Article Text

Abstract
Background: The Cdx genes are expressed in the colorectal epithelium and are frequently downregulated during tumorigenesis. Overexpression of Cdx genes has been shown previously to result in cellular differentiation.
Aim: To study expression of CDX2 in normal and neoplastic human colon using a newly isolated monoclonal antibody. To define expression of CDX1 and CDX2 in an in vitro model system of colorectal tumour progression and to ascertain whether these are subject to regulation during differentiation.
Methods: Normal and neoplastic human colon was immunostained for CDX2. CDX1 and CDX2 expression was assayed in cell lines derived from premalignant colonic adenomas by western blotting. Differentiation was induced by sodium butyrate treatment or post confluent growth, and changes in CDX expression compared with carcinoma cell lines with low levels of CDX expression.
Results: CDX2 protein displayed no gradient of expression within the colonic crypt. Cell lines derived from adenomas, with high levels of CDX1 and CDX2, showed no regulation of these proteins when induced to differentiate by butyrate or confluency. CDX expression in these cell lines was independent of their APC or Ras status. CDX1 and CDX2 were expressed at very low levels in some carcinoma cell lines and were modestly upregulated on differentiation but were not restored to levels seen in adenoma cells.
Conclusion: The lack of significant regulation on cellular differentiation and the absence of a detectable gradient in the crypt implies that CDX2 may confer tissue specificity but may not play the previously suggested role in crypt patterning.
- CDX1
- CDX2
- colon
- cancer
- differentiation
- APC, adenomatous polyposis coli protein
- FAP, familial adenomatous polyposis
- DMEM, Dulbecco's modified Eagle's medium
- FBS, fetal bovine serum
- PBS, phosphate buffered saline
Statistics from Altmetric.com
- APC, adenomatous polyposis coli protein
- FAP, familial adenomatous polyposis
- DMEM, Dulbecco's modified Eagle's medium
- FBS, fetal bovine serum
- PBS, phosphate buffered saline
The human Cdx1 and Cdx2 genes encode transcription factors and constitute mammalian homologues of the Drosophila homeobox containing gene Caudal. In adult mammals, expression of these genes is largely restricted to the epithelium of the gut. They are expressed throughout the small and large intestines with the anterior limit occurring at the beginning of the duodenum.
Both genes have been reported to exhibit a gradient of expression with Cdx1 being present at the highest level caudally whereas Cdx2 expression peaks in the proximal colon and decreases caudally.1–,4 Numerous studies have also reported variation in expression of CDX1 and CDX2 along the crypt/villus or crypt/cuff axis.1,3,5,6Cdx mutant mice display an anterior homeotic shift7,8 and Cdx1 is expressed in intestinal metaplasia of the stomach and oesophagus.5 Taken together these data suggest that Cdx genes may confer positional information to the epithelium along the rostral-caudal axis of the gut.
Cdx2 heterozygote mice develop polyp-like lesions in the proximal colon which are distinct in their pathology from the more common adenomas of the colonic epithelium.8,9 These polyps, which do not express CDX2, display intercalary growth. They contain areas of heterotopic gastric and small intestinal mucosa and are reported to have their origins in embryogenesis.9
Several in vitro studies have reported that overexpression of Cdx2 in gut epithelial cells leads to a period of growth arrest accompanied by upregulation of several markers associated with intestinal differentiation.10,11 Furthermore, Mallo and colleagues12 showed that cotransfection of HT29 with Cdx1 and Cdx2 led to greater inhibition of growth than with Cdx2 alone (Cdx1 alone having no significant effect). This retardation of growth was associated with reduced resistance to apoptosis and also decreased tumorigenicity. Transfection of IEC-6 cells with Cdx1 increased resistance to apoptosis, enhanced growth, and promoted expression of differentiation markers in the absence of any Cdx2 expression.13
CDX2 has been demonstrated to induce transcription of several genes associated with differentiation of the intestinal epithelium, implying a key role for this protein in tissue specific transcriptional regulation (reviewed in Freund and colleagues14).
Expression of both Cdx genes is often reduced in colorectal cancer.5,15–,17Cdx2 expression has been found to decrease with tumour grade in human cancers, and also in rat and mouse models of the disease.8,15 Ee and colleagues15 reported comparative levels of CDX2 in adenomas displaying low grade dysplasia and the surrounding normal tissue whereas adenomas with high grade dysplasia show a reduction in CDX2 expression compared with normal tissue. In the majority of cases, this decrease cannot be attributed to mutation of the Cdx alleles,18 although small deletions within both Cdx2 alleles have been observed in a replication error (RER+) human colorectal cancer.19
Colorectal cancer remains a major cause of cancer deaths in western society. As the colonic epithelium is in direct contact with the luminal contents, dietary factors may influence epithelial turnover in this tissue. Short chain fatty acids are produced in the large bowel as a result of fermentation of dietary fibre/resistant starches by intestinal microflora. Butyrate is used as an energy source20 and may be present in the lumen in concentrations of 10–20 mmol.21 Studies using both colorectal adenoma and adenocarcinoma cell lines have shown butyrate to inhibit cell proliferation and upregulate markers of intestinal differentiation such as E-cadherin and alkaline phosphatase while inducing extensive apoptosis.22–,29
Because of the potential importance of CDX1 and CDX2 in the maintenance of the colonic epithelium and reduction in their expression in colorectal cancer, we sought to define and develop an in vitro model system for their study. Previous in vitro studies have used carcinoma derived cell lines. In this study we examined expression of CDX1 and CDX2 in six human colorectal adenoma derived cell lines. We also examined expression of CDX2 in vivo using a newly isolated monoclonal antibody on sections of normal mucosa from various sites along the rostral-caudal axis and in tumour specimens of different pathology. Furthermore, as overexpression of Cdx genes has been reported to drive differentiation processes in intestinal epithelial cells we wished to see if induction of differentiation in adenoma and carcinoma cells by butyrate, a luminal factor, and post confluent growth lead to regulation of CDX expression.
