Article Text

Abstract
Background Although evidence points to a role for histamine and serotonin in visceral hypersensitivity, activation of calcium channels such as transient receptor potential vanilloid 4 (TRPV4) also causes visceral hypersensitivity. We hypothesised that TRPV4 is important for the generation of hypersensitivity, mediating histamine- and serotonin-induced visceral hypersensitivity.
Methods In response to histamine, serotonin and/or TRPV4 agonist (4αPDD), calcium signals and TRPV4 localisation studies were performed on dorsal root ganglia (DRG) neurons projecting from the colon. To evaluate visceral nociception, colorectal distension (CRD) was performed in mice treated with serotonin or histamine and with 4αPDD. Intrathecal injection of TRPV4 silencer RNA (SiRNA) or mismatch SiRNA was used to target TRPV4 expression.
Results Pre-exposure of DRG neurons projecting from the colon, to histamine or serotonin, increased Ca2+ responses induced by 4αPDD by a protein kinase C (PKC), phospholipase Cβ (PLCβ), mitogen-activated protein kinase kinase (MAPKK) and phospholipase A2 (PLA2)-dependent mechanisms. Serotonin or histamine treatments enhanced TRPV4 expression at the plasma membrane by a MAPKK mechanism. Hypersensitivity induced by serotonin or histamine were both significantly inhibited by TRPV4 SiRNA intrathecal injection. Administration of sub-nociceptive doses of serotonin or histamine potentiated 4αPDD-induced hypersensitivity in response to CRD.
Conclusions Serotonin and histamine sensitise TRPV4 response to 4αPDD both in vivo (increased visceral hypersensitivity) and in vitro, in sensory neurons, by a PKC, PLA2, PLCβ and MAPKK-dependent mechanism. Serotonin and histamine caused a MAPKK-dependent increase in TRPV4 expression in colonic sensory neurons plasma membranes. Further, histamine- or serotonin-mediated visceral hypersensitivity depend on TRPV4 expression in sensory neurons. TRPV4 appears as a common mechanism to several known mediators of visceral hypersensitivity.
- Visceral pain
- transient receptor potential vanilloid
- histamine
- serotonin
- sensory neurons
- summary box
- abdominal pain
- ION channels
- neurogastroenterology
- serotonin
Statistics from Altmetric.com
- Visceral pain
- transient receptor potential vanilloid
- histamine
- serotonin
- sensory neurons
- summary box
- abdominal pain
- ION channels
- neurogastroenterology
- serotonin
Introduction
Transient receptor potential vanilloid 4 (TRPV4) is a widely expressed cation channel of the transient receptor potential (TRP) superfamily. TRP channels are intrinsic membrane receptor channels with six transmembrane spans and a cation-permeable pore region.1 TRPV4 can be activated by physical stimuli such as cell swelling or innocuous warmth, and by chemical ligands.2 Among other cell types, TRPV4 has been shown to be expressed on sensory neurons, where it caused Ca2+ mobilisation, but also the release of nociceptive peptides such as substance P and calcitonin gene-related peptide (CGRP).3 Overall, different studies have defined a role for TRPV4 in the transduction of pain in somatic models,4–6 and, more recently, we have demonstrated that TRPV4 activation was a major component of visceral nociception.7 Further, we have shown that TRPV4 activation was a downstream signal of the visceral hypersensitivity symptoms induced by the activation of the protease-activated receptor-2 (PAR2), a G protein-coupled receptor that has been implicated in the peripheral generation of pain signals.8 Considering the recently demonstrated importance of protease signalling and PAR2 activation in pain symptoms related to inflammatory bowel syndrome (IBS),9 the fact that TRPV4 mediates PAR2-induced hypersensitivity in mouse models highlights TRPV4 as an important mechanism of a clinical feature common to patients with IBS: visceral hypersensitivity.7 One of the remaining important questions, though, is to determine whether TRPV4 activation is only a downstream effector specific of PAR2 signalling, or whether it could constitute a common mechanism to other mediators implicated in IBS-related visceral hypersensitivity. Among those mediators, histamine and serotonin occupy a predominant place, as they have been shown to be released by tissues from IBS patients.10 In addition, both mediators are implicated in visceral afferent firing and hyperexcitability of enteric neurons,10 highlighting the contribution of those mediators to visceral hypersensitivity symptoms. In the present study, we thus hypothesised that TRPV4 activation could be sensitised by serotonin and histamine, and that TRPV4 activation is a common downstream event of serotonin- and histamine-induced visceral hypersensitivity. By confirming those hypotheses, we showed that TRPV4 is a pivotal and common downstream event to several identified mediators of IBS-related visceral hypersensitivity symptoms. We further characterised the intracellular signalling pathways involved in the potentiation of TRPV4 signalling by serotonin and histamine.
Methods
Animals
C57Bl6 male mice (6–8 weeks) were purchased from Janvier (Le Genest Saint Isle, France).
Chemicals
Histamine, serotonin and TRPV4 agonist (4αPDD) were obtained from Sigma (Saint Quentin Fallavier, France) and dissolved in Hank's balanced salt solution (HBSS; Invitrogen, Cergy Pontoise, France) and 40% ethanol. SiRNA were synthesised by Core DNA Services of the University of Calgary and dissolved in Diethylpyrocarbonate (DEPC) water. GF109203X (a protein kinase C (PKC) inhibitor), H89 (a protein kinase A (PKA) inhibitor), PD98059 (a mitogen-activated protein kinase/extracellular signal-related kinase (MAPK/ERK) kinase inhibitor), U-73122 (a phospholipase C (PLC) inhibitor) and Methyl arachidonyl fluorophosphonate (MAFP)(a phospholipase A2 (PLA2) inhibitor) were obtained from Calbiochem (VWR, Fontenay sous Bois, France) and dissolved in HBSS/DMSO 0.5%. The H1 antagonist pyrilamine and the 5-HT3 antagonist ondansetron were obtained from Sigma and dissolved in HBSS.
