Article Text

Abstract
Objective CD103+ gut dendritic cells (DCs) have been shown to be required for de novo conversion of adaptive T regulatory (Treg) cells. Indoleamine 2,3-dioxygenase (IDO) is an enzyme involved in tryptophan catabolism that is expressed by DCs isolated from tumour-draining lymph nodes. IDO-expressing DCs sustain and differentiate Tregs. The aim of this study was to investigate the expression and the possible physiological role of IDO in the tolerogenic properties of intestinal DCs.
Design The expression level of IDO in CD103+ and CD103− DCs was analysed by qRT-PCR, western blot and immunofluorescence. CD103+ and CD103− DCs were sorted from mesenteric lymph nodes (MLNs) and the small intestinal lamina propria, and the role of IDO in the conversion of Tregs and Th effector cell development was evaluated via specific inhibition or gene deletion. Oral tolerance, experimental colitis and T cell differentiation in vivo were assessed upon IDO inactivation.
Results We show that, primarily, CD103+ but not CD103− gut DCs express IDO whose inhibition results in reduced CD4+Foxp3+ T regulatory cell conversion and enhanced T cell proliferation. When IDO was inhibited or genetically deleted there was an increase in Th1 and Th17 differentiation both in vitro and in vivo. Finally, in vivo IDO blockade affected the development of Tregs specific for orally administered antigens, impaired oral tolerance induction and exacerbated colitis.
Conclusions We identified a new IDO-dependent pathway leading to acquisition of tolerogenic functions in mucosal CD103-expressing DCs, indicating IDO as a possible therapeutic target for gut disorders.
- CD103+ lamina propria dendritic cells
- indoleamine 2,3-dioxygenase
- T regulatory cells
- dendritic cells
- lamina proprial lymphocytes
- oral tolerance
- T lymphocytes
- DC
- dendritic cell
- Treg
- regulatory T cell
- IDO
- indoleamine 2,3-dioxygenase
- OVA
- ovalbumim
- 1-MT
- 1-methyl tryptophan
- DSS
- dextran sodium sulfate
Statistics from Altmetric.com
- CD103+ lamina propria dendritic cells
- indoleamine 2,3-dioxygenase
- T regulatory cells
- dendritic cells
- lamina proprial lymphocytes
- oral tolerance
- T lymphocytes
- DC
- dendritic cell
- Treg
- regulatory T cell
- IDO
- indoleamine 2,3-dioxygenase
- OVA
- ovalbumim
- 1-MT
- 1-methyl tryptophan
- DSS
- dextran sodium sulfate
Introduction
Dendritic cells (DCs) play a fundamental role in the homeostasis of the gut by inducing tolerance to food and self-antigens.1–4 These specialised functions are carried out by a distinct subset of mucosal DCs expressing the type I transmembrane glycoprotein known as the αE integrin (CD103) surface marker.1 2 CD103+ DCs isolated both from the lamina propria (LP) and mesenteric lymph nodes (MLNs) have been shown to drive preferentially the development of CD4+Foxp3+ T regulatory cells via a transforming growth factor β (TGFβ) and retinoic acid dependent mechanism.5–7 These cells migrate directly from the LP where they have presumably acquired their tolerogenic non-inflammatory phenotype.8 Indeed, local environmental factors released by intestinal epithelial cells, including thymic stromal lymphopoietin,9 10 TGFβ and retinoic acid,7 11 can control DC function by inhibiting the release of interleukin 12 (IL12) and the ability to drive T helper (Th)1 T cells, and by skewing T regulatory cell development.
Systemic oral tolerance is a process that generates unresponsiveness versus food antigens when administered systemically.12 DCs have been shown to be involved in the induction of oral tolerance. Indeed, DC expansion at mucosal sites correlates with enhanced tolerance to food antigens.13 Tolerance is initiated at MLNs and antigen transport by DCs has been shown to be essential for this process.14 However, the exact nature and the mechanisms employed by DCs have not been clarified.
Indoleamine 2,3-dioxygenase (IDO) is an enzyme involved in tryptophan catabolism. Its immunosuppressive effects are linked to the reduction of local tryptophan concentration and to the production of immunomodulatory tryptophan metabolites.15 These have detrimental effects on T cell proliferation both directly (via tryptophan deprivation) and indirectly via the activation or de novo induction of T regulatory cells.16 17 IDO has been primarily associated with the tolerogenic properties of plasmacytoid DCs (pDCs) isolated from tumour-draining lymph nodes.18 These cells are responsible for the activation of mature Tregs16 but may also be responsible for de novo Treg conversion.17 19 IDO has been postulated as a possible mediator of the tolerogenic function of DCs in the gut.20
In this study we found that IDO is highly expressed in the intestine of mouse and is associated with CD103+ DCs. Selective inhibition or genetic deletion of IDO affects the development of antigen-specific CD4+Foxp3+ Tregs and promotes Th1/Th17 cell differentiation.