MATERIALS AND METHODS
Immunohistochemistry
Preparation of the mouse monoclonal antibody 7C7/D4 (BioGenex, San Ramon, California, USA) against human CDX2 has been described elsewhere.30 Tissue fixation and staining were carried out as described previously.30 In tumour specimens, the intensity of CDX2 nuclear staining in neoplastic epithelial cells was compared with that in adjacent normal epithelial cells.
Cell lines and culture conditions
AA/C1 is a clonogenic adenoma cell line derived from a 3–4 cm polyp from the descending colon of a familial adenomatous polyposis (FAP) patient.31 BH/C1 is a clonogenic variant of a line derived from pooled adenomas (all <0.5 cm in diameter and from throughout the colon) from a single FAP patient.31 AN/C1 is a clonogenic variant of a line derived from a sporadic colonic adenoma of 1–2 cm in diameter from the descending colon.32 BR/C1 is a clonogenic variant of a line derived from a sporadic sigmoid colon adenoma of 3–4 cm in diameter.33 RR/C1 is a clonogenic variant of a line derived from a sporadic rectal adenoma of <1 cm in diameter.33 All of the above lines are cultured in conditioned medium, as described by Williams and colleagues.34 RG/C2 is a clonogenic cell line derived from a sporadic tubular adenoma of the sigmoid colon of 1–2 cm in diameter and is cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 20% (v/v) fetal bovine serum (FBS).32 All of the adenoma derived cell lines are anchorage dependent and are non-tumorigenic in athymic nude mice. HCA7 was established from a moderately well differentiated mucinous carcinoma of the colon35 and was a kind gift from Dr Sue Kirkland (London, UK). HCA7-col29 was subcloned from the parental line36 and will be hereafter referred to as HCA7. HT29 was derived from a sporadic colonic adenocarcinoma.37 HCA7 and HT29 were cultured in DMEM supplemented with 10% (v/v) FBS, and all cell lines were cultured as adherent cells in 25 cm2 tissue culture flasks.
Western blot analysis
Samples of 1×106 cells were prepared and western blotted, as described by Williams and colleagues.39 E-cadherin was detected using HECD-1 antiserum which was provided by Professor Massimo Pignatelli (Bristol, UK) and the CDX1 antibody used was a kind gift from Dr Debra Silberg (Philadelphia, USA). CDX2 was detected using a polyclonal rabbit antiserum. This anti-serum was generated by immunising a rabbit with a purified recombinant fusion protein in which CDX2 amino acids 2–194 were fused to glutathione-S-transferase sequences. This region of CDX2 lacks the sequence specific homeobox domain and shares very limited similarity with other mammalian caudal related proteins (for example, CDX1). Blots were subsequently probed with anti-α-tubulin (Sigma, UK) to show equal sample loading.
Flow cytometry
Samples of 2×106 cells were washed in phosphate buffered saline (PBS) and permeabilised by incubation on ice for 10 minutes in 0.1% Triton X-100. Following PBS washes, cells were stained with CDX2 antibody for one hour at 4°C with constant agitation. Stained cells were washed three times in wash buffer (PBS containing 5% FBS and 0.1% sodium azide) and then incubated for one hour at 4°C in the presence of an FITC conjugated antirabbit IgG (Sigma). Following a further three washes, cells were resuspended in wash buffer and analysed on a Becton Dickinson FACScan flow cytometer. Analysis was carried out using laser excitation at 488 nm and a 525 nm bandpass filter for FITC fluorescence.
RESULTS
CDX2 is evenly expressed along the crypt-cuff axis of the colonic crypt at various sites along the proximal-distal axis of the colon
Figure 1⇓ shows the results of immunohistochemical analysis of sections taken from human ascending (fig 1A⇓) and descending (fig 1B⇓) colon and rectum (fig 1C⇓) using a mouse monoclonal antibody raised against CDX2. It is representative of more than 12 sections obtained from different regions of the large bowel (tissues ranging from the caecum to the rectum of seven patients) and shows clearly expression of CDX2 in all epithelial cells. No significant differences in staining intensity were noted along the axis of the crypt, regardless of the original rostral-caudal position of the tissue.

CDX2 is evenly expressed along the proximal-distal axis of the colon and the crypt-cuff axis of the colonic crypt. Immunohistochemical analysis of sections of human colorectal mucosa derived from the ascending (A) or descending (B) colon or rectum (C) using a monoclonal CDX2 antibody.
