Article Text

Abstract
Background and aims: Factors that induce luminal bacteria to cross the intestinal epithelium following injury remain poorly defined. The aim of this study was to investigate the interaction between glutamine metabolism, energy supply, and inflammatory mediators in determining the translocation of non-pathogenic bacteria across cultured enterocytes.
Methods: The effect of tumour necrosis factor α (TNF-α) on translocation of Escherichia coli C25 across Caco-2 epithelial monolayers was studied in the presence of products and inhibitors of glutamine metabolism. Simultaneous measurements of transepithelial electrical resistance (TEER) and flux of lucifer yellow were used to assess effects on the paracellular pathway. Lactate dehydrogenase release was used to monitor enterocyte integrity. Imaging of monolayers in these experimental conditions was undertaken with transmission electron microscopy.
Results: Exposure to basolateral TNF-α (20 ng/ml) for six hours induced translocation of E coli across Caco-2 but only if accompanied by simultaneous glutamine depletion (p<0.01). Translocation was inhibited by addition of glutamine for two hours (p<0.01) but not by an isonitrogenous mixture of non-glutamine containing amino acids. Inhibition of glutamine conversion to α-ketoglutarate, but not blockade of glutathione or polyamine synthesis, also induced translocation in the presence of TNF-α. Manipulations that induced bacterial translocation were associated with a marked reduction in enterocyte ATP levels. No effect of these treatments on paracellular permeability or lactate dehydrogenase release was observed. Conditions in which translocation occurred were associated with the presence of bacteria within enterocyte vacuoles but not the paracellular space.
Conclusions: In inflammatory conditions, the availability of glutamine as an enterocyte fuel substrate is essential for the preservation of a functional barrier to microorganisms. In conditions of acute glutamine depletion, cytokine mediated bacterial translocation appears to be primarily a transcellular process.
- intestinal barrier
- sepsis, cytokines
- Caco-2 cells
- glutamine
- tumour necrosis factor
- Gln, glutamine
- TNF-α, tumour necrosis factor α
- DMEM, Dulbecco’s modified Eagle’s medium
- FCS, fetal calf serum
- CFU, colony forming units
- HBSS, Hank’s balanced saline solution
- AOA, aminooxyacetate
- BSO, buthionine sulphoximine
- AKG, α-ketoglutarate
- DFMO, difluoromethylornithine
- TEER, transepithelial electrical resistance
- LY, lucifer yellow
- LDH, lactate dehydrogenase
Statistics from Altmetric.com
- Gln, glutamine
- TNF-α, tumour necrosis factor α
- DMEM, Dulbecco’s modified Eagle’s medium
- FCS, fetal calf serum
- CFU, colony forming units
- HBSS, Hank’s balanced saline solution
- AOA, aminooxyacetate
- BSO, buthionine sulphoximine
- AKG, α-ketoglutarate
- DFMO, difluoromethylornithine
- TEER, transepithelial electrical resistance
- LY, lucifer yellow
- LDH, lactate dehydrogenase
Glutamine is the most abundant free amino acid in the body1 and plays a key role in intermediary metabolism for rapidly dividing cells such as enterocytes and cells of the immune system. While the intestine derives a major portion of its energy requirements from glutamine oxidation,2 glutamine also acts as a key precursor for intestinal synthesis of the antioxidant glutathione3 and, in addition, stimulates intestinal synthesis of polyamines,4 which play a major role in the control of enterocyte proliferation and repair. Atrophy of the small intestinal mucosa that follows prolonged total parenteral nutrition can be prevented5 or reversed6 by addition of glutamine to intravenous feeding regimens. Glutamine is regarded as conditionally essential as, under conditions of stress, including trauma, sepsis, and ischaemia-reperfusion injury, the requirements for glutamine may exceed the capacity of the other tissues in the body to provide it.7
The barrier function of the gastrointestinal tract has been shown to be impaired following stress or inflammation.8 It has been suggested that this may play a role in the translocation of bacteria and bacterial products across the gut wall, leading to sepsis, a prolonged systemic inflammatory response, and multiple organ failure.9 Proinflammatory cytokines such as tumour necrosis factor α (TNF-α) are key mediators of the systemic inflammatory response and in critical illness states are released both systemically10 and locally in the gut.11 Moreover, TNF-α has been shown to have significant modulatory effects on the permeability of intercellular junctions in vitro.12 Administration of glutamine has been shown to reduce septic complications in multiply injured patients13 and to significantly reduce impairment in gut barrier function and bacterial translocation seen in both in vivo14,15 and in vitro16 models of critical illness and gut injury. The mechanism by which glutamine exerts these effects is presently unclear. Provision of the major fuel substrate for enterocytes under conditions of stress may underlie the potential therapeutic role of glutamine in gut barrier dysfunction. However, prevention of bacterial translocation could equally relate to the role of glutamine in the maintenance of gut glutathione or polyamine levels or even to a stimulatory effect on the immune system,17 as opposed to the gastrointestinal tract.
The aim of this study was to test the hypothesis that cellular glutamine availability plays a key role in the enterocyte in modulation of bacterial translocation, and to determine which of the glutamine metabolic pathways are involved in this effect. An in vitro model of cytokine mediated bacterial translocation was developed to allow direct study of the interaction between enterocytes, TNF-α, glutamine, and translocating bacteria without the potential confounding factors of immune cells, gut motility, intestinal blood flow, bacteria-bacteria interactions, and the intestinal mucus layer.
