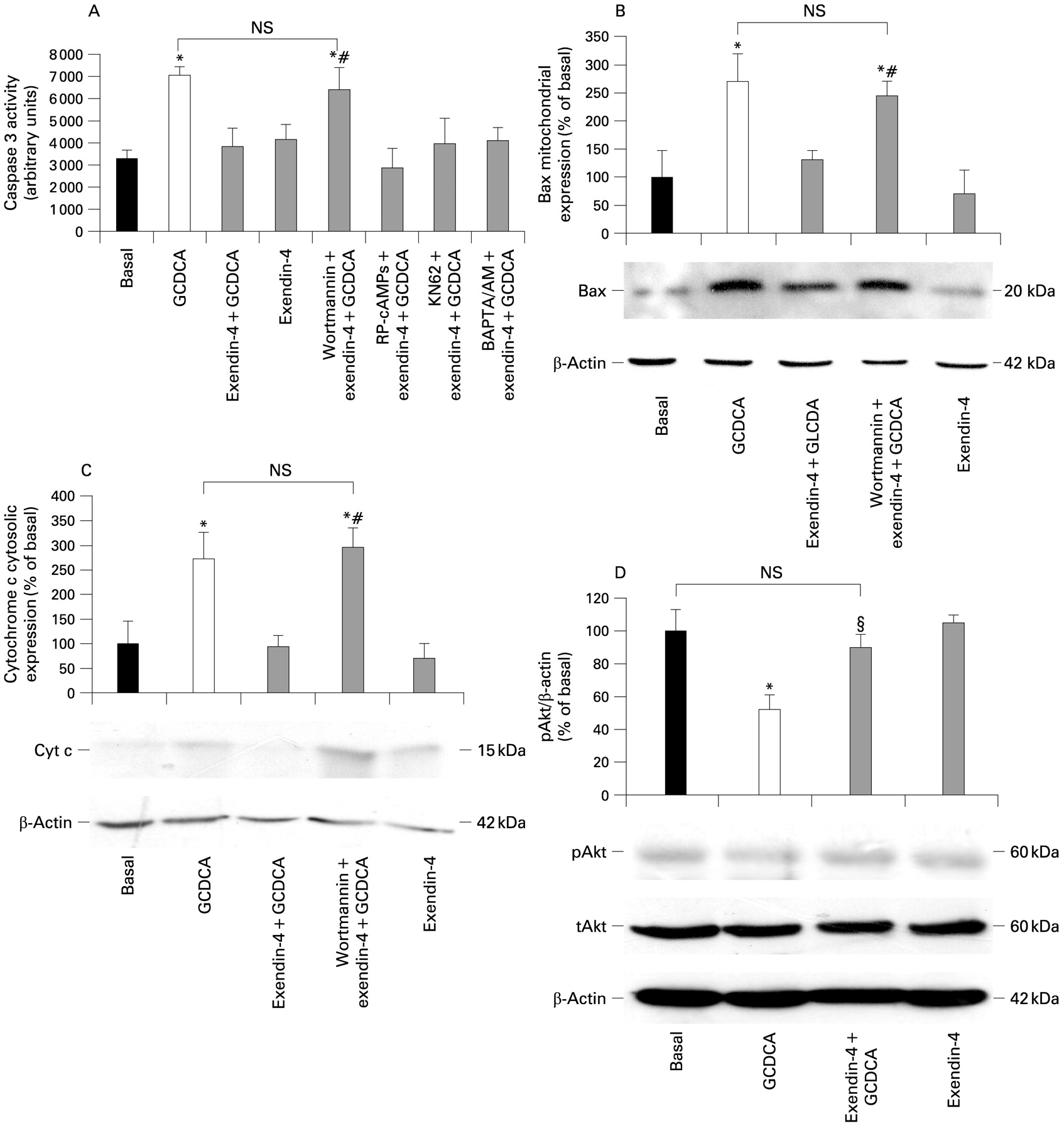
Intracellular signals mediating exendin-4 antiapoptotic effects on cholangiocytes. (A) Only the phosphatidylinositol 3-kinase (PI3K) inhibitor wortmannin (100 nmol/l) was able to neutralise the effect of exendin-4 (100 nmol/l) on the glycochenodeoxycholic acid- (GCDCA; 400 μmol/l) induced increase in caspase 3 activity. In contrast, no effects were observed when cells were preincubated with Rp-cAMPS (a cAMP-dependent protein kinase A inhibitor, 100 μmol/l), KN62 (a calmodulin-dependent protein kinase II inhibitor, 10 μmol/l) or BAPTA/AM (an intracellular Ca2+ chelator, 5 μmol/l). Similarly, wortmannin neutralised the effects of exendin-4 on the GCDCA-induced increase in Bax mitochondrial expression (B) and cytochrome c cytosolic expression (C). (D) Exendin-4 limited Akt dephosphorylation induced by GCDCA. Data are the mean (SE) of at least three experiments. *p<0.05 vs basal; #p<0.05 vs exendin-4 + GCDCA; §p<0.05 vs GCDCA; NS, not significant; pAKt, phosphorylated AKt; tAKt, total AKt.