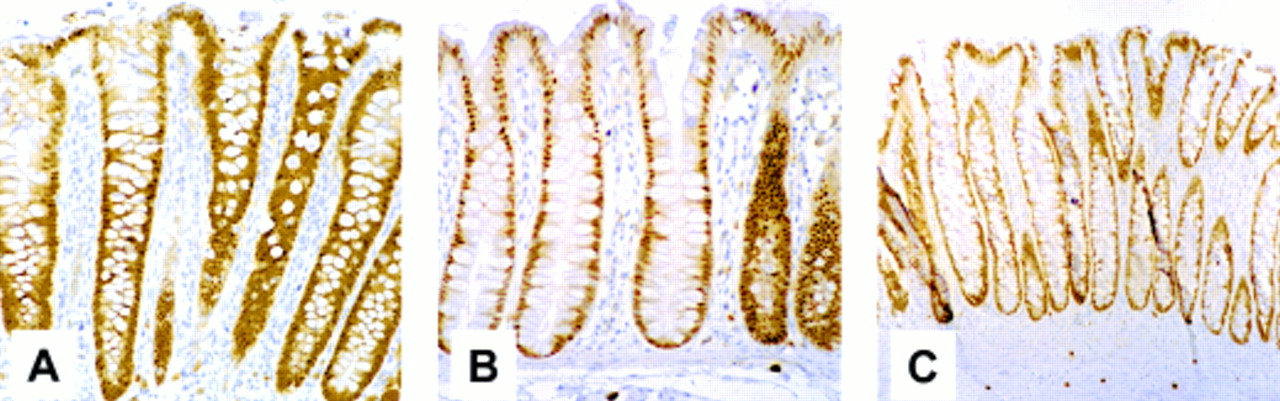
Several types of colon tumour display uniform epithelial expression of CDX2
Figure 2⇓ shows representative fields of view of sections of colon tumours stained for CDX2. The epithelial components of all three tumour types stained uniformly for CDX2. The well differentiated carcinoma (fig 2C⇓) showed equivalent staining between the normal epithelium (N) and the tumour component (T). Similar findings of essentially equivalent expression of CDX2 in normal and neoplastic epithelial cells were seen in the majority of more than 25 well differentiated colorectal carcinomas studied (Hinoi and colleagues30; Hinoi and Fearon, unpublished observations). It should be noted that although this section is representative of well differentiated carcinomas, poorly differentiated carcinomas were frequently found to contain reduced levels, or showed an absence of CDX2,30 as previously reported by others (reviewed in Freund and colleagues14).

Several types of colon tumour display even epithelial expression of CDX2. Immunohistochemical analysis of sections of human colorectal tumours derived from a tubular (A) or villous (B) adenoma and well differentiated carcinoma (C), using a monoclonal CDX2 antibody. Normal colonic mucosa (N) can be seen to the right of the carcinoma (T) in (C).
CDX1 and CDX2 are expressed at higher levels in colonic adenoma than colon carcinoma derived cell lines
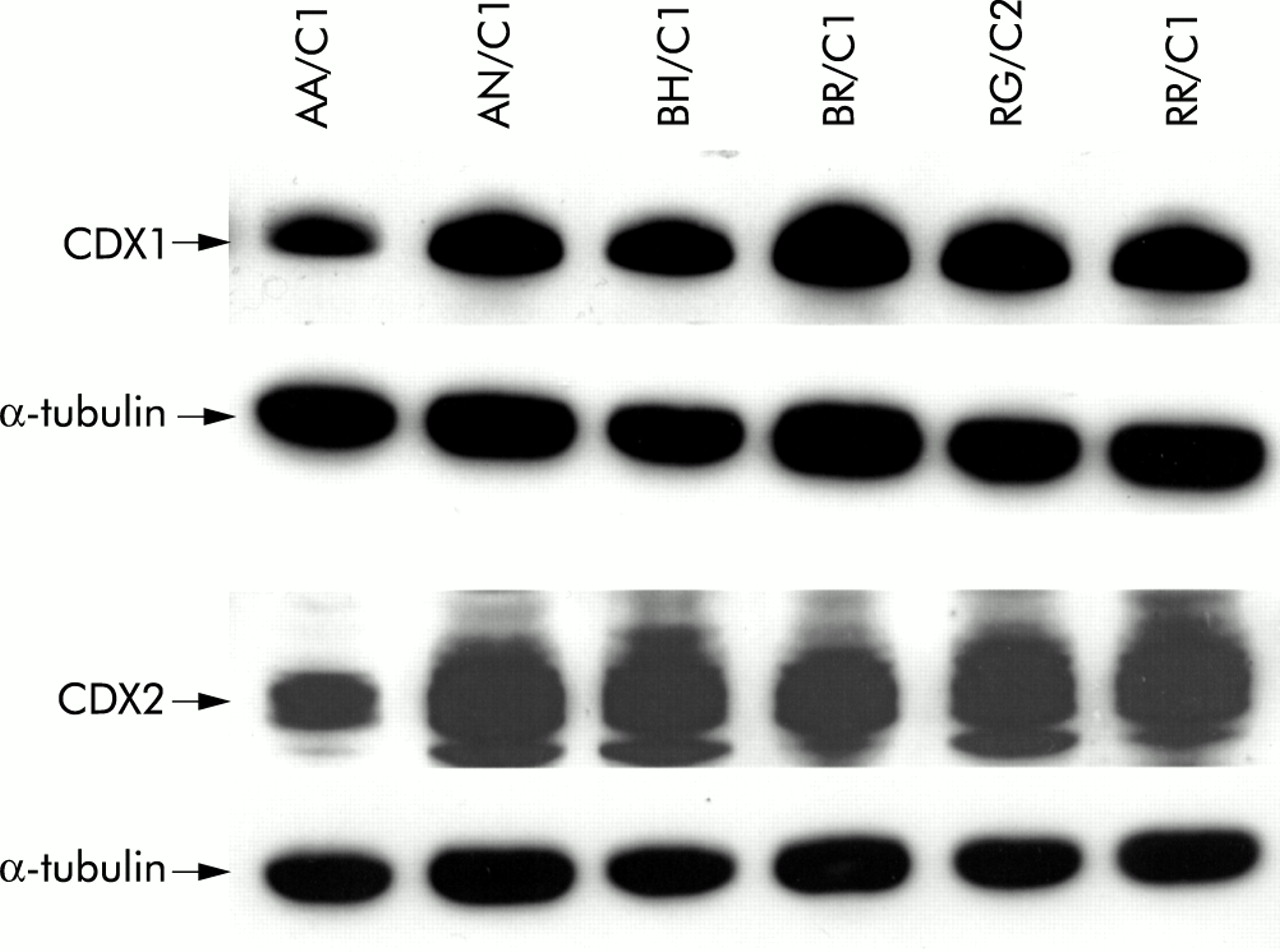
We assessed relative levels of expression of CDX1 and CDX2 in colonic adenoma and carcinoma derived cell lines. The CDX1 antibody detects a single band of approximately 33 kDa whereas the CDX2 antibody gives a doublet of 38–40 kDa in between two non-specific bands of unknown origin. Figure 3⇓ shows expression of both CDX proteins in six adenoma derived cell lines. All six lines showed expression of both CDX1 and CDX2. Levels of expression detected were comparable with the exception of AA/C1 which showed slightly lower levels than the other lines studied.