Materials and methods
Mice
C57BL/6 Ly5.1+ mice, BALB/c mice and OT-II TCR Ly5.2+ transgenic mice were purchased from Charles River Laboratories (Charles River, Milan, Italy). The Ido1−/− mice were generated in the laboratory of A.L. Mellor (at Medical College of Georgia, Augusta, Georgia, USA) and back-crossed for ten generations to BALB/c mice.21 All the mice were bred and maintained in a specific pathogen-free animal facility at IFOM-IEO Campus, Milan, Italy.
Isolation and culture of CD11c subsets
CD103+ and CD103– DCs were isolated from murine MLNs with cell sorting up to >95% purity. The details of the procedures for isolation and the sorting strategy used are described in supplementary materials and methods.
In vitro T cell conversion assay
T cell conversion was performed as previously described,5 with minor modifications as detailed in supplementary materials and methods.
Adoptive transfer and antigen oral feeding
Naïve T cells CD4+CD25−CD62L+ (2×106) isolated from OT-II Ly5.2+ mice were adoptively transferred intravenously into C57BL/6 Ly5.1+ recipients implanted on the dorsal skin with slow-release pellets containing 200 mg 1-methyl-dl-tryptophan or placebo pellets 1 day before (7-day release at <1 mg/h; Innovative Research of America, Sarasota, Florida, USA). After 12 h animals received 1.5% ovalbumin in the drinking water for 5 days (grade III; Sigma-Aldrich, St Louis, Missouri, USA). On day 6, spleen, MLNs, PPs and small intestine were collected from Ly5.1+ hosts, and Foxp3 expression was analysed in the Ly5.2+ transplanted cells by flow cytometry.
Tolerance induction by intragastric antigen delivery and assessment of delayed type hypersensitivity to OVA
Oral tolerance to ovalbumin and assessment of delayed-type hypersensitivity (DTH) to OVA was performed as previously described,22 with minor modifications as detailed in supplementary materials and methods.
T cell transfer model of colitis
Induction of colitis with naïve CD4+CD25−CD45RBhi T cell transfer to Rag1−/− mice was performed as previously described.23 Rag1−/− recipient mice were implanted with slow-release pellets of 1-methyl-dl-tryptophan or placebo pellets 1 day before T cell transfer until the end of the experiments (release at <1 mg/h; Innovative Research of America). Details regarding CD4+CD25−CD45RBhi T cell purification and assessment of intestinal inflammation are described in supplementary materials and methods.
Induction of DSS colitis
Detailed procedures for induction of dextran sodium sulfate (DSS) colitis are described in supplementary materials and methods.
Isolation and analysis of leucocyte subpopulations
Cell suspensions were prepared from spleen, MLN, PP and the LP as previously described.6 Foxp3 and intracellular IL17/interferon (IFN)γ staining was performed according to the manufacturer's instructions and is described in detail in supplementary materials and methods.
Isolation of human colonic lamina propria cells
CD103+ and CD103− DCs were isolated from healthy colonic specimens derived from patients undergoing surgery for colon cancer. Details of the procedures for isolation of LP cells and the sorting strategy are described in supplementary materials and methods.
Immunofluorescence staining and immunoblotting
Details concerning the immunofluorescence staining and immunoblotting of IDO are described in supplementary materials and methods.
Quantitative RT-PCR analysis
Details of the procedures and primers used for the quantitative RT-PCR are described in supplementary materials and methods.
Statistical analysis
The Student paired t test was used to determine statistical significance, which was defined as *p<0.05, **p<0.01, and ***p<0.001. Statistical calculations were performed using JMP 5.1 software.
Results
IDO is highly expressed throughout the intestine
Expression of IDO was analysed in the small (duodenum, jejunum and ileum) and large (colon) intestines and in the spleen of wild-type mice. RNA and proteins were extracted from homogenised tissue and IDO expression was quantified by qRT-PCR and western blot. IDO was found to be primarily expressed in the small intestinal tracts with a marked presence in the jejunum and ileum, both at the RNA and protein level (figure 1A,B).

IDO is expressed throughout gut tissues. Tissue samples were collected from C57BL/6 mice to extract RNA and proteins. (A) Indo expression was analysed by qRT-PCR and normalised to the control gene Rpl32. Data represent mean±SD from five mice. (B) The expression of IDO protein was analysed by western blot in total tissue extracts. Histograms represent relative quantification of IDO protein versus GAPDH. One of two independent experiments is shown. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; IDO, indoleamine 2,3-dioxygenase.
Lamina propria CD103+ DCs express IDO both in mouse and human
We then sought to identify the IDO+ cells present in the murine gut. Confocal microscopy images revealed that IDO was associated largely with CD11c+ DCs (figure 2A). Indeed, a consistent proportion of the IDO+CD11c+ cells were found in the upper part of the villi.

CD11c+CD103+ dendritic cells in the LP express IDO. (A) Immunofluorescence images of frozen tissue sections from small bowel of C57BL/6 mice stained with α-CD11c (panel a, red) and α-IDO (panel b, green). Panel d shows the overlay of IDO (green) with CD11c (red) and DAPI (blue). Original magnification, ×400. Panel e represents a 5-fold magnification of the white inset in panel d. (B) Intracellular expression of IDO by LP CD103− (green histogram) and CD103+ (red histogram) DCs was assessed by flow cytometry. The blue histogram represents the isotype control for the IDO primary antibody. (C) Mean fluorescence intensity (MFI) of intracellular IDO expression in CD11c+CD103− (grey bars) or CD11c+CD103+ cells (black bars)±SD of six mice analysed independently. Data are representative of one out of three (A–C) independent experiments. ***p<0.001. DAPI, 4′-diamidine-2′-phenylindol dihydrochloride; DCs, dendritic cells; IDO, indoleamine 2,3-dioxygenase; LP, lamina propria.