Fast-Blue injection
Animals were anaesthetised with a xylazine/ketamine/NaCl 0.9% (1:20:29 ratio, respectively; intraperitoneally) combination. The abdomen was opened by midline laparotomy, and the colon was gently exteriorised. Small volumes (1–2 μl) of the retrograde tracer Fast Blue (FB) (1 mg in 80 μl of saline; Cedarlane Laboratories, Burlington, Ontario, Canada) were injected into the wall of the colon. Five injections of 3 μl were made on both sides of the colon using a 30-gauge needle fitted to a Hamilton syringe (total volume 15 μl). The exterior of the colon was swabbed after each injection to remove residual tracer to avoid erroneous labelling.
Isolation of dorsal root ganglia neurons
Dorsal root ganglia (DRG) from T7-L2 and L5-S1 were rinsed in cold HBSS, and enzymatically dissociated as described previously.11 Neurons in the pellet were suspended in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) containing 2.5% fetal bovine serum, 2 mmol/l glutamine, 1 mg/ml penicillin/streptomycin and 10 μmol/l each of cytosine arabinoside (ARAC), Floxuridine (FUDR), uridine (all from Sigma). Cells were plated in CC2 LabTek II (Nunc, Domique Dutscher, Brumath, France) for calcium signalling assay or in LabTeck II (Nunc) coated with poly-l-ornithine/laminin (Sigma) for immunohistochemistry.
Calcium imaging
HEK-FLPTREX-TRPV4 (HEK-FLPTREX cells stably transfected with pcDNA5/FRT/TO+hTRPV4) express the TRPV4 protein on a tetracycline-dependent promoter.3 Cells were grown in DMEM containing 10% tetracycline-free fetal bovine serum, hygromycin (50 μg/ml) and blasticidin (10 μg/ml) in 96-well plates. To induce TRPV4 expression, tetracycline (0.1 μg/ml) was added to the medium 16 h before use.3 HEK-FLPTREX-TRPV4 or neurons were incubated for 30 min at 37°C in HBSS–BSA (Sigma) solution containing 2.5 mmol/l probenecid (Sigma), pH 7.45 supplemented with fluo-3 acetoxymethyl (AM) 1 mmol/l (Invitrogen) and 20% pluronic F-17 (Sigma). After the incubation period, cultures were placed into a 37°C incubator in the dark for 30 min. The fluorescence was measured at 460–490 nm excitation and 515 nm emission in each well. Data of HEK-FLPTREX-TRPV4 were collected and analysed with the Novostar software (BMG Labtech, Champigny s/Marne, France). Neurons were imaged using an inverted microscope (Zeiss, Le Pecq, France) and a ×10 objective. Images were acquired using Metamorph software (Molecular Devices, Evry, France). For both cell types, acquisition parameters were kept constant within each experiment. A kinetic of 200 recordings was performed and free intracellular calcium concentration quantified as described previously.11 The percentage of responding neurons was based on six independent experiments with 14–16 neurons analysed per experiment. In a first set of experiment neurons were pre-treated with serotonin (10, 50 or 100 μmol/l), histamine (10, 50 or 100 μmol/l) or vehicle for 5 min and treated with 4αPDD (50 μmol/l). HEK-FLPTREX-TRPV4 were treated with serotonin (100 μmol/l), histamine (100 μmol/l), 4αPDD (100 μmol/l) or vehicle. In a second set of experiments, neurons were pre-incubated for 30 min with inhibitors: PKC(3), PLC(3), PKA(3), MAPK/ERK kinase,12 PLA213 or vehicle (10 μmol/l each). In a final set of experiments, neurons were pre-incubated for 30 min with the 5-HT3 antagonist ondansetron, the H1 antagonist pyrilamine or their vehicle (10 μmol/l each).
Immunohistochemistry
DRG neurons were isolated from mice and cultured as described above. Neurons were washed in phosphate-buffered saline (PBS) containing 1% bovine serum albumin (BSA) and then incubated for 20 min in 4% paraformaldehyde. This step was followed by incubation (5 min) with PBS, 1% BSA and 0.05% Triton X-100. After three washes of 10 min each (in PBS, BSA 1%), cells were incubated with primary antibodies against TRPV4 (rabbit, 1:100; AbCam, TEBU Bio, Le Perray en Yvelines, France) overnight at 4°C. Neurons were washed and incubated with secondary antibodies conjugated to Alexa Fluor 488 (1:1000; Molecular Probes, Invitrogen) at room temperature for 2 h. After two washes, staining of the nuclei was performed by incubation of the neurons with TO-PRO3 (1/1000; Invitrogen) for 10 min at room temperature. As a control for antibody specificity, the primary antiserum was pre-incubated with the peptide used for immunisation for 24 h at 4°C before staining. Confocal images were acquired using a Zeiss LSM-510 META confocal microscope using a ×40 objective. Images were processed to obtain a three-dimensional reconstruction. For each neuron, the area positively stained for TRPV4 was measured with Metamorph software, at the plasma membrane, and in perinuclear and nuclear zones (TO-PRO3 labelled). The results are expressed as the ratio between the area of fluorescence observed at the nuclear level and the area of fluorescence observed at the membrane. For calcium imaging, neurons were pre-treated for 30 min with inhibitors (10 μmol/l) or vehicle and with serotonin (100 μmol/l), histamine (100 μmol/) or buffer.