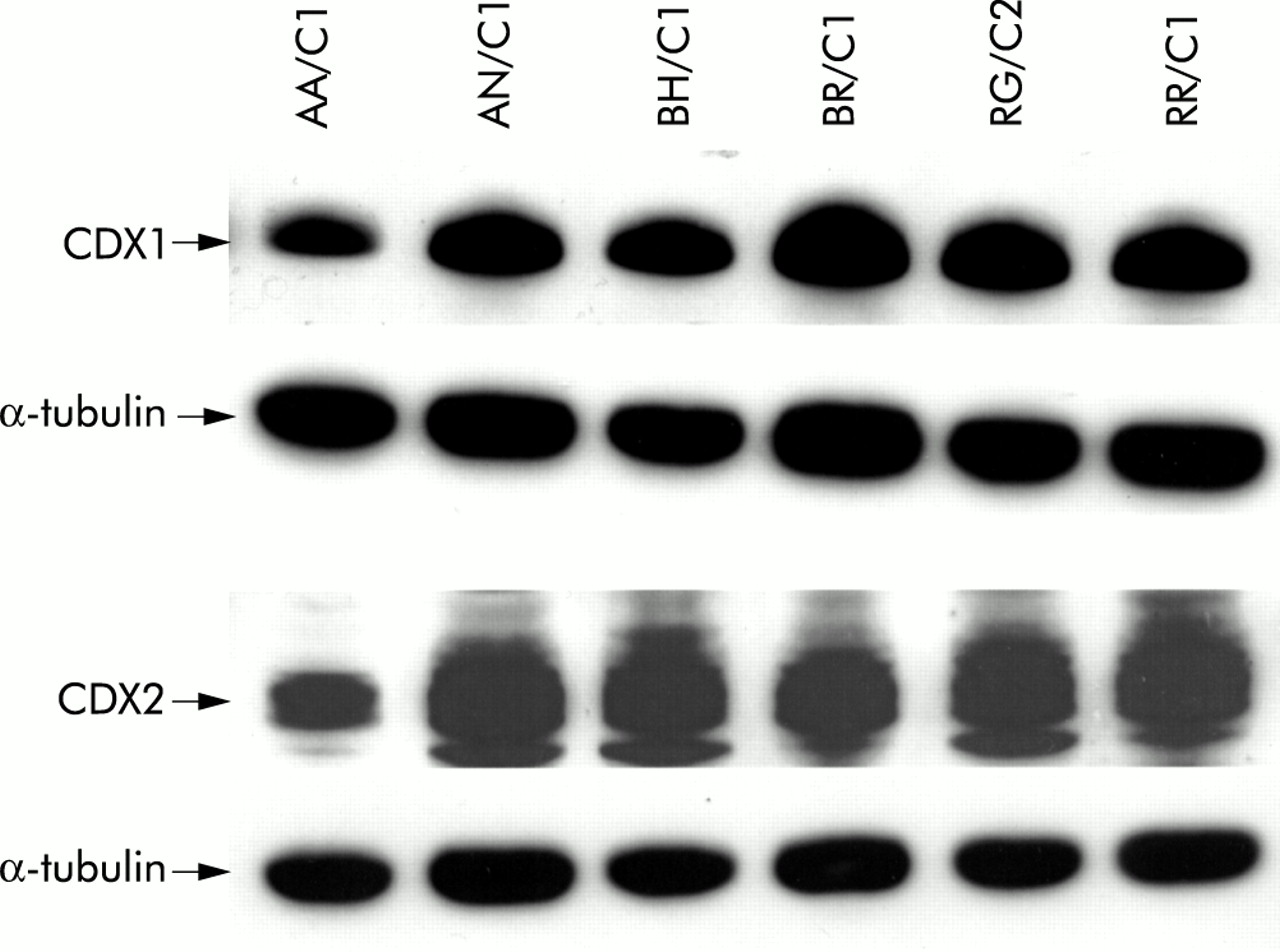
CDX1 and CDX2 were expressed at high levels in colonic adenoma derived cell lines. Western blot analyses of six adenoma derived cell lines for expression of CDX1 and CDX2 using rabbit polyclonal antibodies. The CDX1 antibody shows a single band of approximately 33 kDa whereas the CDX2 antibody shows a doublet of approximately 38–40 kDa in between two non-specific bands of unknown origin. Also shown is the corresponding reprobing of the blot with α-tubulin as a loading control (see control lanes in fig 5A⇓ for comparison with low levels of CDX1 and CDX2 detected in the carcinoma cell lines HCA7 and HT29).
We also studied the colorectal carcinoma cell lines HCA7 and HT29. By comparing the control lanes in fig 5A⇓, these carcinoma lines show levels of CDX1 and CDX2 expression that were much lower than in the adenoma cells. HT29 cells have previously been reported to contain very low levels of CDX212,16,17 and no prior study has reported on these proteins in HCA7.