MATERIALS AND METHODS
Preparation of Caco-2 monolayers
Caco-2 cells (ATCC, Manassas USA, passage 100–120) were seeded at 2×105 cells/cm2 onto 24 mm Transwell (Corning Costar, USA) bicameral systems with polycarbonate membranes (3 μm pore size, 4.7 cm2 surface area). Monolayers were grown in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum (FCS, containing 0.1 mM glutamine (Gln)), 2 mM Gln, and 2 mM non-essential amino acids, with 5 IU/ml penicillin and 5 μg/l streptomycin (Gibco, Paisley UK). Cells were cultured at 37°C in a humidified 5% CO2/95% air atmosphere with a change of medium every 3–4 days. At the start of each experiment, DMEM containing 2 mM Gln was replaced with experimental media, as detailed below.
Measurement of electrical resistance and paracellular permeability
Monolayer integrity was determined by serial measurement of transepithelial electrical resistance (TEER) using an ohmmeter (Evometer World Precision Instruments, Sarasota, USA). Monolayers were used between 18 and 24 days after seeding, with a TEER of 219 (2) Ω/cm2 (mean (SD), n=124).18 Multilayering of Caco-2 cells was excluded with transmission electron microscopy (data not shown). TEER was recorded for each experiment before and after treatment with the different experimental media.
Paracellular permeability of the Caco-2 monolayers was assessed at the same time as bacterial translocation by measuring the flux of lucifer yellow (LY; Sigma-Aldrich, Poole, UK) from apical to basal chambers of the Transwell. LY is a fluorescent dye and was used in these experiments as a marker of paracellular permeability18 because it is not metabolised by Escherichia coli, in contrast with other permeability markers such as mannitol. Preliminary studies (data not shown) confirmed that coincubation with LY at the concentration used did not affect the growth characteristics or viability of E coli and that inoculation of solutions of LY with up to 108 colony forming units (CFU)/ml E coli did not affect the precision of the LY assay. LY (50 μM) was added to the apical chamber at the same time as bacterial inoculation. After four hours, LY concentration in the basolateral chamber was measured with an LS 50B luminescence spectrometer (Perkin-Elmer, Cambridge, UK) against a standard concentration curve (excitation 430 nm, emission 535 nm).
Preparation of bacteria
E coli C25, which has previously been shown to translocate across Caco-2 monolayers,19 was generously supplied by Dr E Deitch, (UMDNJ, New Jersey, USA). Nutrient broth No 2 (Lab M, Bury, UK) was inoculated with E coli C25 and incubated for 12 hours to plateau phase, producing a concentration of ∼108 colony forming units (CFU)/ml, which was verified by serial dilution culture on blood agar plates. Aliquots of 1 ml were then centrifuged at 5000 rpm for five minutes. The supernatant was removed and bacteria resuspended in Hank’s balanced saline solution (HBSS; Gibco, Paisley, UK), supplemented with 10 mM sodium bicarbonate and 180 mg/dl glucose, pH balanced to 7.4.
Measurement of bacterial translocation
All translocation measurements were conducted in HBSS to minimise the effect of bacterial growth on the count of translocated bacteria. HBSS reduced the growth of E coli C25 by approximately 1000-fold compared with DMEM, as evidenced by the marked increase in doubling time (21 (3) minutes in DMEM v 74 (6) minutes in HBSS; n=12). Preliminary studies (data not shown) indicated that monolayer TEER and LY permeability remained unaltered in HBSS for at least six hours. At the end of each incubation period, the experimental culture medium was removed by washing the monolayers twice with HBSS at 37°C. Monolayers were then allowed to equilibrate in HBSS for 30 minutes. Monolayers for which TEER had not returned to within 10% of the value before removal of experimental media were discarded. The apical chamber of the Transwell was then inoculated with E coli C25 to a final concentration of 108 CFU /ml. Bacteria were added together with LY to allow simultaneous estimation of bacterial flux and paracellular permeability. After a four hour incubation period (37°C; 5% CO2/95% room air), during which translocation of bacteria from apical to basal chambers of the Transwell was allowed to occur, the concentration of E coli in the basal chamber of the Transwell was determined by serial dilution in HBSS, followed by plating on blood agar and overnight culture in a 5% CO2 atmosphere at 37°C. Translocation of E coli was expressed as log CFU/ml in the basal chamber.
Microscopic examinations
Preparation of samples for microscopy
At the conclusion of incubations, membranes with adherent enterocyte monolayers were fixed with 2.5% glutaraldehyde in 0.1 mol/l sodium cacodylate buffer. After fixation, membranes were cut into 5 mm sections, washed again in 0.1 mol/l sodium cacodylate sucrose, and postfixed with 1% osmium tetroxide. Monolayers were then washed again in 0.1 mol/l sodium cacodylate, dehydrated in 100% ethanol, and embedded in Agar 100 resin (Agar Scientific, Essex UK) via propylene oxide. After polymerisation, the resultant blocks had ultrathin (60 nm) sections cut using an ultramicrotome (Reichert Ultracut, Vienna, Austria). Sections were mounted on 200 mesh copper grids, stained with uranyl acetate and lead citrate, and examined using either an AEI EM801 (Manchester, UK) or Philips CM10 (Eindhoven, Netherlands) electron microscope. Micrography was performed using Kodak electron microscope film 4489 (Anachem Ltd, Luton, UK).
