Article Text

Abstract
Background: Iron absorption increases during pregnancy to cater for the increased iron requirements of the growing fetus.
Aims: To investigate the role of the duodenal iron transport molecules and hepatic regulatory molecules in coordinating the changes in iron absorption observed during pregnancy.
Methods: Rats at various days of gestation and 24–48 hours post-partum were examined for hepatic expression of hepcidin, transferrin receptors 1 and 2, and HFE (the gene mutated in the most prevalent form of hereditary haemochromatosis), and duodenal expression of divalent metal transporter 1 (DMT1), duodenal cytochrome b (Dcytb), iron regulated mRNA (Ireg1), and hephaestin (Hp) by ribonuclease protection assay, western blotting, and immunohistochemistry.
Results: Decreased hepatic non-haem iron and transferrin saturation and increased expression of transferrin receptor 1 in the liver indicated a progressive reduction in maternal body iron stores during pregnancy. Duodenal expression of the iron transport molecules DMT1, Dcytb, and Ireg1 increased during pregnancy, and this corresponded with a reduction in hepcidin, HFE, and transferrin receptor 2 expression in the liver. Expression of all molecules returned towards control values by 24–48 hours post-partum.
Conclusions: These data indicate that increased expression of key iron transport molecules is responsible for the elevated iron absorption associated with pregnancy, and implicate hepcidin, HFE, and transferrin receptor 2 in determining how the maternal iron homeostatic machinery responds to the increased iron demands accompanying gestation.
- duodenal cytochrome b
- divalent metal transporter 1
- hereditary haemochromatosis
- iron regulation
- Dcytb, duodenal cytochrome b
- DMT1, divalent metal transporter 1
- GAPDH, glyceraldehyde 3-phosphate dehydrogenase
- HFE, the gene mutated in the most prevalent form of hereditary haemochromatosis
- Hp, hephaestin
- IRE, iron responsive element
- Ireg1, iron regulated mRNA (also known as ferroportin 1)
- NP, non-pregnant
- PP, post-partum
- RPA, ribonuclease protection assay
- TfR, transferrin receptor
Statistics from Altmetric.com
- Dcytb, duodenal cytochrome b
- DMT1, divalent metal transporter 1
- GAPDH, glyceraldehyde 3-phosphate dehydrogenase
- HFE, the gene mutated in the most prevalent form of hereditary haemochromatosis
- Hp, hephaestin
- IRE, iron responsive element
- Ireg1, iron regulated mRNA (also known as ferroportin 1)
- NP, non-pregnant
- PP, post-partum
- RPA, ribonuclease protection assay
- TfR, transferrin receptor
Iron requirements increase during pregnancy due to expansion of the maternal erythrocyte mass and high iron demand of the growing fetus. These requirements are met initially through mobilisation of maternal iron stores (principally from the liver) but as iron stores become depleted, intestinal iron absorption increases to maintain an adequate iron supply for both the mother and her offspring.1–3 The fetus obtains its iron via the placenta which sequesters transferrin bound iron from the maternal circulation. The rate of materno-fetal transfer increases with the increasing size of the fetus and placenta, and is maximal just prior to parturition.4,5 Iron absorption is also maximal at this time. For example, in humans, there is a ninefold increase in iron absorption between weeks 12 and 36 of gestation6 while in rats absorption increases substantially from day 16 through to term at day 21.1 Absorption rapidly returns to control levels post-partum (for example, within 18 hours in rats),7 indicating the strength of the absorption stimulus provided by pregnancy.
The ability of the small intestine to respond to the iron demands of pregnancy is indicative of a highly regulated process. Under normal physiological conditions the two main factors that regulate iron absorption are the level of body iron stores and rate of erythropoiesis.8 Iron absorption increases when stores are low and decreases when stores are high, so it is not surprising that the depleted body iron levels associated with pregnancy are associated with enhanced iron absorption. However, one study has found that iron loaded rats have increased absorption during pregnancy, in spite of their high iron stores.1 This observation suggests that the level of maternal iron stores may not be the sole regulator of iron absorption during pregnancy.
Recent advances in our understanding of the mechanism and regulation of intestinal iron transport have come from the discovery of a number of key molecules involved in the process. These include the iron transporters divalent metal transporter 1 (DMT1) and iron regulated mRNA (Ireg1; also known as ferroportin 1), and the enzymes duodenal cytochrome b (Dcytb) and hephaestin (Hp). The brush border transporter DMT1, putative basolateral transporter Ireg1, and the ferrireductase Dcytb are all upregulated under iron deficient conditions9 and these changes underlie the increase in iron absorption accompanying iron depletion. Disruption of the ceruloplasmin homologue Hp, a putative iron oxidase, leads to microcytic hypochromic anaemia in mice10 but this protein does not appear to be modulated significantly by variations in iron levels.11 Several other molecules have been implicated in determining how the body responds to changes in iron requirements, including HFE (the gene mutated in the most prevalent form of hereditary haemochromatosis), transferrin receptor 2 (TfR2), and the antimicrobial peptide hepcidin. These are all highly expressed in the liver and mutations in any of these genes can result in iron overload.12–14 Indeed, HFE is the gene most commonly mutated in the prevalent iron loading disorder hereditary haemochromatosis. Hepcidin is secreted into the circulation and has been proposed to be the messenger between body iron stores and iron absorption by duodenal enterocytes.15
There has been no comprehensive analysis of the molecular mechanisms underlying the enhanced iron absorption associated with pregnancy. We thus sought to investigate the role of the duodenal iron transport molecules and hepatic regulatory molecules in coordinating the changes in iron absorption observed as gestation progresses. These studies demonstrated a progressive increase in expression of iron transport molecules in the duodenum during pregnancy that corresponded with decreasing body iron stores and a reduction in liver hepcidin expression. Interestingly, expression of HFE and TfR2 also declined as pregnancy proceeded. After birth, all parameters returned towards control levels within two days.