To assess which subtype of DCs expressed IDO, LP mononuclear cells were isolated from the murine small bowel and analysed by flow cytometry. As shown in figure 2B,C, IDO expression levels were significantly higher in CD103+ DCs compared to the CD103− counterpart.
LP CD103+ DCs express CCR7 and migrate to the MLN,24–26 thus we sorted MLN CD11c+ cells into CD103+ and CD103− subsets (figure 3A) and tested them for IDO expression. Consistent with the data obtained in the LP, we found that IDO was primarily expressed in CD11c+CD103+ DCs, both at the RNA and protein level (figure 3B,C). These cells resembled migratory conventional DCs as they predominantly expressed CD11b and CCR7, and only a minority expressed CD8, B220 or Ly6C (figure 3D).

CD103+ DCs from MLNs express IDO. (A) CD11c+ DCs were enriched from MLNs and sorted by FACS into CD103+ and CD103− subsets. CD11c+CD103− and CD11c+CD103+ cells were collected and RNA or proteins were extracted. (B) Expression level of Indo relative to Rpl32 in CD11c+CD103− (grey bar) and CD11c+CD103+ (black bar) was analysed by RT-PCR. (C) Immunoblot for IDO in sorted CD11c+CD103− and CD11c+CD103+ cells is shown. Histograms represent relative quantification of IDO protein versus vinculin (Vinc). (D) Phenotypic characterisation of CD103− (green histogram) and CD103+ (red histogram) CD11c+ DCs isolated from MLNs is depicted. Blue histograms represent relative isotype control. Cells were incubated with mAbs against the indicated markers and analysed by flow cytometry. Data are representative of one of three (B–D) experiments. (E) Human colonic LP DCs were sorted as lineage (Lin)-negative HLA-DR+ CD11c+ cells and subsequently separated according to CD103 expression. Indo expression in CD11c+CD103− and CD11c+CD103+ subsets was evaluated by qPCR and normalised to β-actin. The fold increase in the Indo/β–actin ratio of the CD11c+CD103+ subset with respect to CD11c+CD103− is depicted. Colonic normal tissues from five different patients were evaluated. **p<0.01; ***p<0.001. DCs, dendritic cells; FACS, fluorescence activated cell sorting; IDO, indoleamine 2,3-dioxygenase; LP, lamina propria; mAb, monoclonal antibody; MLN, mesenteric lymph node.
IDO is primarily involved in the conversion of the amino acid tryptophan to kynurenine, an intermediate in the synthesis of niacin. To prove that CD11c+CD103+ DCs expressed a functionally active form of IDO, the amount of kynurenine converted from tryptophan was quantified. In agreement with IDO expression, CD103+ DCs released a significantly higher amount of kynurenine compared to the CD103− DCs (supplementary figure 1).
CD11c+CD103+ DCs have also recently been described in human MLNs, and have been shown to drive Treg cell development and imprint gut homing properties to T cells.7 24 Thus, IDO expression in human lineage (Lin)-negative HLA-DR+CD11c+CD103+ DCs sorted from human colonic LP was assessed (figure 3E). Similarly to mouse LP DCs, human intestinal CD103+ DCs expressed higher levels of Indo mRNA when compared to CD11c+CD103− DCs.
Together, these results indicate that both human and mouse intestinal CD103+ DCs express IDO.
Inactivation of IDO influences the tolerogenic properties of CD103+ DCs
MLN CD103+ DCs have been shown to enhance Foxp3+ Treg differentiation in a TGFβ and retinoic acid-dependent fashion.5–7 To address whether IDO expression was involved in the tolerogenic properties of DCs, CD11c+CD103+ and CD11c+CD103− DCs were sorted from murine MLNs and the activity of IDO was selectively inhibited during T cell activation. Sorted cells, incubated with or without the IDO inhibitor 1-methyl-dl-tryptophan (1-dl-MT), were loaded with ovalbumin (OVA) and cultured with highly purified naïve CD4+CD25−CD62L+Foxp3− OVA-specific OT-II T cells (supplementary figure 2). As shown in figure 4A,B, IDO inhibition significantly affected the ability of CD103+ DCs to promote conversion of naïve T cells into Foxp3+Tregs while the ability of CD103− cells was unaffected. In addition, an enhancement of T cell proliferation after 1-MT treatment was detected in the CD103+ DCs-T cell cultures (supplementary figure 3).