Experiments were carried out to rule out the possibility that the difference in CDX expression between the adenoma and carcinoma derived cell lines was due to serum levels in the medium which were optimised for cell growth. The AA/C1 and RG/C2 adenoma lines expressed high levels of both CDX1 and CDX2, irrespective of whether they were cultured in the presence of 10% or 20% FBS. Similarly, HT29 and HCA7 expressed very low levels of the two proteins when cultured in 10% or 20% FBS (data not shown).
Sodium butyrate treatment does not regulate CDX1 and CDX2 in adenoma derived cell lines but produces a modest upregulation of both proteins in carcinoma cells under conditions resulting in growth inhibition and differentiation
Having established levels of CDX1 and CDX2 expression in adenoma and carcinoma derived cell lines, we sought to determine whether these levels were subject to regulation during differentiation. For this purpose we selected the AA/C1 and RG/C2 adenoma derived cell lines as induction of differentiation of these cells by butyrate has been reported previously.29 We also studied differentiation of HCA7 and HT29 cells, both of which demonstrate very low levels of CDX expression.
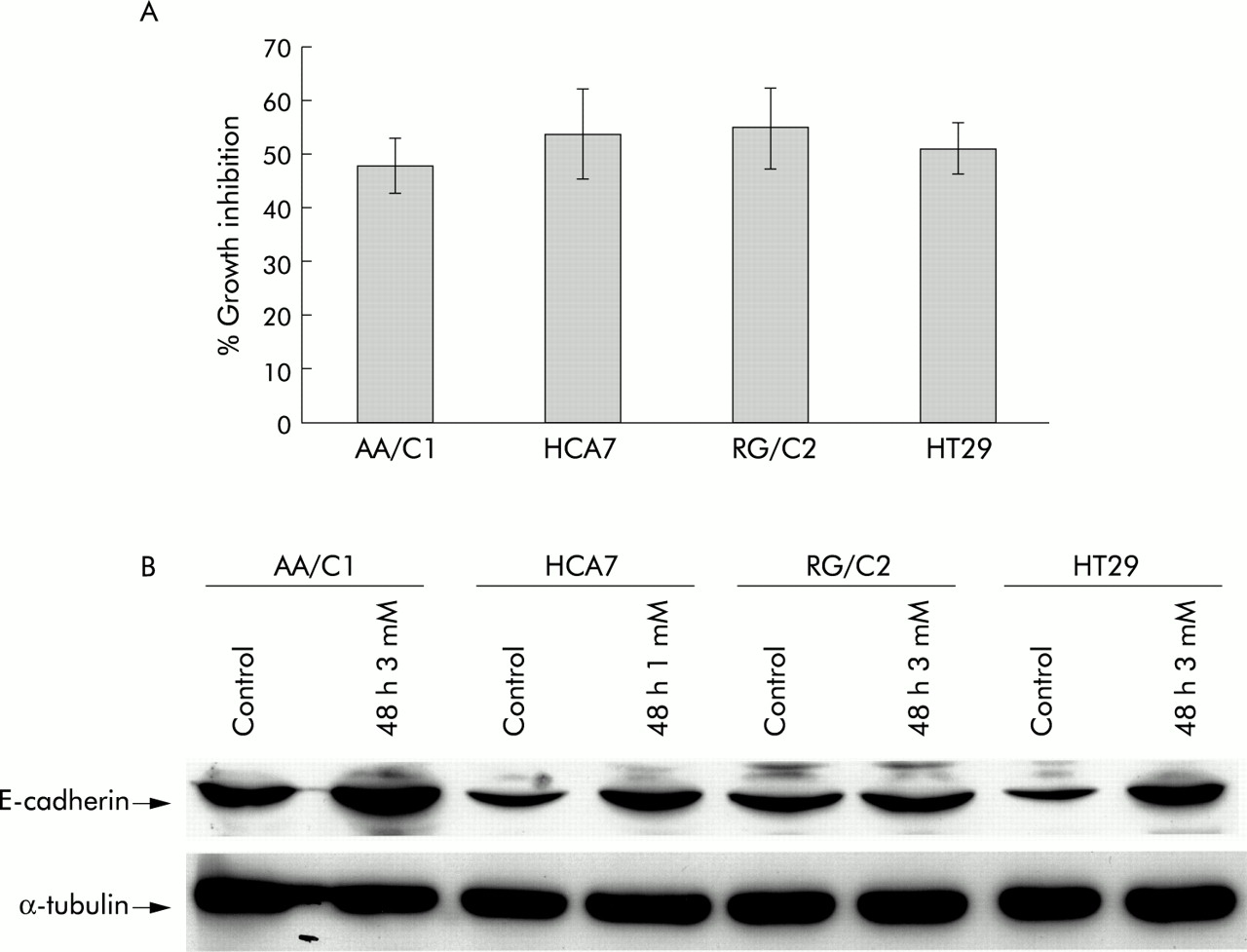
Each of the four cell lines were treated with a dose of sodium butyrate sufficient to result in approximately 50% growth inhibition over a 48 hour time period (see fig 4A⇓) and samples for western blotting prepared for comparison to appropriate vehicle controls. HCA7 cells demonstrated a much higher sensitivity to butyrate treatment and a lower dose (1 mM) was used to standardise the experiments. The reasons for this heightened sensitivity to butyrate are unclear.