Colorectal distension and electromyography recordings
Groups of electrodes were implanted in the abdominal external oblique musculature of anaesthetised mice as previously described.9 Colorectal distension (CRD) was performed 5 days after the electrode implantation9 by insertion of a balloon (10 mm long) into the colon at 5 mm from the anus. The balloon was then inflated in a stepwise fashion from 0 to 60 mm Hg with 15 mm Hg increments. Ten-second distensions were performed in triplicate at pressures of 15, 30, 45 and 60 mm Hg with 5-min intervals as previously described.
In a first set of experiments, mice received intracolonic injection of 100 μl of histamine or serotonin (5, 10, 25 and 100 μg/mice). At various times after administration (0.5, 2, 4, 6 h), we performed series of CRD. Kinetic and dose–response studies were performed. In a second set of experiments, anaesthetised mice (halothane 5%) received three inter-vertebral injections (10 μl over 36 h, every 12 h) by subcutaneous inter-vertebral injection between L5 and L6 of TRPV4 siRNA: 6-FAM (6-carboxyfluorescein) 5′-UCUACCAGUACUAUGGCUUd(TT)-3′; 3′-d(TT)AGAUGGUCAUGAUACCGAA-5′ or a mismatched siRNA designed with the same percentage of GC and AT but with no corresponding sequence: 6-FAM 5′-CAUGCUAGGUUAGUACUUGd(TT)-3′; 3′-d(TT)GUACGAUCCAAUCAUGAAC-5′;7 these mice received an intracolonic dose of serotonin (10 μg/mice, 100μl), or histamine (25 μg/mice, 100 μl) causing hypersensitivity, or their vehicle (40% ethanol); CRD was performed 2 h later. In a third set of experiments, we used eight groups of eight mice, administrated with histamine (10 μg/mice, 100 μl), serotonin (5 μg/mice, 100 μl) or their vehicle (40% ethanol) and 1 h later with a dose of 4αPDD (7 μg/mice, 100 μl) not causing hypersensitivity, CRD was performed 1 h after TRPV4 agonist treatment.
Statistical analysis
Data are presented as mean±SEM. Analyses were done by running the GraphPad Prism 4.0 software. Between-group comparisons were performed by the unpaired t test. Multiple comparisons within groups were performed by repeated-measures one-way ANOVA, followed by Tukey's post test. Statistical significance was accepted at p<0.05.
Results
Effects of histamine and serotonin on the calcium response to TRPV4 agonist in colonic sensory afferents
Colonic sensory afferents were labelled by injecting into the colonic wall of mice with the retrograde marker Fast Blue. Blue-labelled neurons isolated from those mouse DRGs were then exposed the TRPV4 agonist 4αPDD and Ca2+ responses were recorded from those neurons. 4αPDD (50 μmol/l) caused an increased intracellular concentration of Ca2+, which was potentiated by pre-exposure of those neurons for 5 min to serotonin or histamine (50 or 100 μmol/l each), compared to vehicle exposure (figure 1A). In addition, a significant increase in the number of neurons responding to 4αPDD following pre-exposure to histamine or serotonin was observed (only at the dose of 100 μmol/l), compared to vehicle pre-exposure (48% vs 31% for histamine and 43% vs 31% for serotonin) (figure 1B). To assess the possibility of a direct activation of TRPV4 by serotonin or histamine, HEK cells transfected with TRPV4 (HEK-FLPTREX-TRPV4) were exposed to serotonin or histamine (100 μmol/l). Neither histamine nor serotonin was able to induce a calcium flux in those cells (figure 1C). Taken together, these results showed that serotonin or histamine sensitises cellular response to TRPV4 agonists in colonic primary afferents.

Serotonin and histamine-induced sensitisation of TRPV4 Ca2+ signals. Neurons were pre-treated for 5 min with histamine (10, 50 or 100 μmol/l), serotonin (10, 50 or 100 μmol/l) or vehicle before being challenged with 4αPDD (50 μmol/l). [Ca2+]i quantification in mouse DRG neurons projecting from the colon challenged with 4αPDD (50 μm) (A). Proportion of neurons that responded to 4αPDD (50 μmol/l) by increasing [Ca2+]i (B). *p<0.05 compared to buffer /4αPDD group; ANOVA followed by Tukey's test; n=45–49 neurons. (C) Calcium response to 4αPDD (100 μmol/l), serotonin (100 μmol/l), histamine (100 μmol/l) or their vehicle in HEK-FLPTREX-TRPV4 treated (black bar) or not (white bar) with tetracycline. **p<0.01 compared to – tetracycline corresponding group. DMSO, dimethyl sulfoxide; DRG, dorsal root ganglia; HBSS, Hank's balanced salt solution; TRPV4, transient receptor potential vanilloid 4.
Intracellular mechanisms involved in the potentiation of TRPV4 response by histamine or serotonin in primary afferents
The inhibitor of PKA (H-89; 10 μmol/l) did not modify the potentiated TRPV4 Ca2+ signalling in DRG neurons after serotonin (figure 2A) or histamine (figure 3A) pre-exposure, nor did it modify the number of neurons responding to TRPV4 activation in the same conditions (figures 2B and 3B). In contrast, inhibitors of PKCs (GF109203X; blocks PKCα, β, γ, δ, ɛ and ζ; 10 μmol/l), of MAPK/ERK kinase (PD98059; 10 μmol/l), of PLCβ (U73122; 10 μmol/l) and inhibitors of PLA2 (MAFP; 10 μmol/l) inhibited serotonin- or histamine-induced potentiation of Ca2+ responses to 4αPDD (figures 2A and 3A). Similarly, the increased number of responding neurons to the TRPV4 agonist 4αPDD after serotonin or histamine exposure was inhibited by those inhibitors (figures 2B and 3B). Calcium mobilisation response to 4αPDD in DRG neurons was not modified by individual incubations with each of those inhibitors (data not shown). Moreover, none of the inhibitors were able to modify the minimal or the maximal calcium responses in the neurons, demonstrating the lack of effect of these inhibitors on sensory neurons calcium states (data not shown).