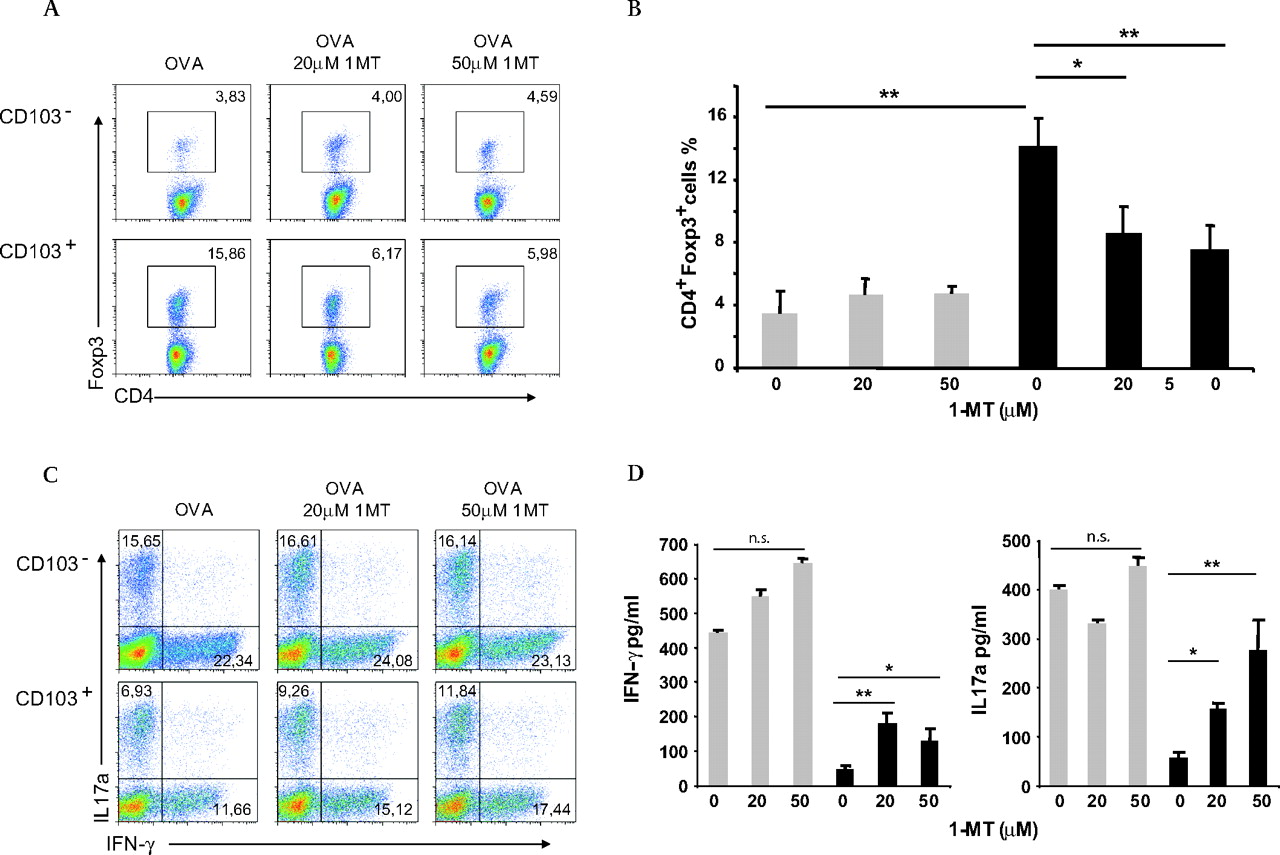
CD103+ DCs from MLNs display unique IDO-dependent tolerogenic properties. (A) CD11c+CD103+ and CD11c+CD103− cells were isolated from MLNs of C57BL/6 mice by cell sorting. Cells treated with different concentrations of 1-MT, as indicated, were loaded with OVA and co-cultured with CD4+CD25−CD62L+ T cells from OT-II mice. Recombinant TGFβ1 was added after 3 days of culture. On day 6, conversion of naïve T cells into CD4+Foxp3+ cells was determined by flow cytometry. Numbers represent the percentage of Foxp3+ cells in the CD4+ subset. (B) Percentages of CD4+Foxp3+ cells in different conditions are shown as means of samples from three combined experiments±SD. (C) CD4 naïve OT-II T cells were co-cultured with OVA-loaded CD11c+CD103− (grey bars) or CD11c+CD103+ cells (black bars) as in A. On day 5 cells were re-stimulated for 5 h at 37°C in the presence of PMA, ionomycin and Golgistop and stained for CD4, IFNγ and IL17. Dot plots represent the percentage of CD4+IFNγ+ and CD4+IL17+ cells. (D) On day 5, cytokine content in the supernatants was measured by cytometric bead array (IFNγ, IL17a). Graphic bars represent combined means from three independent experiments±SD. *p<0.05; **p<0.01. DCs, dendritic cells; IDO, indoleamine 2,3-dioxygenase; IFN, interferon; IL, interleukin; MLN, mesenteric lymph node; 1-MT, 1-methyl tryptophan; OVA, ovalbumin; PMA, phorbol 12-myristate 13-acetate; TGF, transforming growth factor.
Since 1-methyl-dl-tryptophan is a mixture of two isomers and 1-d-MT has been shown to be more effective in inhibiting IDO activity, the two isomers (1-d-MT and 1-l-MT) were independently tested.27 In line with previous reports, only 1-d-MT treatment was capable of reducing the ability of the IDO-expressing CD11c+CD103+ DCs to convert Foxp3+Treg cells (supplementary figure 4).