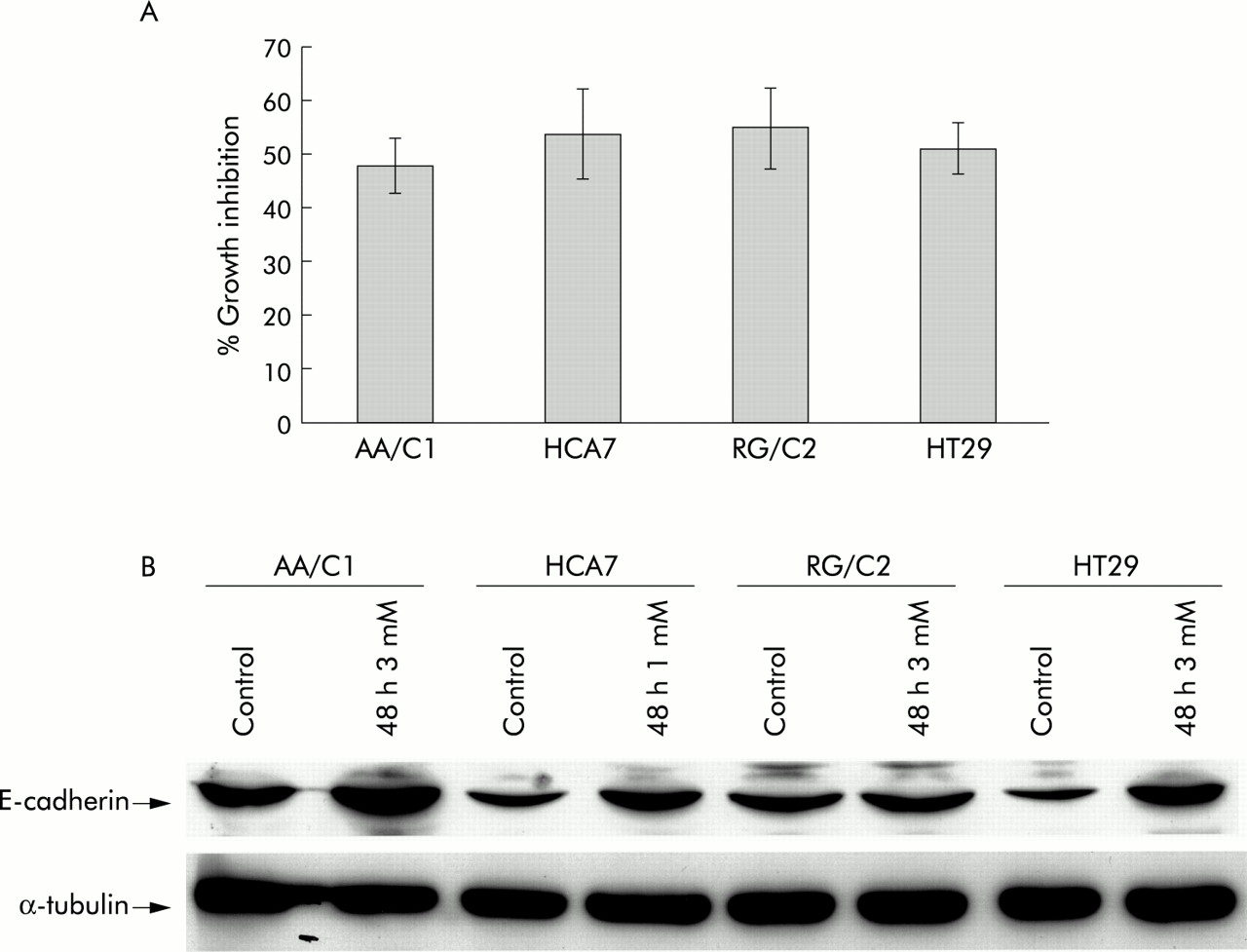
Effects of butyrate treatment on cell growth and E-cadherin. (A) Effect of 48 hour treatment with sodium butyrate on the cell populations in the adenoma and carcinoma derived cell lines AA/C1, HCA7, RG/C2, and HT29. Results shown are means (SEM) of three experiments and show the attached cell yield expressed as a percentage of the control cultures. (B) Effect of 48 hour treatment with sodium butyrate on E-cadherin expression in adenoma and carcinoma derived cell lines. The HECD-1 antibody recognised the E-cadherin protein (120 kDa). Also shown is the corresponding α-tubulin blot to show equal loading. This figure is representative of triplicate experiments.
Differentiation has been assessed in earlier studies in AA/C1 and RG/C229 and in HT2927,40 in terms of induction of E-cadherin expression and increased alkaline phosphatase activity. In this study we have confirmed induction of differentiation by assaying E-cadherin expression (fig 4B⇑). Induction of E-cadherin in HCA-7 by butyrate has not been reported previously. The level of E-cadherin peptide is clearly elevated by butyrate treatment in AA/C1, HCA7, and HT29, indicative of differentiation associated events taking place. This regulation is not as marked in the RG/C2 cell line but this observation is in agreement with other work29 in which upregulation of alkaline phosphatase was observed under the same conditions.
Using these conditions under which growth inhibition and differentiation occur, we studied expression of CDX1 and CDX2 by western blotting (fig 5⇓). No detectable change in the level of CDX proteins was observed for either of the adenoma lines following butyrate treatment (fig 5A⇓), even at lower exposure levels. The carcinomas showed a slight but reproducible increase (when performed six times) in levels of both CDX proteins following butyrate treatment, detectable if the western blots were exposed for longer (fig 5B⇓). This stimulated expression was however still much lower than basal levels observed in untreated adenoma cells (see RG/C2 in fig 5B⇓ for comparison). Thus although this butyrate treatment resulted in growth inhibition and differentiation, CDX1 or CDX2 expression was not restored in the carcinoma derived cells to levels seen in the adenoma cell lines.