The blockage of PI3K signalling by wortmannin also abolished the ability of exendin-4 to prevent the GCDCA-induced increase in Bax mitochondrial expression (fig 3B; GCDCA, mean 270.34% (49.84%), 95% CI 172.63% to 368.04%; exendin-4+GCDCA, mean 131.01% (17.45%), 95% CI 96.79% to 165.22%; wortmannin+exendin-4+GCDCA, mean 245.74% (24.87%), 95% CI 196.99% to 294.48% of basal). Similarly, wortmannin neutralised the effects of exendin-4 on the GCDCA-induced increase in cytochrome c expression in the cytosol (fig 3C; GCDCA, mean 272.84% (53.99%), 95% CI 167.00% to 378.67%; exendin-4+GCDCA, mean 94.51% (23.25%), 95% CI 47.92% to 139.09%; wortmannin+exendin-4+GCDCA, mean 296.13% (39.32%), 95% CI 219.04% to 373.21% of basal).
As a confirmation, cell preincubation with wortmannin prevented the reduction in Akt phosphorylation observed in cells exposed to GCDCA when compared with untreated cells (fig 3D: GCDCA, mean 51.97% (8.96%), 95% CI 34.40% to 69.55%; exendin-4+GCDCA: mean 89.87% (8.01%), 95% CI 95.86% to 114.14% of basal).
Exendin-4 ameliorates indexes of hepatocellular injury and cholestasis and reduces inflammation in an in vivo model of cholestasis and cell death
As previously shown,20 in vivo, a single CCl4 injection to rats whose had undergone BDL for 1 week produced a significant increase in serum levels of ALT, ALP and bilirubin, and in liver inflammatory infiltrate20 (table 1). In contrast, administration of exendin-4 to rats produced a significant reduction of levels of hepatocellular injury and cholestasis and of the degree of liver inflammation (table 1).
Exendin-4 prevents cholangiocyte apoptosis in an in vivo model of cholestasis and cell death
In vivo, a single CCl4 injection to rats subjected to BDL for 1 week induced cell death by apoptosis, as suggested by the relevant increase in the number of TUNEL-positive cells20 (BDL, mean 1.15 (0.29), 95% CI 0.58 to 1.71; BDL+CCl4, mean 6.51 (0.94), 95% CI 4.66 to 8.35). In contrast, administration of exendin-4 to rats produced a significant reduction of the apoptotic cholangiocytes (mean 2.31 (0.25), 95% CI 1.81 to 2.80; fig 4).