Experimental protocols
Effect of glutamine availability on bacterial translocation
To study the effects of Gln availability and TNF-α on the translocation of E coli C25, Caco-2 monolayers were transferred for six hours to DMEM containing 2 mM Gln (ConGln) or no supplementary Gln (GlnF). GlnF medium thus contained only 0.1 mM Gln derived from FCS. For studies with TNF-α, monolayers were incubated in experimental media (see below) with addition of TNF-α (20 ng/ml; Sigma-Aldrich, Poole, UK) to the basolateral chamber. At the end of the six hour incubation period, bacterial translocation was measured as detailed above.
To assess reversibility and specificity of Gln effects, monolayers were first incubated with basolateral TNF-α (20 ng/ml) in GlnF DMEM for six hours. This was followed by replacement of Gln (2 mM) in the apical chamber of the Transwell. In a separate experiment to determine the specificity of Gln mediated effects, a 2 mM isonitrogenous mixture of non-Gln containing amino acids was added (non-essential amino acids; Gibco, Paisley UK: alanine 0.89 mg/ml, asparagine 1.32 mg/ml, aspartic acid 1.30 mg/ml, glutamic acid 1.47 mg/ml, glycine 0.75 mg/ml, proline 1.15 mg/ml, and serine 1.05 mg/ml). In each case, incubation was continued for a further two hours after which the experimental medium was removed, monolayers washed in HBSS, and bacterial translocation measured in HBSS as indicated above.
Metabolic pathways involved in glutamine effects on bacterial translocation
To determine the biochemical mechanisms underlying the effect of Gln on bacterial translocation, addition of Gln to the apical chamber was repeated in the presence of inhibitors of key steps of Gln metabolism. Aminooxyacetate (AOA) is an aminotransferase inhibitor preventing oxidative Gln metabolism within the citric acid cycle by blocking its conversion to α-ketoglutarate (AKG).4 Buthionine sulphoximine (BSO) is a gamma glutamylcysteine synthetase inhibitor which prevents conversion of Gln into the antioxidant glutathione.20 Difluoromethylornithine (DFMO) is an ornithine decarboxylase inhibitor that prevents glutamine stimulated synthesis of polyamines.21
In these studies, monolayers were first incubated with basolateral TNF-α (20 ng/ml) for six hours in GlnF DMEM. Following this, 2 mM Gln in isolation, or together with 600 μM AOA, 3.2 mM BSO (Sigma-Aldrich, Poole, UK), or 5 mM DFMO (Calbiochem, Nottingham, UK) were added to the apical chamber and incubation continued for two hours after which bacterial translocation was measured.
To exclude a toxic effect of AOA, an additional group of monolayers was incubated in GlnF DMEM with basolateral TNF-α (20 ng/ml) for six hours. Following this, 600 μM AOA, together with either 2 mM Gln or 2 mM AKG (Sigma-Aldrich) were added to the apical chamber, and incubation continued for a further two hours, before monolayers were washed with HBSS and bacterial translocation measured as indicated above.
Finally, the effects of AOA, BSO, and DFMO on bacterial translocation in the absence of TNF-α were assessed. Each agent was added to monolayers incubated in ConGln DMEM for six hours, prior to replacing the medium with HBSS and undertaking studies of bacterial translocation.
Lactate dehydrogenase release (LDH)
LDH release from Caco-2 monolayers was measured as a marker of cellular integrity. Enterocyte monolayers were exposed to the treatments indicated above and extracellular fluid collected and centrifuged at 15 000 g for 15 minutes. The supernatant was then assayed using a colorimetric Boehringer Mannheim LDH cytotoxicity detection kit (Roche, Welwyn Garden City, UK) at 490 nm absorbance in a Thermomax microplate reader (Molecular Devices, Wokingham, UK). Total intracellular LDH levels were obtained by incubating cells with 2% Triton X100 (Sigma-Aldrich, Poole, UK) and measuring LDH in the resulting cell lysate. LDH release is expressed as a percentage of Triton soluble intracellular levels from control monolayers.
Cellular ATP levels
To determine the effect of changes in Gln availability and exposure to TNF-α on cellular ATP content, monolayers were washed twice in HBSS followed by lysis with somatic cell ATP releasing buffer (Sigma-Aldrich, Poole, UK). ATP levels in the cell lysates were measured with an ATP determination kit (Molecular Probes, Cambridge, UK), using a standard curve of known ATP concentration, at 560 nm emission in a 1450 Microbeta luminometer (Perkin-Elmer, Cambridge, UK). Cellular protein content was measured using a BCA protein assay (Pierce Inc., Rockford, USA) at 570 nm against a standard curve of known albumin concentration. ATP levels were expressed as pmol ATP/μg protein.