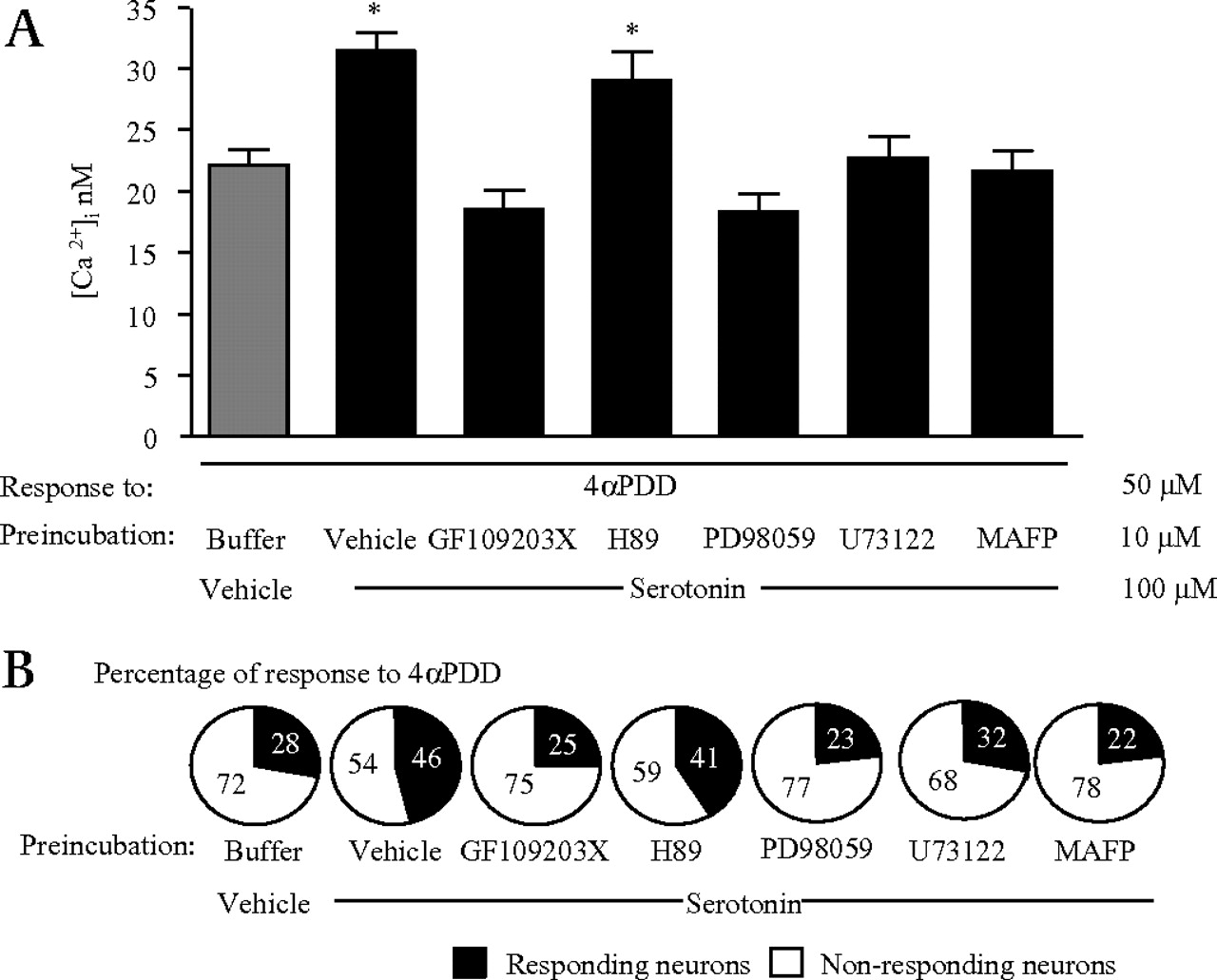
Mechanisms of serotonin-induced sensitisation of transient receptor potential vanilloid 4 (TRPV4) Ca2+ signals. Effects of 30 min incubation with signalling pathways inhibitors (GF109203X, H89, PD98059, U73122, MAFP, all 10 μmol/l) on serotonin-induced sensitisation of Ca2+ signals to 4αPDD (50 μmol/l) in sensory neurons (A) and on the percentage of responding neurons (B). Cells were pre-treated with serotonin or vehicle (100 μmol/l) for 5 min before being challenged with TRPV4 agonists. *p<0.05 compared to vehicle /buffer /4αPDD group; ANOVA followed by Tukey's test; n=76–95 neurons.

Mechanisms of histamine-induced sensitisation of transient receptor potential vanilloid 4 (TRPV4) Ca2+ signals. Effects of 30 min incubation with signalling pathways inhibitors (GF109203X, H89, PD98059, U73122, MAFP, all 10 μmol/l) on histamine-induced sensitisation of Ca2+ signals to 4αPDD (50 μmol/l) in sensory neurons (A) and on the percentage of responding neurons (B). Cells were pre-treated with histamine or vehicle (100 μmol/l) for 5 min before being challenged with TRPV4 agonists. *p<0.05 compared to vehicle /buffer /4αPDD group; ANOVA followed by Tukey's test; n=76–95 neurons.
Effects of H1 and 5-HT3 antagonist treatment on the potentiation of TRPV4 response by histamine or serotonin in primary afferents
Treatment with the H1 receptor antagonist pyrilamide decreased histamine-induced potentiation of Ca2+ responses to 4αPDD (figure 4A). However, the increased number of responding neurons to the TRPV4 agonist 4αPDD after histamine exposure was unchanged by pyrilamide (figure 4B). The increased number of responding neurons to the TRPV4 agonist 4αPDD after serotonin exposure was inhibited by pretreatment with the 5-HT3 receptor antagonist ondansetron (figure 4B). In addition, pretreatment with ondansetron decreased serotonin-induced potentiation of Ca2+ responses to 4αPDD (figure 4A). Calcium mobilisation response to 4αPDD in DRG neurons was not modified by individual incubations with each of those antagonists (data not shown).