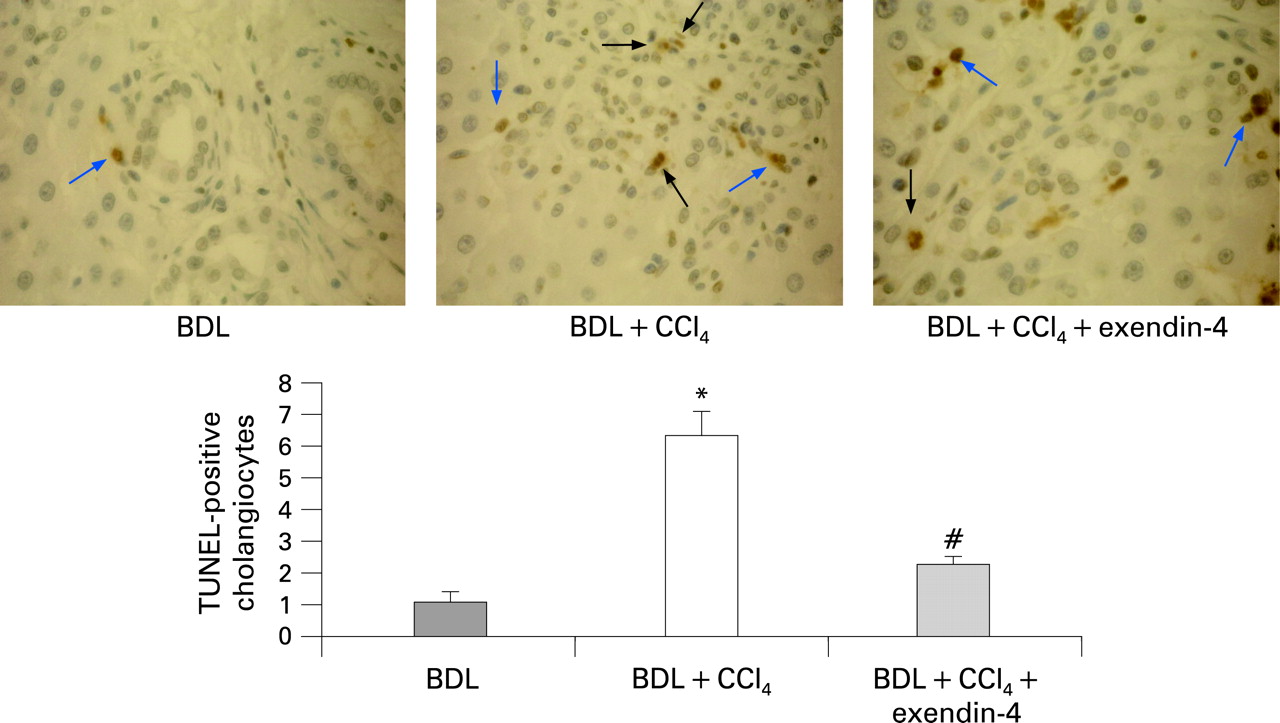
Effect of in vivo exendin-4 administration on cholangiocyte apoptosis. CCl4 intoxication induced cholangiocyte death in rats subjected to bile duct ligation (BDL); treatment with exendin-4 markedly reduced the number of terminal deoxynucleotidyl transferase-mediated triphosphate end-labelling (TUNEL)-positive cholangiocytes. Black arrows indicate cholangiocytes positive by TUNEL staining. Blue arrows indicate other cells positive by TUNEL staining (internal control). Four slices for each rat were considered; seven fields per slice were considered. *p<0.05 vs BDL; #p<0.05 vs BDL + CCl4.
Exendin-4 prevents the loss of bile ducts in an in vivo model of cholestasis and cell death
Loss of bile ducts was induced in vivo by a single CCl4 injection to rats subjected to BDL for 1 week, as witnessed by the strong reduction of the bile duct mass (BDL, mean 8.01 (1.12), 95% CI 5.81 to 10.22; BDL+CCl4: mean 3.36 (0.50), 95% CI 2.38 to 4.34). In contrast, administration of exendin-4 to rats prevented the CCL4-induced reduction of bile duct mass (mean 6.33 (0.78), 95% CI 4.79 to 7.87; fig 5).

Effect of in vivo exendin-4 administration on bile duct mass. CCl4 intoxication markedly diminished, in rats subjected to bild duct ligation (BDL), the bile duct mass, estimated by the computerised analysis of cytokeratin-19 staining. Exendin-4 treatment prevented the CCl4-induced loss of bile ducts. Four slices for each rat were considered; seven fields per slice were considered. *p<0.05 vs BDL; #p<0.05 vs BDL+CCl4.