Statistical analyses
Results are expressed as mean (SD) unless otherwise stated, for at least six monolayers in each group. Data were analysed by ANOVA with post hoc Dunnett’s or Bonferroni tests as appropriate.22 A p value of ≤0.05 was accepted as the level of statistical significance. All analyses were undertaken using Graphpad Prism software (Graphpad Prism Inc, San Diego, California, USA).
RESULTS
TNF-α and glutamine depletion cause a synergistic increase in bacterial translocation
Incubation of monolayers with ConGln DMEM was associated with low rates of translocation of E coli C25 to the basal chamber, equivalent to less than 0.01% of the original apical inoculum. Acute withdrawal of Gln in the apical chamber for six hours (GlnF) did not influence the rate of bacterial translocation (fig 1) nor did it affect the integrity of the monolayers, as indicated by LDH release, LY permeability, and TEER (table 1).
Effect of incubation media on markers of monolayer integrity and cellular ATP content
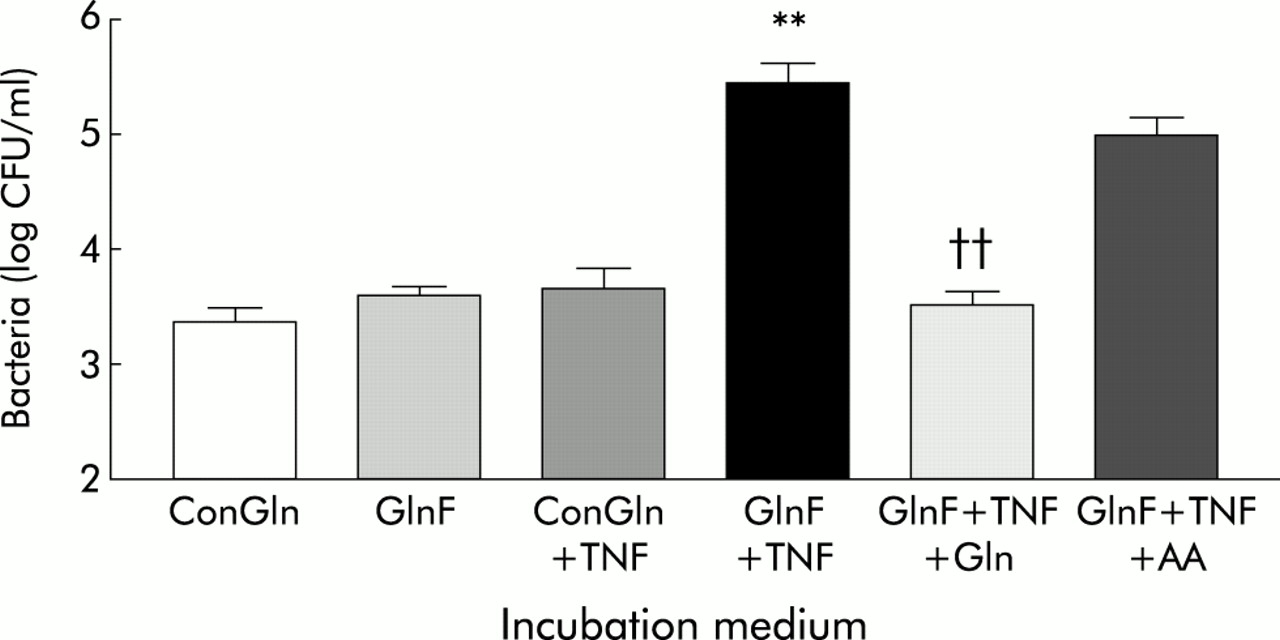
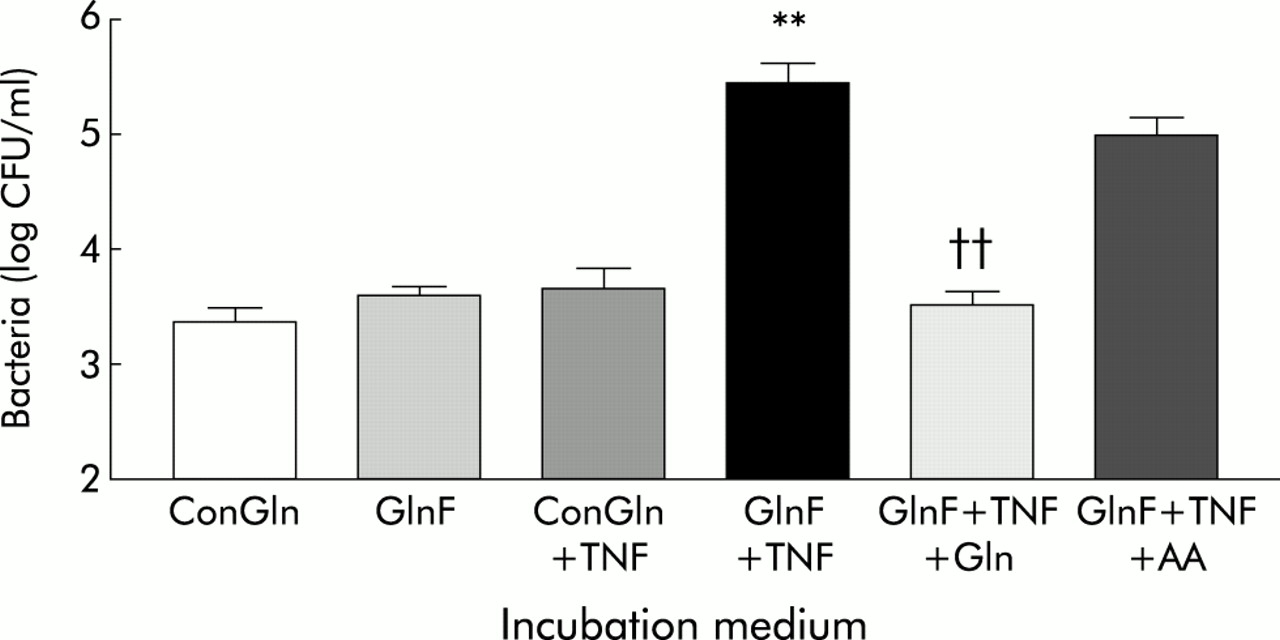
Effect of glutamine availability and tumour necrosis factor α (TNF-α) on translocation of E coli C25 across Caco-2 monolayers. ConGln, Dulbecco’s modified Eagle’s medium (DMEM) containing 2 mM glutamine; GlnF, glutamine free DMEM; TNF, coincubation with 20 ng/ml TNF-α in the basal chamber