We then analysed T cell polarisation induced by MLN CD103+ DCs upon 1-MT treatment. Interestingly, IDO inhibition almost doubled the polarisation of naïve CD4+ T cells into IL17-producing cells (figure 4C,D). At the same time inhibition of IDO influenced IFNγ production; however, it did not completely restore its secretion from T cells (figure 4C), presumably because CD103+ DCs had received tissue conditioning in the LP. These observations are in agreement with recent reports in which IDO blocking in pDCs fostered Th17 effector cell development.28 29 At the same time IDO inhibition induced IL10 production in activated T cells (supplementary figure 5).
Upon re-stimulation of T cells with α-CD3 and α-CD28 antibodies we did not observe a skewing towards a Th2 phenotype as demonstrated by the lack of IL13 and IL5 in cell cultures. It should be noted that while there was an increase in IL4 production, this difference only became statistically significant upon administration of 20 μmol/l 1-MT (supplementary figure 5).
IDO blockade alters Treg cell development in vivo
We then tested whether IDO was involved in Treg cell differentiation in vivo. IDO blockade was achieved by subcutaneous implantation of pellets releasing 1-MT for seven consecutive days. One day after pellet implant Ly5.1 mice were adoptively transferred with highly purified naïve CD4+CD25−CD62L+ Ly5.2+ OT-II T cells and were given drinking water containing 1.5% OVA for five consecutive days. On day 6, mice were killed and cells from the spleen, MLN, PP and LP were collected. The frequency of CD4+Ly5.2+ T cells positive for Foxp3 was assessed by flow cytometry. As shown in figure 5A,B, CD4+ Ly5.2+ T cells proliferated primarily in the LP, PP and MLN where they gave rise to a higher frequency of Foxp3+ Tregs as compared to the spleen. By contrast, in mice treated with 1-MT the frequency of Tregs in LP, PP and MLN was significantly reduced while no difference was observed in the spleen (figure 5A,B).

IDO inhibition affects the conversion of naïve CD4+ Ly5.2+ OT II T cells into Foxp3+ cells in GALT after oral administration of OVA. (A) Ly5.1 recipient mice were implanted with slow-release pellets containing either 1-methyl-dl-tryptophan (1-MT, 200 mg, 7-day release at <1 mg/h) or placebo. After 1 day mice had been adoptively transferred with CD4+CD25−CD62L+ naïve Ly5.2+ OT-II T cells. Recipient mice were then fed with OVA antigen in the drinking water for 5 days. On day 6, conversion of CD4+Ly5.2+ into Foxp3+ expressing T cells was assessed in various tissues by flow cytometry. By gating on Ly5.2+CD4+ cells, the percentage of intracellular Foxp3 expressing cells was calculated. Percentage of Ly5.2+Foxp3+ cells is depicted for spleen (SP), MLN and PPs. (B) Summary of the percentage of Ly5.2+ OT-II T cells expressing Foxp3 in the placebo (grey bars) and in the 1-MT (black bars) treated group. Data were combined from two independent experiments. (C) Mice implanted (black bars) or not (grey bars) with 1-MT releasing pellets were fed twice either with PBS or 20 mg OVA as depicted in the flow chart. Seven days after the last antigen gavage, mice were immunised by subcutaneous injection of OVA in PBS/CFA emulsion and challenged 7 days later in the footpad with 50 μg of OVA. (D) Induction of tolerance was analysed by measurement of the DTH reaction 48 h after challenge (mean footpad swelling±SD of five mice per group). Data are representative of one out of two independent experiments. (E) OVA-specific IFNγ production by spleen cells collected 48 h after challenge was measured by CBA Flex set upon re-stimulation in vitro with 0, 100 or 1000 μg/ml of OVA (mean/ml±SD of triplicate wells). *p<0.05; **p<0.01; n.s. not statistically significant. CBA, cytometric bead array; CFA, complete Freund's adjuvant; DTH, delayed-type hypersensitivity; GALT, gut-associated lymphoid tissue; IDO, indoleamine 2,3-dioxygenase; IFN, interferon; MLN, mesenteric lymph node; OVA, ovalbumin; PBS, phosphate-buffered saline; PPs, Peyer's patches.
Thus, after oral exposure to antigen, IDO plays a primary role in the induction of Tregs in the gut associated lymphoid tissue (GALT) in vivo.
IDO inhibition impinges on the development of oral tolerance
The involvement of IDO in the generation of Tregs in vivo prompted us to study oral tolerance establishment in a setting of IDO inhibition during antigen feeding.