DISCUSSION
The current study shows that exendin-4 protects the biliary epithelium from cell death by apoptosis, both in vivo and in vitro. Specifically, our study demonstrates that: (1) exendin-4 prevents cholangiocyte apoptosis induced, in vitro, by GCDCA; (2) the antiapoptotic effect of exendin-4 depends on PI3K inhibition of Bax mitochondrial translocation, cytochrome c release and caspase 3 activation; and (3) exendin-4 prevents cholangiocyte apoptosis and loss of bile ducts in an in vivo model of cholestasis and ductopenia.
Apoptosis is a highly organised type of cell death that ensures proper organogenesis and health of adult organs.32 For years, the dogma depicting apoptosis as an “innocuous” event has been accepted.33 Recent evidence suggests this concept to be true for “physiological” but not for “pathological” apoptosis.33 In physiological conditions, apoptosis is limited to a small number or subset of cells, both in number and over time.32 Pathological apoptosis involves a large number of cells in a non-selective fashion, is sustained over time and is typically associated with inflammatory conditions.32 Therapeutic modulation of apoptosis may represent a valid strategy for the treatment of several human diseases.32
Several liver diseases (including cholangiopathies) are considered to be due to dysregulation of cell survival.1 32 Cholangiopathies target cholangiocytes and progress as a result of enhanced apoptosis that prevails over compensatory proliferation, thus leading to ductopenia.1 In addition to that, in the course of primary biliary cirrhosis (PBC, the most common of the cholangiopathies),1 apoptotic cholangiocytes process some mitochondrial proteins differently compared with other cells.34 This amplifies the attraction of immune cells and enhances liver injury.34 Liver cell apoptosis is also thought to promote fibrogenesis,33 35 a typical feature of liver injury in late stage cholangiopathies.1 Hepatic stellate cells engulf apoptotic debris, an event that triggers their activation, for example the transdifferentiation towards a myofibroblast-like cell.33 35
Currently, there is no molecule known to be effective in maintaining an adequate survival of the biliary epithelium. In this study, we demonstrate that exendin-4 prevents cholangiocyte death by apoptosis, both in vitro and in vivo. In vitro, we found that the cytotoxic bile acid GCDCA but not TCDCA significantly induced apoptosis, in a similar fashion to what is observed in hepatocytes.15–17 25 Exendin-4 was able to prevent the induction of apoptosis by GCDCA (fig 2). In vivo, the administration of exendin-4 to rats subjected to BDL and CCl4 intoxication resulted in a significant reduction of TUNEL-positive cells and in the maintenance of the bile duct mass (figs 4 and 5). Thus, exendin-4 was found to be effective in two distinct models of cholangiocyte apoptosis, both reproducing important features of cholangiopathies, such as the overexposure to cytotoxic bile acids and the simultaneous presence of cholestasis and loss of bile ducts.1 20 34 36 To this extent, in its effects exendin-4 exhibited features similar to ursodeoxycholic