Implication of the H1 and 5-HT3 receptors in histamine or serotonin-induced sensitisation of transient receptor potential vanilloid 4 (TRPV4) Ca2+ signals. Effects of 30 min incubation with H1 or 5-HT3 receptor antagonists (pyrilamine and ondansetron, respectively; 10 μmol/l) on serotonin- or histamine-induced sensitisation of Ca2+ signals to 4αPDD (50 μmol/l) in sensory neurons (A) and on the percentage of responding neurons (B). Cells were pre-treated with histamine or vehicle (100 μmol/l) for 5 min before being challenged with TRPV4 agonists. *p<0.05 compared to vehicle/buffer /4αPDD group, #p<0.05 compared to vehicle/serotonine or histamine/4αPDD group; ANOVA followed by Tukey's test; n=55–64 neurons.
Effects of histamine and serotonin treatment on TRPV4 localisation in primary afferents
In basal conditions (no specific incubations), we observed that TRPV4 was localised both at the plasma membrane and in the nuclear and perinuclear area of primary afferents isolated from mouse DRGs (figure 5A,B). TRPV4 fluorescent labelling was higher in the nuclear and perinuclear zones (co-staining with TO-PRO nuclear labelling) than at the plasma membrane surface in the control (buffer exposure), with a ratio nuclear and perinuclear zones/membrane of 1.19±0.005 (figure 5A,B). Exposure of DRG neurons to histamine or serotonin decreased the ratio nuclei/plasma membrane to values of 0.65±0.003 vs 1.21±0.010 and 0.62±0.004 vs 1.19±0.005 for histamine and serotonin pre-exposure respectively (figure 5A,B). In addition, it was clear that after histamine or serotonin pre-exposure, TRPV4 staining became visible in neurites and not only in cell bodies. PD98059 (10 μmol/l) inhibited serotonin and histamine-induced TRPV4 plasma membrane localisation (figure 5A,B). However, inhibitors (10 μmol/l) of PLCβ, PKA, PKCs and cPLA2 had no effect on serotonin or histamine-induced TRPV4 plasma membrane localisation (figure 5A,B).

Immunostaining of transient receptor potential vanilloid 4 (TRPV4) in sensory neurons from dorsal root ganglia (DRG). Quantification and TRPV4 labelling (in green) and TOPRO-3 fluorescence (in blue) on three-dimensional reconstruction in Z-plan images of cultured sensory neurons treated with serotonin (100 μmol/l; A) or histamine (100 μmol/l; B) or their vehicle and pre-treated during 30 min with GF109203X (10 μmol/l), PD98059 (10 μmol/l), U73122 (10 μmol/l), Methyl arachidonyl fluorophosphonate (MAFP; 10 μmol/l) or H-89 (10 μmol/l). *p<0.05 compared to control; ANOVA followed by Tukey's test, n=30–36 neurons.
Effects of intracolonic administration of histamine or serotonin on visceral nociception
Visceral motor responses (VMRs) to CRD of mice that received intracolonic administration of saline or vehicle (40% ethanol in saline) were similar (not shown). The intracolonic administration of histamine (25 and 100 μg/mice; 100 μl) or serotonin (10, 25 and 100 μg/mice; 100 μl), provoked a dose-dependent increase in VMR to CRD for all of distension pressure (figure 6A), 2 h after their administration. Serotonin (10, 25 and 100 μg/mice; 100 μl) and histamine (25 and 100 μg/mice; 100 μl) provoked allodynia, characterised by an increased VMR to 15 mm Hg, a normally innocuous pressure of distension (figure 6A) and hyperalgesia characterised by an increased VMR to noxious pressures (30, 45 and 60 mm Hg) of distension (figure 6A). Allodynia and hyperalgesia following serotonin (10 μg/mice; 100 μl) or histamine (25 μg/mice; 100 μl) treatments were observed from 1 to 4 h with a maximum at 2 h after their administration (figure 6B).

Intracolonic administration of serotonin or histamine induced visceral hypersentivity in response to colorectal distention (CRD). Dose–response curves of visceral motor response (VMR) induced by the intracolonic administration of serotonin (A, left panel) or histamine (A, right panel) (10, 25 or 100 μg/mice; 100 μl) in response to CRD (15, 30, 45 and 60 mm Hg). Mice received an intracolonic administration of serotonin (10 μg/mice; 100 μl; B, left panel) or histamine (25 μg/mice; 100 μl; B, right panel) and were submitted to CRD 1, 2, 4 and 6 h after the intracolonic injection. VMR to CRD was recorded in response to pressure of distension or 15, 30, 45 and 60 mm Hg. Time 0 corresponds to VMR recorded in response to CRD, before serotonin or histamine administration. Values are mean+SEM, n=5 mice per group. *p<0.05 compared with 40% ethanol-injected group.
Effects of TRPV4 blockade on histamine or serotonin-induced allodynia and hyperalgesia
We have previously determined that 3 days after inter-vertebral injection of TRPV4-targeted siRNA, the inhibition of TRPV4-induced visceral hypersensitivity was maximal7 and coupled with a loss of TRPV4 expression in the dorsal root ganglia neurons. Therefore, we used the same time-point of 3 days after inter-vertebral injection of TRPV4− or mismatched siRNA for the following functional studies. The hyperalgesia observed 2 h after intracolonic administration of serotonin (10 μg/mice; 100 μl) or histamine (25 μg/mice; 100 μl) was prevented by an inter-vertebral injection of TRPV4-directed siRNA, 3 days before the experiment (figure 7A). Allodynia observed 2 h after intracolonic injection of serotonin (10 μg/mice; 100 μl) was prevented by an inter-vertebral injection of TRPV4-directed siRNA, 3 days before the experiment, but histamine (25 μg/mice; 100 μl)-induced allodynia was not prevented by the same treatment (figure 7A). The inter-vertebral injection of the mismatched siRNA 3 days before CRD had no effect on allodynia and hyperalgesia induced by serotonin or histamine, as compared to values obtained after control distension (figure 7A).