As reported above, mice were implanted subcutaneously (s.c.) with pellets releasing placebo or 1-MT over the course of 7 days. Two days after implantation OVA was intragastrically administered to induce oral tolerance. Nine days later mice were challenged s.c. with OVA in cytometric bead array (CFA) to induce systemic immunity and a week later a final dose of adjuvant-free OVA was injected into the footpad to evaluate the delayed-type hypersensitivity reaction (DTH) as revealed by footpad swelling (figure 5C). As expected, mice that received oral OVA and placebo pellets showed reduced DTH, while mice implanted with 1-MT pellets during OVA administration showed no established systemic tolerance (figure 5D). We then evaluated whether spleen cells isolated from OVA-fed mice had an impaired capacity to produce IFNγ. Consistent with the induction of oral tolerance, spleen cells isolated from OVA-fed mice receiving placebo pellets showed reduced ability to release IFNγ when re-stimulated with different amounts of OVA in vitro (figure 5E). By contrast, in mice receiving both OVA and 1-MT, IDO inhibition resulted in restored IFNγ production in culture supernatants (figure 5E). Together these data suggest that IDO inhibition in vivo results in the reduction of antigen-specific Treg expansion and the concomitant reduction of systemic oral tolerance.
Altered balance of Treg and T effector cells in the gut of IDO deficient mice
To test whether indoleamine 2,3-dioxygenase in the gut could regulate the balance between Treg and T effector cells, IDO deficient mice were studied. CD4+ T cells isolated from the small bowel of Ido1+/+ and Ido1−/− mice were stained for Foxp3 or re-stimulated in vitro to assess IL17 and IFNγ production (figure 6A,B).

Lack of IDO activity in the gut alters the IFNγ–IL17 balance and affects Treg development. (A) LP mononuclear cells were isolated from the small intestine of Ido1+/+ and Ido1−/− mice. Isolated cells were re-stimulated in vitro with PMA, ionomycin and GolgiStop for 5 h. Intracellular staining to assess IFNγ and IL17 content was performed. Dot plots show percentages of CD4+ IFNγ+ and CD4+ IL17+ cells. Five mice per group were analysed independently, as shown in the graphs. (B) Isolated cells from Ido1+/+ and Ido1−/− mice were stained for CD4 and Foxp3. Percentages of double-positive cells are shown in the dot plots; data from five mice per group are gathered in the graphs. Each symbol represents an individual mouse. (C) C57BL/6 mice were implanted with slow-releasing pellets containing 1-methyl-dl-tryptophan (1-MT, 200 mg, 7-day release at <1 mg/h) or placebo. On day 7, LP mononuclear cells were isolated from the small intestine and re-stimulated in vitro as described in (A). The percentage of CD4+ IFNγ+ and CD4+ IL17+ cells is shown in the dot plots and in the graphs, as five mice per group were analysed independently. Each symbol represents an individual mouse. (D) LP mononuclear cells isolated from placebo or 1-MT-treated mice were stained for CD4 and Foxp3. The percentage of CD4+ Foxp3+ cells is shown in the dot plots; for each group, percentages of CD4+ Foxp3+ cells from five mice are summarised in the graph. Each symbol represents an individual mouse. Data are representative of one out of two (A–D) independent experiments. *p<0.05; **p<0.01; n.s. not statistically significant. IDO, indoleamine 2,3-dioxygenase; IFN, interferon; IL, interleukin; LP, lamina propria; PMA, phorbol 12-myristate 13-acetate; Tregs, regulatory T cells.
In accordance with our in vitro evidence, IDO deficient mice exhibited a marked alteration in the pattern of T cell polarisation in the gut. Indeed, Ido1−/− mice had an almost double proportion of IL17+CD4+ T cells in the intestine compared to wild-type animals, as well as a notable increase in the number of IFNγ+ secreting CD4+T cells (figure 6A). Remarkably, IDO deficient mice also displayed a decreased percentage of Foxp3+ Tregs in the LP (figure 6B), while no such difference was observed in the spleen (data not shown). These observations serve to support our hypothesis that IDO is a critical regulator of immune homeostasis in the gut.
Similar results were obtained in mice that were exposed to 1-MT, which displayed a pronounced increase in the proportion of Th1 and Th17 cells in the gut, although no change in the Tregs frequencies was observed (figure 6C,D). This is probably due to pre-existing Fop3+ cells before IDO inhibition.
IDO gives protection from T cell-mediated and DSS colitis
At the steady state, IDO proved to be involved in the development of antigen specific Tregs in the gut and oral tolerance in vivo.
We next sought to investigate whether the inhibition of IDO could predispose mice to intestinal inflammation or exacerbate it. To achieve this, we utilised a well-characterised colitis model by adoptively transfering naïve T cells into Rag1−/− mice. The animals, either treated with 1-MT or untreated controls, were injected i.p. with naïve CD4+CD25−CD45RBhiCD45.1+ T cells isolated from spleens of C57BL/6 mice.
The progression of the disease was monitored by body weight loss, diarrhoea and the appearance of blood in the faeces. Fifty days after cell transfer, mice showed evident signs of colitis (eg, diarrhoea, blood in faeces, marked body weight loss). Mice were killed and their colons were dissected in order to evaluate histology score, T cell infiltration and cytokine content.