acid (UDCA), the only compound that has a certain degree of clinical effectiveness at least in some cholangiopathies (such as PBC).1 34 We recently found UDCA to be antiapoptotic for cholangiocytes when tested in an in vivo model of cholestasis and bile duct loss.10 However, a specific antiapoptotic effect of UDCA on cholangiocytes in in vitro systems has never been demonstrated. In addition, the outcome of UDCA administration in vivo in different experimental set-ups resulted in heterogeneous changes in duct mass.10 37 UDCA may even amplify liver injury in the course of chronic cholestatic injury.38 In contrast, exendin-4 is effective in maintaining cholangiocyte proliferative and functional responses and duct mass in the course of cholestasis,9 as well as being beneficial even in other types of liver injury.39 Together, these findings suggest that exendin-4 has the potential to be effective in slowing down the progression of cholangiopathies, since it selectively acts on cholangiocytes, reversing the dysregulated balance between cell survival and growth.1
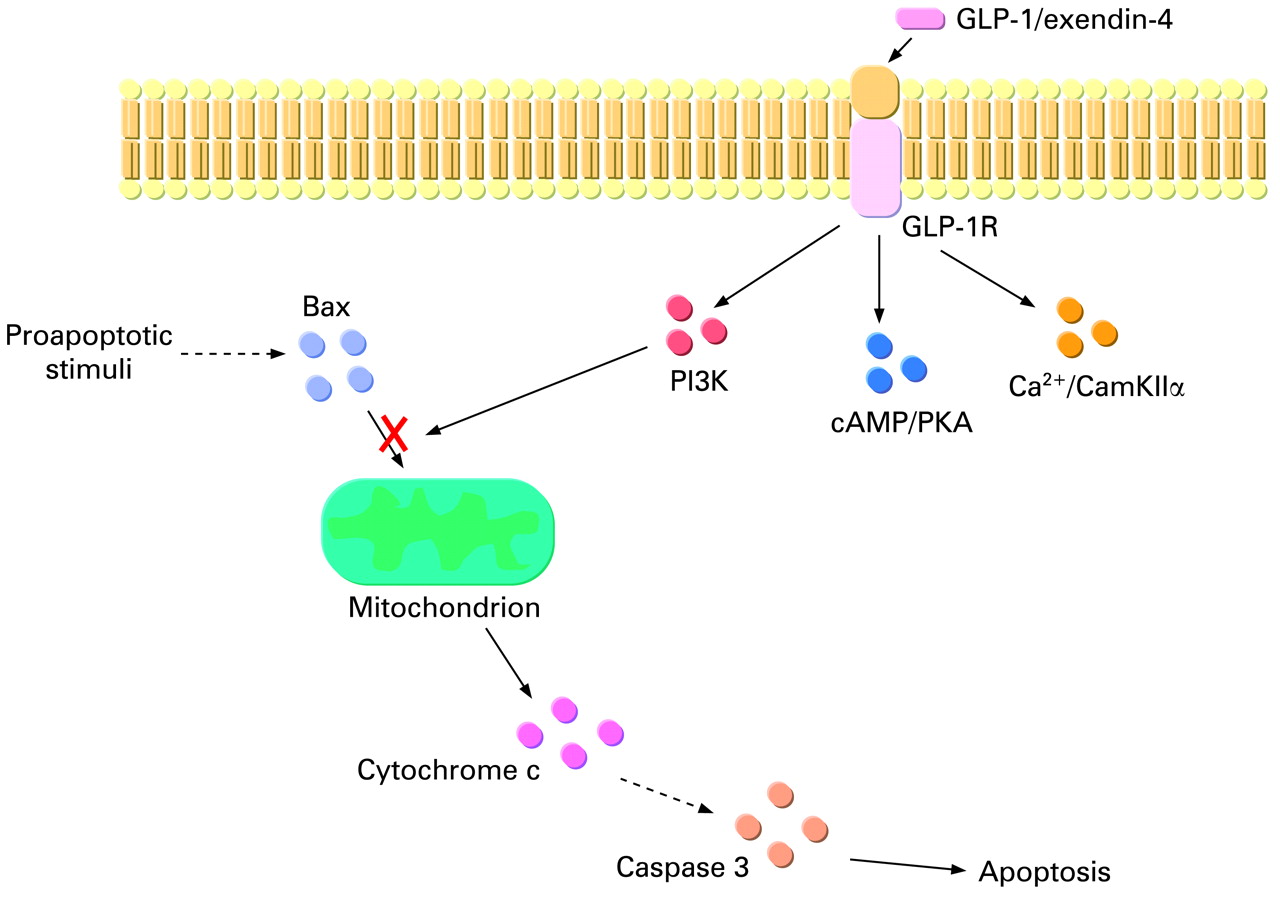
Apoptosis can occur substantially because of the activation of two defined molecular pathways, the extrinsic or intrinsic pathway.32 The extrinsic pathway is triggered at the plasma membrane following the activation of specific death receptors; the intrinsic pathway is triggered by different extracellular or intracellular stimuli that cause mitochondrial dysfunction.32 The activation of either one or the other pathway may depend on the kind of cell injury or on the cell type.32 However, the extrinsic and intrinsic pathways are not mutually exclusive32; in some cells, including hepatocytes and cholangiocytes, the death receptor pathway requires the mitochondrial signals to be amplified enough to deliver the proapoptotic message effectively.32 Thus, we wanted to investigate how exendin-4 affects the last steps of the apoptotic cascade in cholangiocytes. The in vitro incubation of cholangiocytes with GCDCA was associated with increased Bax expression in mitochondria and increased cytochrome c expression in cytosol (fig 2). Bax is a member of the family of Bcl-2 proteins, involved in the regulation of cell death.32 Upon the activation of the apoptotic machinery (either extrinsic or intrinsic), Bax migrates from the cytosol to mitochondria, where it causes membrane permeabilisation; as a consequence, cytochrome c is released from mitochondria into the cytosol, where it activates proteases mediators and effectors of programmed cell death, for example caspases.32 One of the main executor caspases is caspase 3. A similar sequence of events has also been shown to occur in hepatocytes upon incubation with GCDCA or other cytotoxic bile acids.25 40 41 Our in vitro system, therefore, reproduced the same cascade of events. Interestingly, when cells were preincubated with exendin-4 such a sequence of events was abolished (fig 2). These data suggest that exendin-4 is able to counteract the apoptotic machinery in cholangiocytes. In addition to that, we also showed that the antiapoptotic effects of GLP-1R activation are mediated