of Transwell; GlnF+TNF+Gln, two hours replacement of Gln or an isonitrogenous mixture of other amino acids (AA). Results are mean (SEM) of six separate experiments. **p<0.01 compared with all other groups except GlnF+TNF+AA; ††p<0.01 compared with GlnF+TNF and GlnF+TNF+AA.
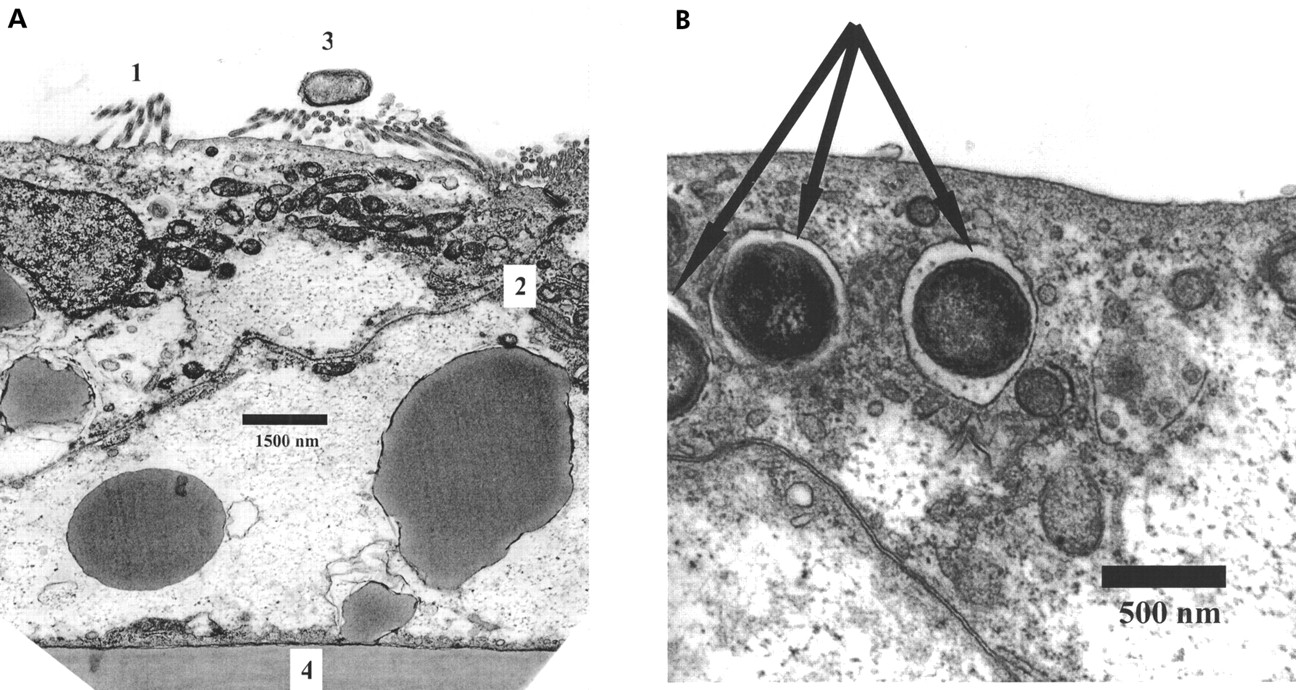
Basolateral addition of TNF-α for six hours to monolayers in ConGln also failed to affect bacterial translocation (fig 1), LDH release, LY permeability, or TEER (table 1). In contrast, basolateral addition of TNF-α to monolayers incubated in GlnF medium markedly increased (100-fold) the rate of bacterial translocation from 3.38 (0.25) to 5.48 (0.43) log CFU/ml (p<0.01) (fig 1). These changes were however not associated with alterations in monolayer integrity (table 1). The increase in bacterial translocation that occurred following incubation of monolayers with TNF-α in GlnF DMEM was completely inhibited by addition of Gln for two hours. This effect appeared to be specific to Gln as adding other amino acids failed to prevent the increase in bacterial translocation (fig 1). In neither case were the changes in bacterial translocation associated with alterations in monolayer integrity (table 1). Electron microscopy of monolayers in which bacterial translocation had occurred consistently demonstrated bacteria in vacuoles within enterocytes (fig 2).