Effect of 48 hour treatment with sodium butyrate on CDX1 and CDX2 expression in adenoma and carcinoma derived cell lines. (A) Effect of 48 hour treatment with sodium butyrate (1 or 3 mM) on expression of CDX1 and CDX2 using rabbit polyclonal antibodies in the adenoma and carcinoma derived cell lines AA/C1, HCA7, RG/C2, and HT29. The CDX1 antibody shows a single band of approximately 33 kDa whereas the CDX2 antibody shows a doublet of approximately 38–40 kDa in between two non-specific bands of unknown origin. Also shown are the corresponding reprobings of these blots with α-tubulin as a loading control. This blot is representative of those obtained when the experiment was performed six times. (B) Higher exposure of the CDX1 blot in (A) shows moderate regulation of CDX1 with butyrate treatment in HCA7 and HT29. Also included is the RG/C2 control at the same level of exposure for comparison.
Post confluent cell growth results in upregulation of CDX1 and CDX2 protein in HT29 but not in adenoma cell lines
In addition to induction with sodium butyrate, we also studied the effect of post confluent cell growth on CDX1 and CDX2 expression in the adenoma lines AA/C1 and RG/C2, and the carcinoma line HT29. AA/C1 has been demonstrated to exhibit “doming” when post confluent,38 which is associated with functional differentiation. Domes appear as the cells differentiate into columnar epithelial cells which can transport fluid from their luminal surface through their basal surface ensuring survival of underlying cells.35 HT29 cells show morphological features of enterocytic differentiation when maintained in a confluent state.41 The HCA7-col29 cells were not used for this study as they grow as a monolayer and do not demonstrate “doming”, unlike many other subclones of the same parental line.36
Cells were grown to a post confluent state and displayed the morphological changes associated with functional differentiation of the epithelial monolayer and their CDX expression assessed by western blotting (fig 6⇓). No change was observed in levels of either of the CDX proteins in the adenoma lines AA/C1 and RG/C2 (fig 6⇓). HT29 cells demonstrate upregulation of both CDX1 and CDX2 over time in post confluent culture to levels similar to those observed with butyrate treatment These experiments were carried out six times and the levels detected were consistent. However, this increase was not to levels observed in premalignant adenoma cells and also in this instance only the upper band of the CDX2 doublet appeared to be upregulated. Therefore, as with butyrate treatment, induction of differentiation seemed to occur in the carcinoma cells in the absence of restoration of CDX expression to levels seen in adenomas.

Effect of post confluent growth on CDX1 and CDX2 expression in adenoma and carcinoma derived cell lines. The effect of post confluent growth on expression of CDX1 and CDX2 using rabbit polyclonal antibodies in the adenoma and carcinoma derived cell lines AA/C1, RG/C2, and HT29. The CDX1 antibody shows a single band of approximately 33 kDa whereas the CDX2 antibody shows a doublet of approximately 38–40 kDa in between two non-specific bands of unknown origin. Also shown are the corresponding reprobings of these blots with α-tubulin as a loading control. This figure is representative of six separate experiments.
Increased CDX2 expression observed in HT29 after butyrate treatment occurs uniformly across the cell population
James and colleagues3 reported CDX2 staining to be heterogeneous in vivo. To assess the distribution of CDX2 expression throughout the cell population, HT29 cells were treated with butyrate as before and subsequently stained with the CDX2 antibody and, following staining with an FITC conjugated secondary antibody, were analysed by flow cytometry (fig 7⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Increased expression of CDX2 in butyrate treated HT29 occurred uniformly across the cell population. (A–C) Dot plots of side scatter (SSC) against forward scatter (FSC) to show the analysed population. (D–F) Histograms of counts over fluorescence log. The darker peaks represent the staining control (that is, second antibody only). The hollow peaks represent the stained samples, and geo mean fluorescence is included to show the difference in staining levels between the two cell lines. Untreated RG/C2 were included as a positive staining control.
The study of HT29 cells showed a distribution of fluorescence across the population without any sizeable subpopulations of brightly fluorescing or negative cells (fig 7D⇑). Treatment with butyrate significantly altered the light scattering characteristics of the cells (fig 7A and 7B⇑), but not the fluorescence characteristics (fig 7E⇑) in that the distribution of CDX2 staining appeared unchanged. The changes observed in light scatter with butyrate treatment are due to changes in cell size and complexity/granularity. Therefore, it appears that butyrate stimulates slight upregulation of CDX2 throughout the HT29 population and is not selectively regulating CDX2 in a subpopulation of cells.
Untreated RG/C2 were included in this experiment as a staining control. The much higher level of fluorescence detected in these cells is in agreement with our western blotting observations in showing that RG/C2 express much higher levels of CDX2 than HT29 (fig 5⇑).
DISCUSSION
The cell lines derived from colonic adenomas arguably represent a more “normal” cell type than carcinoma cells, which have been used extensively for studies of CDX1 and CDX2. Adenoma derived lines are, for example, non-tumorigenic and anchorage dependent, and retain sensitivity to the growth inhibitory effect of transforming growth factor β.29,42 Studies in vivo have shown that adenomas with low levels of dysplasia contain CDX2 levels comparable with the surrounding tissue.15 Expression of CDX1 and CDX2 has not been studied previously in premalignant adenoma cell lines. In this study, six adenoma lines were shown to express both proteins at roughly uniform levels. Furthermore, we have shown that the epithelial component of both tubular and villous adenoma specimens display evenly distributed expression of CDX2. The AA/C1 adenoma cell line exhibits a lower level of both CDX proteins than the other adenoma lines studied although still much higher than the carcinomas. The AA/C1 and AN/C1 lines were both derived from the descending colon and this positioning within the tissue does not seem to influence levels in the six adenoma cell lines studied. Furthermore, it seems unlikely that this reduced expression in AA/C1 is due to the FAP origin of this polyp as the BH/C1 cell line (also FAP) displayed a level of expression comparable with that seen in lines derived from sporadic polyps. The colon carcinoma cell lines HCA7 and HT29 expressed barely detectable levels of the Cdx gene products. This has been demonstrated previously for HT2912,16,17 but not for HCA7.