Role of TRPV4 DRG neuron expression in histamine and serotonin-induced visceral hypersensitity. Mice received inter-vertebral administration of siRNA control (white squares and black triangles) or directed against TRPV4 (grey inverted triangles), 3 days before the intracolonic administration of serotonin (10 μg/mice; 100 μl, black triangles and grey triangles) or histamine (25 μg/mice; 100 μl, black and grey triangles). Two hours after the intracolonic administration, CRD was performed and visceromotor response was recorded. Values are mean±SEM, n=8 mice. *p<0.05 significantly different from control values (A). Mice received intracolonic administration of histamine or serotonin (10 and 5 μg/mice; 100 μl respectively, grey inverted triangles and black circles) or its vehicle (40% ethanol, white square and grey triangles). One hour later, mice received intracolonically 4αPDD (7 μg/mice; 100 μl, black circles and grey triangles) or its vehicle (40% ethanol, grey inverted triangles and white squares). CRD were performed 1 h after 4αPDD administration and VMR were recorded. Values are mean±SEM, n=8 mice per group. *p<0.05, significantly different from 40% ethanol group (B). CRD, colorectal distention; DRG, dorsal root ganglia; TRPV4, transient receptor potential vanilloid 4.
We investigated the possibility that histamine and serotonin could sensitise TRPV4-induced visceral hypersensitivity, by co-administering doses of histamine or serotonin and 4αPDD that individually did not cause hypersensitivity. Intracolonic administration of 10 μmol/l of 4αPDD, 5 μg/mice; 100 μl of serotonin or 10 μg/mice; 100 μl of histamine did not cause visceral allodynia and hyperalgesia (figure 7B). However, intracolonic pre-treatment of mice with serotonin (5 μg/mice; 100 μl) provoked allodynia and hyperalgesia, 1 h after the intracolonic administration of a non-nociceptive dose of 4αPDD (7 μg/mice; 100 μl) (figure 7B). Histamine pre-treatment (intracolonic 10 μg/mice; 100 μl) did not favour allodynia in mice that received a non-nociceptive dose of 4αPDD (7 μg/mice; 100 μl), but induced hyperalgesia in those mice (figure 7B). Taken together, those data provide evidence that histamine and serotonin exposure potentiate TRPV4-induced visceral hyperalgesia, and that serotonin, but not histamine, potentiates TRPV4-induced visceral allodynia.
Discussion
Patients with functional disorders such as IBS sense pain and discomfort in the absence of any noxious stimuli, tissue damage or clearly defined pathological changes of tissue homeostasis. A number of studies are now providing evidence that peripheral dysfunctions and peripherally released mediators could be implicated in the aetiology of IBS.14 The present study provides more evidence of the potential implication of peripheral mediators in visceral hypersensitivity states. We demonstrated here that serotonin and histamine, two mediators that have been shown to be released in colonic tissues from patients with IBS,10 were able to sensitise responses of the calcium channel TRPV4 in vivo and in peripheral sensory neurons, thereby participating to the generation of hypersensitivity symptoms.
Barbara et al have demonstrated that histamine released from mast cells in biopsy supernatants from patients with IBS was able to increase intracellular calcium concentration in rat primary afferents suggesting that histamine plays an important role in visceral afferent sensitisation.10 Along the same lines, our results also support the role of histamine in visceral hypersensitivity, as we demonstrated that intracolonic administration of histamine causes visceral allodynia and hyperalgesia. In addition, we characterised the role of TRPV4 in histamine-induced visceral hypersensitivity, as mice injected TRPV4-targeted SiRNA, did not develop visceral hyperalgesia in response to histamine. Interestingly, histamine-induced allodynia did not seem to depend on TRPV4 expression on sensory neurons, as visceral motor response to 15 mm Hg pressure of distention after histamine intracolonic administration was not modified by TRPV4 SiRNA injection, nor was induced allodynia by combining histamine and TRPV4 agonist treatments. The relationships between histamine and members of the TRPV family in vivo have been well described for TRPV1.15 In that study, the authors propose a PLA2/lipoxygenase/TRPV1 pathway in sensory neurons for the scratching induced by histamine in vivo.15 From our results, we can propose that TRPV4 is implicated in histamine-induced visceral hyperalgesia and that TRPV4 sensitisation by histamine on peripheral sensory afferents could be responsible for histamine-mediated visceral hypersensitivity.