Transmission electron micrographs of Caco-2 monolayers. (A) Microvilli (1) and tight junctional complex (2). Escherichia coli (3) are clearly seen on the apical surface of the monolayer, which overlies the polycarbonate membrane (4). Scale as indicated. (B) Caco-2 cell showing E coli (arrowed) within intracellular vacuoles. Scale as indicated.
Metabolic pathways involved in effects of glutamine depletion on C-25 translocation
Addition of AOA, BSO, or DFMO alone to monolayers in ConGln without addition of TNF-α was unable to induce increased bacterial translocation (fig 3). However, incubation of monolayers in ConGln and TNF-α caused a dramatic increase in bacterial translocation after addition of AOA (but not BSO or DFMO), similar in magnitude to that resulting from TNF-α exposure in GlnF DMEM (fig 3).

Effect of inhibitors of glutamine metabolism on translocation of E coli C25 across Caco-2 monolayers in the presence of 2 mM glutamine (ConGln), with or without TNF-α 20 ng/ml (TNF). AOA, aminooxyacetate; BSO, buthionine sulphoximine; DFMO, difluoromethylornithine. Results are mean (SEM) of four separate experiments. **p<0.01 versus other groups.
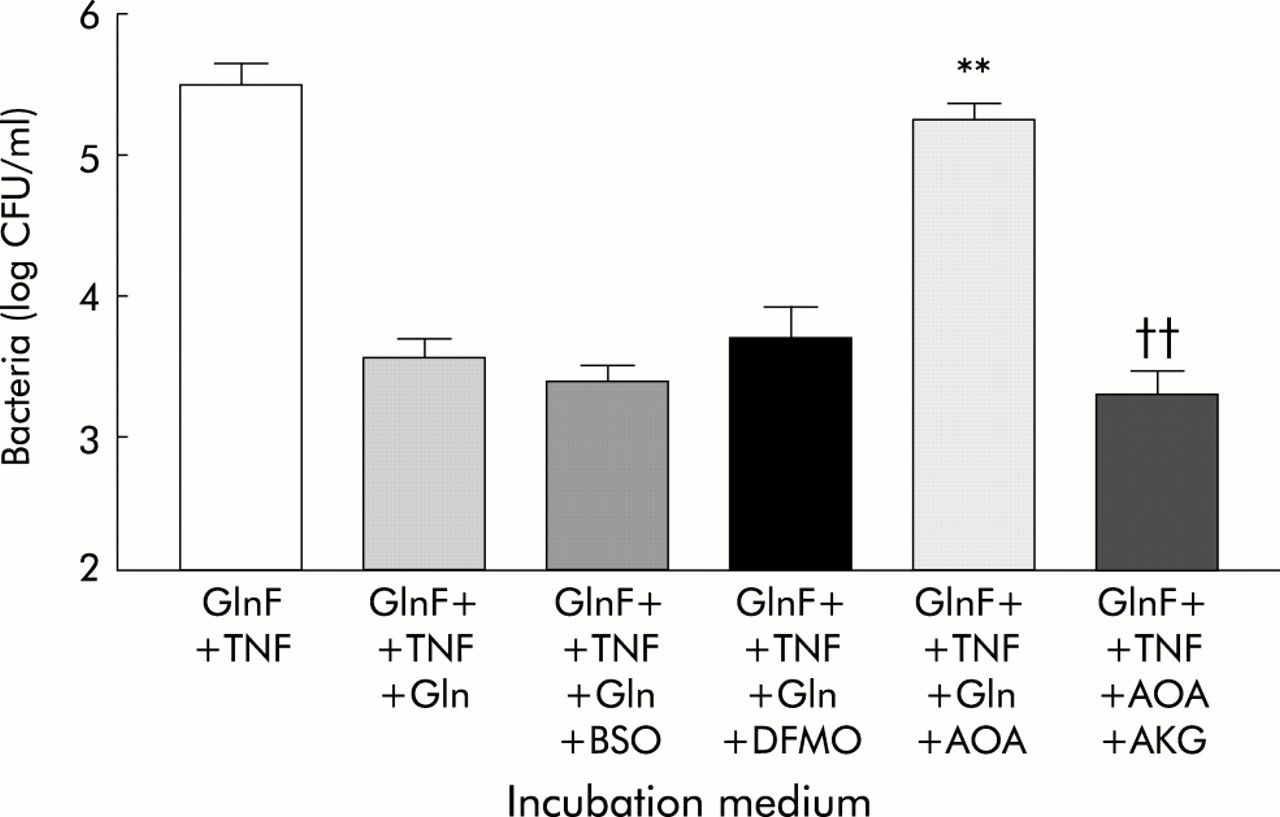
Restoration of 2 mM Gln for two hours inhibited the increased translocation observed after incubation of monolayers in GlnF DMEM and basolateral TNF-α (fig 4). The ability of Gln to reverse the increased bacterial translocation associated with Gln deprivation and TNF-α exposure was blocked by coincubation with AOA, but not by coincubation with BSO or DFMO (fig 4). Despite the presence of AOA however, bacterial translocation under these conditions was inhibited by the addition of AKG (fig 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of inhibitors of glutamine metabolism on glutamine mediated reversal of bacterial translocation across Caco-2 monolayers incubated in glutamine free Dulbecco’s modified Eagle’s medium (DMEM) (GlnF), with basolateral 20 ng/ml TNF-α, with addition of: Gln, 2 mM glutamine; AOA, aminooxyacetate; BSO, buthionine sulphoximine; DFMO, difluoromethylornithine; AKG, α-ketoglutarate. Results are mean (SEM) of >4 separate experiments. **p<0.01 compared with: GlnF+TNF+Gln, GlnF+TNF+Gln+BSO, GlnF+TNF+Gln+DFMO; ††p<0.01 versus GlnF+TNF+Gln+AOA.