1-MT treatment significantly worsened the symptoms of colitis. As shown in figure 7A IDO inhibition induced a greater loss in body weight as compared to the placebo group (figure 7A). Accordingly, when IDO was blocked, mice showed extensive inflammation in the colon with a clear increase in the levels of inflammatory infiltrates, epithelial hyperplasia and goblet cell depletion in the colon (figure 7B,C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
IDO inhibition worsens symptoms in a T cell-transfer colitis model. Rag1−/− mice were implanted on the dorsal skin with 1-methyl-dl-tryptophan (1-MT, 200 mg, 7-day release at <1 mg/h) or placebo slow-releasing pellets 1 day before transplant of 5×105 naïve CD4+CD45RBhi cells intraperitoneally. Pellets were substituted weekly until the end of the experiment. (A) Variation in body weight was followed for the entire period and expressed as percentage difference between the initial body weight and the actual body weight measured every 5 days. (B) Proximal colon of Rag1−/− mice 50 days after cell transfer; sections are stained with haematoxylin and eosin. (C) Histology scores of sections of the large intestine after 50 days from CD4+CD45RBhigh cell transfer. (D) Absolute number of CD4+ T cells from colonic LP in Rag1−/− mice 50 days after CD4+CD45RBhigh cells transfer. (E,F) Absolute numbers of IFNγ and IL17a producing CD4+ T cells as determined by flow cytometry. (G) RNA was extracted from total colonic tissues and qPCR was performed. Expression of Ifng, Il17a and Il17f was evaluated and normalised to Rpl32. IDO, indoleamine 2,3-dioxygenase; IFN, interferon; IL, interleukin; LP, lamina propria.
In order to address whether IDO could affect T effector cell activation during colitis, the amounts of Th1 and Th17 cells in the LP were evaluated by intracellular cytokine staining after in vitro stimulation with phorbol 12-myristate 13-acetate (PMA)/ionomycin of colonic LP cells. First, we observed a distinct increase in the number of CD4+CD45.1+ T cells infiltrating the LP of 1-MT-treated Rag1−/− hosts in comparison to placebo controls (figure 7D). Moreover, 1-MT enhanced cytokine production in T cells with a noticeable increase of CD4+ T cells producing IFNγ and IL17 (figure 7E,F).
In contrast, at the mRNA level, pro-inflammatory cytokines, such as IFNγ, IL17a and IL17f, were significantly upregulated in colon homogenates from 1-MT-treated mice compared to placebo-treated mice (figure 7G).
Similarly, in chemical (DSS)-induced colitis IDO inhibition aggravated symptoms of intestinal inflammation. The 1-MT-treated group displayed a more pronounced loss in body weight, mortality rate and colon shortening together with an increased disease activity index (DAI) due to severe diarrhoea and the appearance of blood in the faeces (supplementary figure 6A–F). Likewise, lack of IDO activity exacerbated intestinal inflammation and tissue damage (supplementary figure 6B). The expression of IDO was upregulated after 1-MT treatment in CD103+ but not CD103− DCs (supplementary figure 7). This was expected as IDO is induced by IFNγ, whose expression is enhanced during IDO inhibition (supplementary figure 6F).
Together, these data suggest that IDO may play an essential role in regulating inflammation through the control of Th1 and Th17 balance in the gut.
Discussion
The immunosuppressive effects of IDO are linked to the reduction of local tryptophan concentration and to the production of immunomodulatory tryptophan metabolites15 that can participate in Treg differentiation and activation.16 17 IDO has been shown to be expressed by DCs isolated from tumour-draining lymph nodes18 but little was known concerning the function of IDO under physiological conditions.
In this study we analysed the expression of IDO by LP DCs. We found that IDO was associated with CD11c+CD103+ DCs, which have been shown to be required for de novo Treg conversion.5 6 CD103+ DCs have been shown to release TGFβ and to metabolise or accumulate retinoic acid. These compounds are involved both in the development of Tregs5 6 and in conferring gut homing properties to T and B cells.24 30 31 Inhibition of IDO using 1-MT reduced the ability of CD103+ DCs to drive Foxp3+ Treg cell development in vitro. Although the IDO-mediated pathway for Treg cell conversion may not be the only one, our results clearly show that it plays an important role. Interestingly, blockade of IDO activity was associated with increased T cell proliferation and release of a peculiar pattern of cytokines (IL17, IFNγ, IL10 and partly IL4). We have shown that epithelial cell-derived mediators can ‘educate’ DCs with a full mucosal phenotype. They can induce the expression of CD103 and the ability to drive Tregs;11 and at the same time they can inhibit IL12 release and the differentiation of IFNγ-producing T cells.9 10 On the other hand, DCs may have also been exposed to commensal bacteria-derived components in the LP. Microbiota-derived metabolites, including ATP, have recently been shown to drive the development of Th17 polarising DCs.32 33 It is likely that CD103+ DCs were exposed to microbial products in the LP and hence acquired the ability to drive Th17 and Th1 T cells, a capacity that was inhibited by the activity of IDO. Hence IDO expression by DCs is required to both induce Foxp3+ Treg differentiation and to inhibit IL17 production, thus contributing to intestinal homeostasis. The skewing towards a Th17 phenotype is very interesting as it has been described that, in mouse chronic granulomatous disease, tryptophan catabolism is impaired – presumably due to defects in IDO expression – and this is associated with an increase in Th17 inflammatory cells.34 In agreement, IDO inhibition in pDCs isolated from tumour draining lymph nodes or spleen pDCs activated with CpG DNA skews towards a Th17 phenotype.28 29