Increased numbers of rectal enterochromaffin cells and an enhanced peak of postprandial serotonin release have been reported to occur in patients with post-infectious IBS.16 17 Serotonin stimulates vagal and intrinsic afferent nerve fibres18 and has been shown to be a signalling molecule, participating in mucosal sensory transduction.19 Our data support the fact that serotonin is able to signal to sensory neurons and is implicated in visceral pain, as we have shown here that intracolonic administration of serotonin induced visceral hypersensitivity. Further, we describe the role of TRPV4 in serotonin induce hyperalgesia and allodynia. The relationship between TRPV4 and serotonin has been described in rat pulmonary arterial smooth muscle cells, where serotonin is able to activate a TRPV4-like current by a the cytochrome P450 epoxigenase pathway.20 In vivo, serotonin provokes somatic hyperalgesia in response to mechanical stimuli. This hypersensitivity was not dependent on TRPV4 activation.21 Our results at the visceral level are in conflict with the somatic observations as the decrease of TRPV4 expression on sensory neurons inhibits serotonin-induced visceral hypersensitivity. The differences between the effects of TRPV4 in somatic and visceral nociception are also observed in basal conditions. The nociceptive response observed after colorectal distention is lower in TRPV4−/− than in wild-type mice,7 but the paw withdrawal frequency in response to application of a 0.17 mN von Frey hair is similar in TRPV4−/− and wild-type mice.21 Moreover, there are no significant differences in the baseline mechanical threshold for C-fibres in the saphenous nerve from wild-type and TRPV4−/− mice;22 however, for fibres projecting from the colon, the mechanosensory thresholds are increased in TRPV4−/−.23
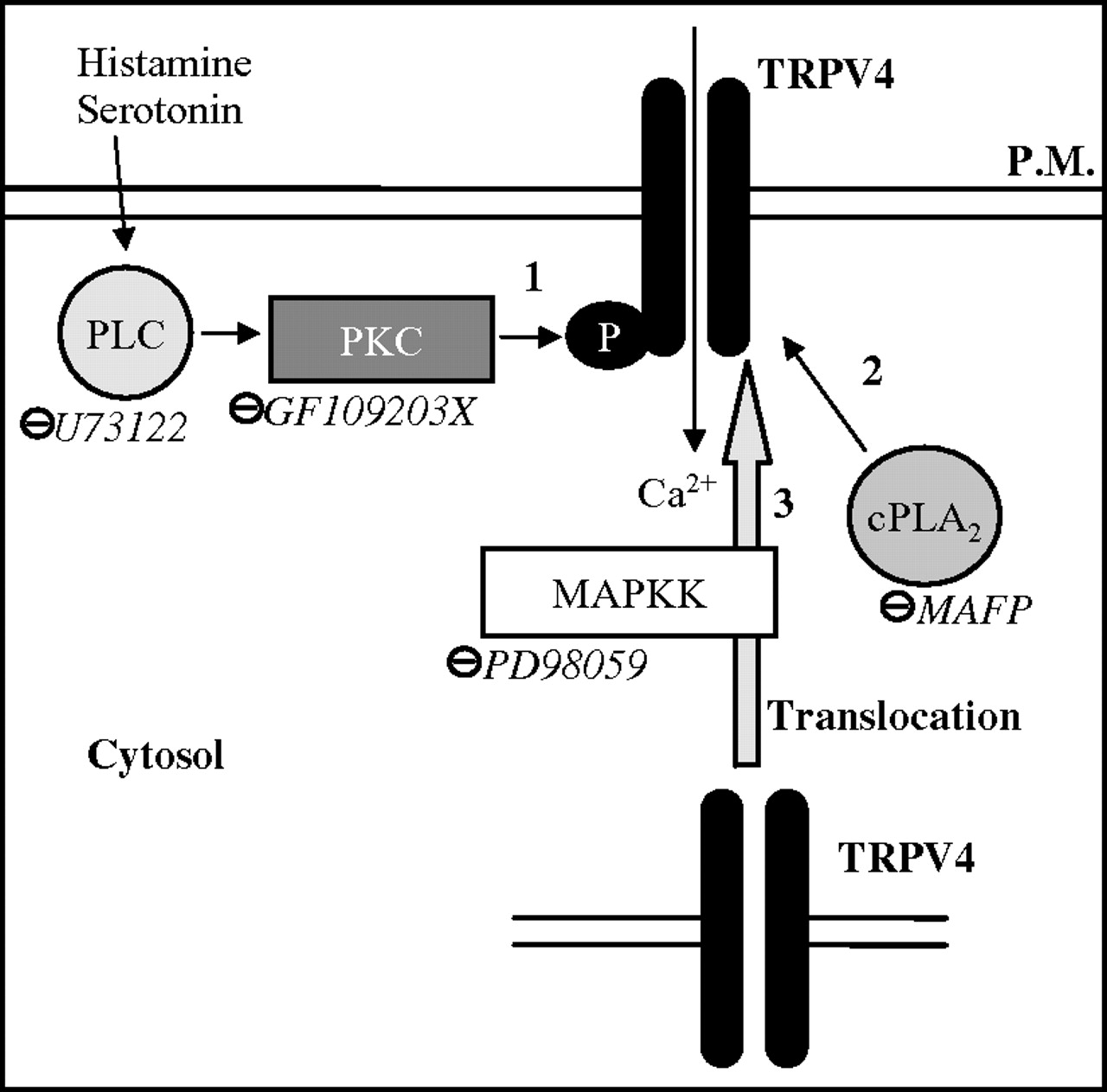
Pre-treatment of sensory neurons by serotonin or histamine enhanced Ca2+ signals in response to 4αPDD, suggesting that serotonin and histamine are able to sensitise TRPV4 activity. In a recent study, Jiang and collaborators demonstrated a crucial role of 5-HT3 and H1 receptor histamine in the stimulation of mesenteric afferents via 5-HT and histamine after intestinal anaphylaxis.24 Based on this study, here we demonstrate in sensory neurons that the sensitisation of TRPV4 by serotonin is mediated by the 5-HT3 receptor and the sensitisation by histamine is partially mediated by the H1 receptor. Further studies would be needed to evaluate the implication of other histamine receptors in TRPV4 sensitisation. Histamine or serotonin pre-treatment not only enhanced the amplitude of the response to TRPV4 agonist but also increased the number of responding neurons. This effect on amplitude and on the number of neurons recruited in response to TRPV4 agonist could reflect the consequences of TRPV4 phosphorylation and/or an increase of TRPV4 expression at the plasma membrane level. In non-stimulated neurons we observed that the repetition of TRPV4 expression was in favour of a perinuclear and nuclear expression, as already described in rat cortical astrocytes.25 Following histamine or serotonin treatments of sensory neurons, we observed an increased expression of TRPV4 at the plasma membrane level, concomitant with a decreased expression in the nuclear and perinuclear compartment. As this effect was observed only 5 min after the treatment, we can hypothesise that the increase of TRPV4 is not due to a new synthesis of TRPV4 but to an addressing from the nuclei to the plasma membrane. The translocation of TRPV4 to the membrane has already been described in endothelial cells.26 In our study, we observed that the MAPKK inhibitor was able to inhibit TRPV4 plasma membrane relocation induced by histamine or serotonin, but no other inhibitors (PKC, PLC, PKA or PLA2 inhibitors) modified this addressing of TRPV4. This indicates that a specific MAPKK pathway is involved in plasma membrane relocation of TRPV4 under serotonin or histamine exposure. Although, with the present results, we have clearly established that TRPV4 addressing to the plasma membrane is influenced by serotonin and histamine, concomitant phosphorylation of the receptor that would render it more susceptible to activation by its agonists cannot be ruled out. As a matter of fact, serotonin- and histamine-induced sensitisation of calcium response to TRPV4 agonists appears to be dependent on intracellular pathways different from the ones involved in TRPV4 plasma membrane addressing. Indeed, we observed that sensitisation of TRPV4 calcium response was dependent on a PKC and PLC mechanism. Following