Cellular ATP levels
Glutamine deprivation, or exposure to TNF-α alone, had no effect on ATP levels (table 1). However, ATP levels in GlnF conditions were reduced by almost 50% after basolateral addition of TNF-α (104.3 (17.8) v 197.3 (11.1) pmol/μg protein; p<0.01). Incubation with AOA and TNF-α in ConGln medium reduced ATP levels to a similar degree. Addition of Gln to monolayers incubated in GlnF medium with basolateral TNF-α returned cellular ATP to control (ConGln) levels, which was also observed when AKG was added to monolayers incubated in ConGln which had been exposed to both AOA and basolateral TNF-α. Addition of Gln to monolayers incubated in ConGln medium and exposed to both AOA and basolateral TNF-α also significantly increased ATP concentration (table 1).
DISCUSSION
Although bacterial translocation was first postulated more than five decades ago,23 the mechanisms by which certain bacteria cross an apparently intact intestinal epithelium and the factors which modulate the process remain largely unknown. While great care should be taken in extrapolating the results of these in vitro studies to the clinical environment, in which a large number of confounding factors may be of equal or greater importance, the use of the present reductionist approach does however allow detailed exploration of bacteria-enteroctye interactions under strictly controlled conditions.24
Under basal conditions, a very small proportion (less than 0.01%) of the apical bacterial inoculum translocated to the basal chamber, a value in close agreement with previous reports of this experimental system.24 Neither removal of Gln from the apical medium nor incubation with TNF-α in the basal chamber alone for six hours significantly influenced the rate of bacterial translocation, TEER, or LY flux. These results are consistent with the findings of previous studies which have shown that Gln depletion for at least 48 hours is needed to reduce TEER or increase translocation of E coli in ileal loops.25 Although the short period of Gln depletion used in the present study seemed to have little functional consequences for the barrier function of the enterocyte monolayer under basal conditions, it did unmask a marked response of the monolayer to treatment with TNF-α. Although proinflammatory cytokines have previously been shown to alter the barrier function of enterocyte monolayers,26,27 the effect of changes in Gln availability on the function of enterocytes subjected to inflammatory mediators has not, to our knowledge, been studied.
The effect of TNF-α was examined in the present study because of evidence that suggests a pathogenetic role for this cytokine in the process of stress induced bacterial translocation. Experimental haemorrhagic shock induces both bacterial translocation28 and intestinal production of TNF-α29 and the two processes appear to be related both temporally and anatomically.30 TNF-α is also produced endogenously by enterocyte cell lines in response to E coli11 and hypoxic stress,26 and treatment of the human HT-29/B6 intestinal cell line with TNF-α at doses similar to those used in the present study (although for longer periods) has been shown to induce both structural changes in tight junctions and impairment of barrier function in cell lines.12 TNF-α has been shown to induce the production of reactive oxygen species,31 which may impair barrier function in enterocyte monolayers,32 via an effect on tight junctions.33 In the present study, neither short term exposure to TNF-α nor Gln depletion induced bacterial translocation but Gln depletion dramatically sensitised the Caco-2 monolayer to the effects of TNF-α. Glutamine concentrations of 2 mM were chosen in the present study because they represent the normal physiological glutamine concentration required for optimal growth and function of the Caco-2 cell line.18 The present study only examined the effect of varying Gln availability on the apical surface of the monolayer as previous observations have indicated a polarity for the effect of Gln on the integrity of the epithelial barrier, with apical effects predominating.25 It should be noted that the concentrations of Gln used in the present study were chosen because they reflect normal physiological conditions for Caco-2 cells.18 While even short term withdrawal of Gln supplementation from the incubation medium might be regarded as unphysiological, the concentrations of Gln employed in the unsupplemented state in the present study (that is, 0.1 mM) have previously been shown not to affect normal physiological function in Caco-2 cells.34
The effects of Gln deprivation are unlikely to be related to cellular injury as the effect was rapidly and specifically inhibited by Gln (but not by other amino acids) and increased LDH release was not observed. While Gln acts as the principal fuel substrate for the enterocyte,2 it may also serve as a metabolic precursor for glutathione,3,35 and stimulates the synthesis of polyamines,4 both of which may play a role in the maintenance of the integrity of the epithelial barrier.36
Several observations in the present study suggest however that the ability of glutamine to prevent bacterial translocation depends on its use as an enterocyte fuel substrate. AOA (which inhibits the conversion of Gln to AKG, a key step in oxidative metabolism) blocked the Gln mediated reversal of TNF-α induced bacterial translocation, while BSO (an inhibitor of glutathione synthesis) and DFMO (an inhibitor of polyamine synthesis) did not. A toxic effect of AOA is unlikely given the ability of AKG to reproduce the effect of Gln, even in the presence of AOA. Finally, TNF-α mediated bacterial translocation only occurred in the present studies when withdrawal of Gln or blockade of Gln metabolism by AOA was associated with a significant fall in cellular ATP levels. Glutamine has previously been shown to prevent the decline in cellular energy potential in the intestinal epithelium of burned rodents, without concomitantly increasing glutathione content.37 It is unclear why a decline in cellular energy availability in the present study was associated with translocation of bacteria and these findings appear partly to contradict those of a previous study in which an inhibitor of oxidative metabolism failed to affect the rate of E coli translocation in a similar experimental system, despite a reduction in TEER. Although Cruz and colleagues19 suggested that these findings indicate that cellular energy availability may not be of major importance in determining the rate of bacterial translocation, cytokine induced translocation was not studied, nor were ATP levels measured. In the present study, inhibition of oxidative glutamine metabolism by AOA treatment alone also failed to increase bacterial translocation unless cells were subjected to an additional stress, which suggests that multiple stimuli may be required to induce bacterial translocation in this model.