by the activation of PI3K (fig 3). Indeed, blocking the PI3K pathway neutralised the effect of exendin-4 on Bax mitochondrial translocation and, as a consequence, cytochrome c release and caspase 3 activation. In several cell types, translocation of Bax can be modulated by PI3K signalling.42–44 As a confirmation, exendin-4 limited the dephosphorylation of Akt, which is immediately downstream of PI3K,45 induced by GCDCA. PI3K is a major determinant of cholangiocyte survival2 29 30 31 46 47; the current study strengthens such a concept, since amongst the different intracellular pathways that allow GLP-1R to enhance cholangiocyte proliferation (fig 6), PI3K is the only one to be involved in its antiapoptotic effects.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Proposed sequence of events that mediate the antiapoptotic effects of glucagon-like peptide-1 receptor (GLP-1R) activation by exendin-4 in cholangiocytes. GLP-1R enhances the activation state of phosphatidylinositol 3-kinase (PI3K), which reduces Bax translocation to mitochondria. As consequence, cytochrome c release from mitochondria to the cytosol and the consequent activation of the caspase cascade is reduced. The cAMP/protein kinase A (PKA) and Ca2+/calmodulin-dependent protein kinase II (CamKII) pathways, which mediate the proproliferative effects, do not participate in the antiapoptotic actions of GLP-1R activation.
Overall, our current findings are consistent with the action of GLP-1R activation on pancreatic β cell survival.4–6 GLP-1R is known to maintain β cell mass not only by enhancing cell proliferation but also by preventing apoptosis.4–6 In parallel to what we observed in cholangiocytes, the key molecule that mediates such a dual property of GLP-1R activation is PI3K.5 6 46 Interestingly, the administration of exendin-4 to Zucker diabetic rats also resulted in a reduction of the number of apoptotic pancreatic ductal cells, cells that share several features in common with cholangiocytes.48–51 A counteraction of apoptosis upon GLP-1R activation has also been observed in rat neurons.6 52
In summary, we demonstrated that activation of the GLP-1R in cholangiocytes by its selective agonist exendin-4 prevents cholangiocyte apoptosis. Overall, our findings suggest that exendin-4 is a molecule that is able to correct the dysregulated balance between cholangiocyte proliferation and death. Besides being relevant for the understanding of the pathophysiology of chronic cholestasis, our study supports exendin-4 trials in patients with cholangiopathies.4–7
REFERENCES
Footnotes
See Commentary, 902
Funding: This work was supported by MIUR grant 2005067975_004 to MM and by the Università Politecnica delle Marche intramural grants ATBEN00205 to AB and ATMAR01105 to MM; by a VA Merit Award, a VA Research Scholar Award, the Dr Nicholas C. Hightower Centennial Chair of Gastroenterology from Scott & White and NHS grants DK062975 and DK076898 to GA. The authors are grateful to Dr Paolo Onori, Department of Experimental Medicine, University of L’Aquila, L’Aquila, Italy, for his valued support in the histochemical studies.
Competing interests: None.