Systemic oral tolerance is a process that allows generating unresponsiveness versus food antigens when administered systemically.12 DCs have been shown to be involved in the induction of oral tolerance,13 but the exact nature and the mechanism employed by DCs has not been identified. We show here that administration of 1-MT affected the establishment of oral tolerance indicating that IDO expression by CD103+ DCs may be involved in this process. Recently, a role for liver pDCs in the induction of oral tolerance to haptens has been proposed.22 However, the expression of IDO or the generation of Tregs was not analysed.22 In our study, CD103+ DCs only marginally expressed markers of pDCs, like Ly6C or B220, and the majority belonged to the conventional type of DCs (CD11b+). It would be interesting to study whether IDO-expressing DCs are mainly involved in Treg development while liver pDCs are involved in clonal cell deletion. Hence IDO plays an important role both in acquired Treg development and tolerance to oral antigens. It also plays a protective role in colitis as administration of 1-MT augments trinitrobenzene sulfonic acid (TNBS),35 DSS and adoptive transfer (this study) colitis presumably skewing the balance between Tregs and Th1/Th17 cells in the LP. What induces the expression of IDO in CD103+ DCs remains to be established, but recent data have suggested that TGFβ may transform IDO− into IDO+ DCs.36 As TGFβ is abundantly expressed in the gut and is also released by mucosal DCs,11 it is possible that it might be involved in upregulating IDO expression. Alternatively, in human DCs, prostaglandin (PG) E2 has been shown to upregulate the expression of IDO.37 Since PGE2 is highly abundant in the gut38 it may induce IDO expression. These two possibilities are not mutually exclusive.
Altogether, our data point to a fundamental role of IDO in the tolerogenic properties of CD103+ DCs. IDO expressed by DCs can participate in several ways to the development of Tregs,16 including the inhibition of T cell proliferation and differentiation to Th17 cells. The functional interaction of IDO with DC-derived and epithelial cell-derived TGFβ and retinoic acid that are also required for Treg cell development remains to be elucidated.5 6 11 One possible explanation may be that inhibition of T cell proliferation by IDO may facilitate the retinoic acid-dependent TGFβ activity on Foxp3 upregulation;39–41 alternatively, and not mutually exclusive, TGFβ may be required to sustain IDO expression.36 IDO inhibition resulted in impairment in Treg cell differentiation that was associated with restoring IL17 producing cells. It is likely that LP-migrated DCs received local conditioning derived both by host mediators and by the microbiota. Together, these participate in conferring the specialised mucosal phenotype of resident DCs and the balance between these two signals may control tolerance versus immunity. In conclusion, our results indicate that IDO is a possible new therapeutic target in intestinal inflammatory disorders.
Significance of this study
What is already known about this subject?
CD103+ dendritic cells (DCs) isolated both from the lamina propria (LP) and the mesenteric lymph nodes (MLNs) have been shown to drive preferentially CD4+Foxp3+ T regulatory cells via a transforming growth factor β (TGFβ) and retinoic acid dependent mechanism
Indoleamine 2,3-dioxygenase (IDO) is an enzyme involved in tryptophan catabolism with immunosuppressive effects
IDO activation has detrimental effects on T cell proliferation both directly (via tryptophan deprivation) and indirectly via the activation or de novo induction of Treg cells
IDO has been primarily associated with the tolerogenic properties of plasmacytoid DCs (pDCs) isolated from tumour-draining lymph nodes
What are the new findings?
IDO is expressed in the gut primarily by CD11c+CD103+ DCs both in human and mouse systems
IDO is involved in the ability of CD103+ DCs to drive Foxp3+ Treg cell development
IDO activity is required for the establishment of oral tolerance
IDO controls the balance between Treg and T effector cells in the LP
IDO inhibition worsens T cell-mediated and dextran sodium sulfate (DSS) colitis
How might it impact on clinical practice in the foreseeable future?
In this manuscript we show that IDO expression in intestinal DCs is important for regulating intestinal immune homeostasis by keeping the balance between Foxp3+ Tregs and Th1/Th17 effector cells in the lamina propria. Deregulation of IDO activity results in impaired oral tolerance and increased intestinal inflammation, pointing to IDO as a possible new target of inflammatory bowel disease. Indeed, pharmacological targeting of IDO during chronic intestinal inflammation like IBD would be beneficial in dampening the inflammatory process and tissue damage in the gut.
Acknowledgments
We would like to thank Simona Ronzoni and Chiara Saccomani for excellent assistance with cell sorting; and Muriel Moser for IDO1-deficient mice.
References
Supplementary materials
Web only data for 59:5;595
Files in this Data Supplement:
Footnotes
Funding This study was supported by the Crohn's and Colitis Foundation of America (to MR), by the European Research Council (to MR) and by the Italian Association for Cancer Research (to GM).
Competing interests None.
Ethics approval All experimental procedures using human cells complied with guidelines set by our review board after informed consent from the patients (European Institute of Oncology, Milan). Experiments using mice were performed according to the Principles of Laboratory Animal Care guidelines (directive 86/609/EEC) and approved by the Italian Ministry of Health.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Digest