Glutamine depletion has been shown to lead to increased gut permeability during parenteral nutrition in both animals38 and humans,39 and exposure of cell lines to TNF-α may induce alterations in the structure and function of tight junctions12,40 and enterocyte apoptosis.41 Although apoptosis was not examined in the present study, it seems unlikely to have accounted for the increase in bacterial translocation because of the rapid (less than two hours) and complete reversal of the translocation that occurred following replacement of Gln in the medium. In addition, derangement of the enterocyte barrier as a consequence of apoptosis would have been expected to have resulted in an increase in paracellular permeability41 which was not observed in the present study, despite a 100-fold increase in the rate of bacterial translocation. Finally, Fas induced apoptosis in a similar experimental model has been shown to selectively increase permeability to small molecules,42 and the converse was observed in the present study.
The aim of this study was to examine the mechanisms by which glutamine modulates bacterial translocation and not to define the route by which translocation occurs. However, the striking lack of association in all of the experiments in the present study between the increase in bacterial translocation and changes in TEER or LY flux indicates that the paracellular pathway was not the principal route for the passage of bacteria across the Caco-2 monolayer. This observation lends further support to the suggestion that bacterial translocation may involve transcellular,19,43 rather than paracellular,44 passage of bacteria. This suggestion is further supported by the finding of translocating bacteria in vacuoles within enterocytes and not within the paracellular space. While these observations appear to differ from those reported by Wells and colleagues,45 who noted transcellular passage of a variety of invasive pathogens in Caco-2 cells, although not E coli C25, bacteria were only visualised within enterocytes in the present study during conditions in which increased translocation also occurred (for example, after glutamine depletion and treatment with TNF). As the study of Wells and colleagues45 was conducted under basal conditions, as opposed to during enterocyte stress, it seems likely that the frequency of bacteria-enterocyte interactions, for an organism with low inherent invasiveness such as E coli C25, was too low for bacteria to be seen within enterocytes under those circumstances, as opposed to those of the present study in which E coli C25 translocation increased 100-fold.
In the present study, increased bacterial translocation only occurred in circumstances in which ATP depletion was observed. It is unclear how a reduction in cellular ATP relates to bacterial translocation although it is unlikely that ATP depletion developed as a consequence of translocation as no bacteria were employed in studies in which ATP content was measured. Rather, our observations suggest instead that ATP depletion in some manner facilitated bacterial translocation. Inhibition of Na+/K+-ATPase has previously been shown to impair endocytosis of invasive pathogens into epithelial cells46 although prolonged Gln depletion, which induced a reduction in Na+/K+-ATPase activity, also increased transcytosis of bacteria.25 While Caco-2 cells have been shown to phagocytose live E coli, they do not appear to possess bactericidal activity47 and, while differences between bacterial strains, cell lines, and the experimental conditions employed could explain these variable findings, it is unclear whether the relationship between ATP depletion and increased bacterial translocation observed in the present study could be explained by reduced ATP dependent intracellular killing, particularly in view of the finding that addition of Gln in the presence of AOA and TNF-α increased cellular ATP levels without preventing the increase in translocation. In addition, although bacterial transcytosis is likely to be an energy dependent process, the relationship between cellular energy content and bacterial flux is unclear. Furthermore, the apical to basal flux of bacteria in the present studies may reflect the net effect of bacterial transport in both directions and it is presently unclear whether these pathways are similarly energy requiring. Further studies which characterise the precise mechanisms of translocation of E coli C25 in stressed epithelia and their relationship to cellular energy content may help explain and enhance the potential clinical benefits of enteral glutamine supplementation.
Acknowledgments
This work was supported by the Medical Research Council, the Royal College of Surgeons of England, the Peel Medical Research Trust, and the Digestive Disorders and Nutritional Research Foundations.