We also investigated the possibility that CDX expression may be regulated in these cells in response to differentiation inducing stimuli. The question also arises as to whether the luminal content of the colon may influence differentiation of the epithelium through effects on CDX1 and CDX2 expression. The ability of sodium butyrate to induce differentiation in colorectal cancer cells is well established (reviewed by Augenlicht and colleagues43). Post confluent growth has also been demonstrated to result in functional differentiation of colonic epithelial cells.35,38,41
Under conditions in which differentiation was induced either by butyrate treatment or by post confluent growth, we demonstrated a modest reproducible upregulation of CDX1 and CDX2 in two carcinoma cell lines exhibiting low levels of CDX expression. This upregulation is too slight to draw any definitive conclusions but appears to occur throughout the population as opposed to the appearance of a high expressing subpopulation. However, it is notable that levels of CDX1 and CDX2 in carcinomas were not restored to those seen in adenoma cells which are considered to represent a more normal cell type. Some colon carcinoma cell lines have been shown to express markers of small intestinal differentiation such as lactase and sucrase isomaltase. As CDX expression has been shown to be lower in the small intestine compared with the colon (see Silberg and colleagues44 and references therein), this phenotype may be due to reduced levels of CDX expression in these cell lines. Furthermore, polyps formed in the intestinal mucosa of Cdx2 heterozygote mice display the appearance of more rostral tissue phenotypes.9
Adenomas which contain relatively high levels of CDX1 and CDX2 showed no obvious regulation of either with butyrate treatment or confluency. It would seem therefore that dramatic changes in the levels of CDX1 and/or CDX2 are not required for cellular differentiation. Similarly, the relatively uniform expression of CDX2 along the crypt axis we observed in vivo suggests that CDX2 may act only to confer tissue specificity and it is possible that a graded expression is not necessary to pattern the crypt.
James and colleagues3 reported a gradient of CDX2 protein expression within the crypts of the distal colon with higher levels being detected in the upper section of the crypt. Conversely, they showed that mRNA for Cdx2 was located in the lower part of the crypt. In the proximal colon they reported staining of all epithelial cells, with no gradient of expression along the crypt axis.3 Our immunohistochemical studies using a new mouse monoclonal concurs with this uniform expression in the proximal colon but we saw no evidence of differential CDX2 expression along the axis of the crypt in tissue from the descending colon or rectum.
This finding is supported by the relative lack of CDX2 regulation during differentiation of our in vitro model system. Furthermore, work by Heerdt and colleagues27 has shown that butyrate induced differentiation and apoptosis are accompanied by an increase in alkaline phosphatase activity, a marker of differentiation. Notably, cells that have detached from the culture substrate after butyrate treatment, which are thought to represent terminal differentiation, have increased levels of alkaline phosphatase compared with their attached counterparts. Interestingly, we have found that these “terminally differentiated” floating cells rather than have higher levels of CDX2 were found to have lower levels of CDX2 than the attached cells when assessed by western blotting (Qualtrough and Paraskeva, unpublished results). Again, our in vitro system where we did not detect an increase in CDX2 expression with differentiation is consistent with our in vivo data showing uniform CDX2 expression in the crypt.
The mechanism by which CDX expression is lost or downregulated in colorectal tumours is not currently clear. A study by da Costa and colleagues45 showed that restoring expression of wild-type adenomatous polyposis coli protein (APC) induced expression of CDX2 in HT29. Interestingly, five of the six adenoma cell lines studied here have been shown not to express any full length APC (the exception being RG/C2)46 yet cells strongly express CDX2. It would appear therefore that loss of APC function alone is insufficient to reduce CDX2 expression to levels seen in, for example, the carcinoma line HT29.
Lorentz and colleagues47 reported that overexpression of oncogenic ras caused downregulation of Cdx2 and an increase in Cdx1 expression in CaCo2 and HT29. However, four of the six adenoma derived lines used in this study harbour ras mutations,33,48 yet strongly express CDX2. Thus it appears that ras activation alone is insufficient to downregulate Cdx2.
In conclusion, the premalignant adenoma derived cell lines we have studied provide a model for elucidating the role of CDX1 and CDX2 in the control of colonic epithelial growth and differentiation.
Cells of this type are much more representative of normal tissue than carcinoma derived cells and can be classified within the genetic model of colorectal carcinogenesis previously proposed49,50 and are therefore a useful tool for elucidating the relationship between CDX expression and tumour progression. Use of this model may lead to a better understanding of the biological consequence of this event during tumour progression and also the mechanism(s) of Cdx downregulation. Modest stimulation of CDX expression in carcinoma cells by butyrate highlights the potential influence of the luminal environment on tissue homeostasis in the colon but also that restoration of CDX expression is not required for differentiation of these cells in vitro.
Furthermore, our demonstration of uniform CDX2 expression along the length of the crypt in vivo and lack of CDX2 regulation in differentiating adenoma cells suggest that this gene is not responsible for the patterning of the crypt and driving the cellular differentiation process.
Acknowledgments
This work was supported by grants from Cancer Research UK and the NIH (USA).