histamine or serotonin activation the activation of PKC by PLC has been well established.27 28 In our study, we can hypothesise that PKC, itself activated by PLC, provokes the phosphorylation of TRPV4 thereby increasing its responsiveness to agonists (figure 8). This is in agreement with the observation by De-Shou et al who demonstrated that phosphorylation of TRPV4 by PKC was able to sensitise this channel.29 PLA2 has also been described as a downstream event for TRPV4 activation: Vriens et al have shown that cell swelling-induced activation of TRPV4 requires PLA2 activity.30 In our study it seems that histamine and serotonin induced PLA2 activation, which then participates to the potentiation of TRPV4-induced calcium response. Based on our results and on the literature we can hypothesise that serotonin and histamine potentiate TRPV4 in three different ways: first, by the phosphorylation of TRPV4 depending on PLC and PKC activation; second, by an increase of its translocation mediated by a MAPKK pathway; and, finally, by the activation of PLA2, which increases the concentration of arachidonic acid metabolites. Among these metabolites we can hypothesise that PLA2 activation provokes an increase of 5,6-EET, which has been described as a TRPV4 agonist31 (figure 8).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mechanisms of histamine and serotonin potentiation of TRPV4 activation. We hypothesised that histamine and serotonin can potentiate TRPV4 by involving three different pathways. 1. Phosphorylation of TRPV4 by phospholipase C (PLC) and protein kinase C (PKC)-dependent pathways increases TRPV4 agonist-induced calcium signalling. 2. Translocation of TRPV4 from the nuclear and perinuclear zones to the plasmic membrane (PM) through a mitogen-activated protein kinase kinase (MAPKK)-dependent mechanism. 3. TRPV4-potentiation by a phospholipase A2 (PLA2)-related mechanism. ⊝, Inhibitors. TRPV4, transient receptor potential vanilloid 4.
Taken together, these results are in favour of a direct sensitising effect of serotonin and histamine receptors on TRPV4 cellular pools and/or TRPV4 agonist content. However, sensitisation of the nerve fibre as a whole, by exposure to doses of serotonin or histamine cannot be ruled out and might participate, at least in part, to the sensitisation of TRPV4 response.
Because of the important role shown for PAR2,9 histamine and serotonin10 in IBS-related pain symptoms, the fact that TRPV4 mediates histamine, serotonin and PAR2-induced hypersensitivity7 highlights TRPV4 as a possible common mediator on a clinical feature reported in all patients with IBS: visceral hypersensitivity. Importantly, our results suggest that TRPV4 directly or its regulation could be a new potential therapeutic target in patients with IBS for the treatment of abdominal pain.
Significance of this study
What is already known about this subject?
Histamine and serotonin play a role in visceral hypersensitivity
Transient receptor potential vanilloid 4 (TRPV4) caused visceral hypersensitivity
Hypotheses: TRPV4 is mediating histamine- and serotonin-induced visceral hypersentivity
What are the new findings?
Histamine or serotonin increased Ca2+ responses induced by 4αPDD by a protein kinase C (PKC), phospholipase Cβ (PLCβ), mitogen-activated protein kinase kinase (MAPKK) and phospholipase A2 (PLA2)-dependent mechanism
Serotonin or histamine treatments enhanced TRPV4 expression at the plasma membrane by a MAPKK mechanism
Histamine- or serotonin-mediated visceral hypersensitivity depend on TRPV4 expression in sensory neurons
How might it impact on clinical practice in the foreseeable future?
TRPV4 appears as a common mechanism to several known mediators of visceral hypersensitivity. Thus, TRPV4 directly or its regulation could be a new potential therapeutic target in patients with inflammatory bowel syndrome (IBS) for the treatment of abdominal pain.
Acknowledgments
The authors thank Dr Nigel W Bunnett from the University of San Francisco California, for providing the HEK-FLPTREX-TRPV4 cells.
References
Footnotes
NC and CA contributed equally to this work.
Funding This research was supported by grants from the Canadian Institute of Health Research (to GWZ and NV), the Crohn's and Colitis Foundation (to NV), the Fondation Bettencourt-Schueller (to NV), the INSERM-Avenir program (to NV), and the Fondation Schlumberger (to NV).
Competing interests None.
Ethics approval All procedures were approved by institutional Animal Care Committees and received an accreditation number (veterinary services).
Provenance and peer review Not commissioned; externally peer reviewed.