METHODS
Animals
Female Sprague-Dawley rats fed a standard rodent pellet diet (370 mg iron/kg) and time mated at 10–12 weeks of age were obtained from the Herston Medical Research Centre, Brisbane, Australia. Rats were sacrificed at 9, 15, 18, and 21 days of gestation, and 24–48 hours post-partum. Those that failed to fall pregnant were used as non-pregnant controls. Rats were anaesthetised (44 mg/kg ketamine and 8 mg/kg xylazine intraperitoneally) and exsanguinated from the abdominal aorta to provide blood for serum iron determination. Samples of liver and duodenum were removed, and duodenal enterocytes were isolated as described previously.16 Tissues and pellets of isolated cells were snap frozen in liquid nitrogen. This study was approved by the Queensland Institute of Medical Research Animal Ethics Committee.
Serum and tissue iron analyses
Serum transferrin saturation was determined using an iron and iron binding capacity kit (Sigma-Aldrich, Sydney, Australia). Liver tissue was dried overnight at 110°C, extracted with acid, and non-haem iron content was determined using a colorimetric assay, as described previously.17
Ribonuclease protection assays
Gene expression in liver and enterocytes was analysed by ribonuclease protection assay (RPA) using 5 μg of total RNA isolated with TRIzol reagent (Invitrogen, Melbourne, Australia) as per the manufacturer’s instructions. Expression of the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and the iron metabolism genes Hp, HFE, hepcidin, Dcytb, Ireg1, transferrin receptor 1 (TfR1), TfR2, and DMT1 (both the iron responsive element (IRE) containing splice variant and the non-IRE form) were examined using riboprobes corresponding to the complementary DNA sequences of these genes, as previously described.9,11 The riboprobe for TfR2 corresponded to nucleotides 1859 to 2039 of the cDNA sequence listed in GenBank (accession No XM 22022). Band intensities were quantified by densitometry, and iron metabolism gene expression was reported as a proportion of GAPDH gene expression.
Western blot analysis
Protein was extracted from isolated duodenal enterocytes and DMT1 and Ireg1 protein expression examined by western blotting using polyclonal antibodies raised as previously described.9,11,18 Band intensities were quantified by densitometry, and expression of DMT1 and Ireg1 were reported as a proportion of actin protein expression to control for any discrepancies in gel loading.
Immunohistochemistry
Sections cut from duodenal tissue embedded and snap frozen in OCT embedding compound (Sakura Finetek, Torrance, California, USA) were used to localise DMT1 and Dcytb proteins by immunofluorescence microscopy, as previously described.9
Statistical analysis
All values are expressed as mean (SEM). Statistical differences between means were calculated using ANOVA and the Student’s t test, correcting for differences in sample variance (Microsoft Excel; Microsoft Pty Ltd, North Ryde, Australia).
RESULTS
As pregnancy progresses and fetal iron demand increases, there is a corresponding drop in maternal iron stores.3 In this study, hepatic non-haem iron levels decreased from 8.27 (0.9) μmol iron/g dry weight in non-pregnant control rats to 2.43 (0.5) μmol iron/g (p<0.01) on day 21 of gestation (fig 1A). Liver iron content increased slightly after birth to 3.80 (0.4) μmol Fe/g but was still well below control levels (p<0.05). Transferrin saturation progressively decreased throughout pregnancy from 45.1 (4.6)% in non-pregnant animals to 6.5 (0.8)% on day 21 (p<0.01) but returned to control levels by 48 hours post-partum (fig 1B). Corresponding with this reduction in liver iron concentration and transferrin saturation was an increase in the hepatic expression of TfR1 mRNA, as measured by RPA (5.9-fold over non-pregnant level on day 21; p<0.05) (fig 2). TfR1 expression had fallen by 24–48 hours post-partum but was still 3.5-fold above non-pregnant levels (p<0.01).

Hepatic non-haem iron and serum transferrin saturation during pregnancy. Rats were studied at various days of gestation and 24–48 hours post-partum (PP) and were compared with non-pregnant rats (NP). Values are reported as μmol non-haem iron/g dry weight liver tissue (A) and percentage saturation of serum transferrin (B). Data are expressed as means (SEM), n = 4. Significant difference from non-pregnant rats: *p<0.05, **p<0.01.
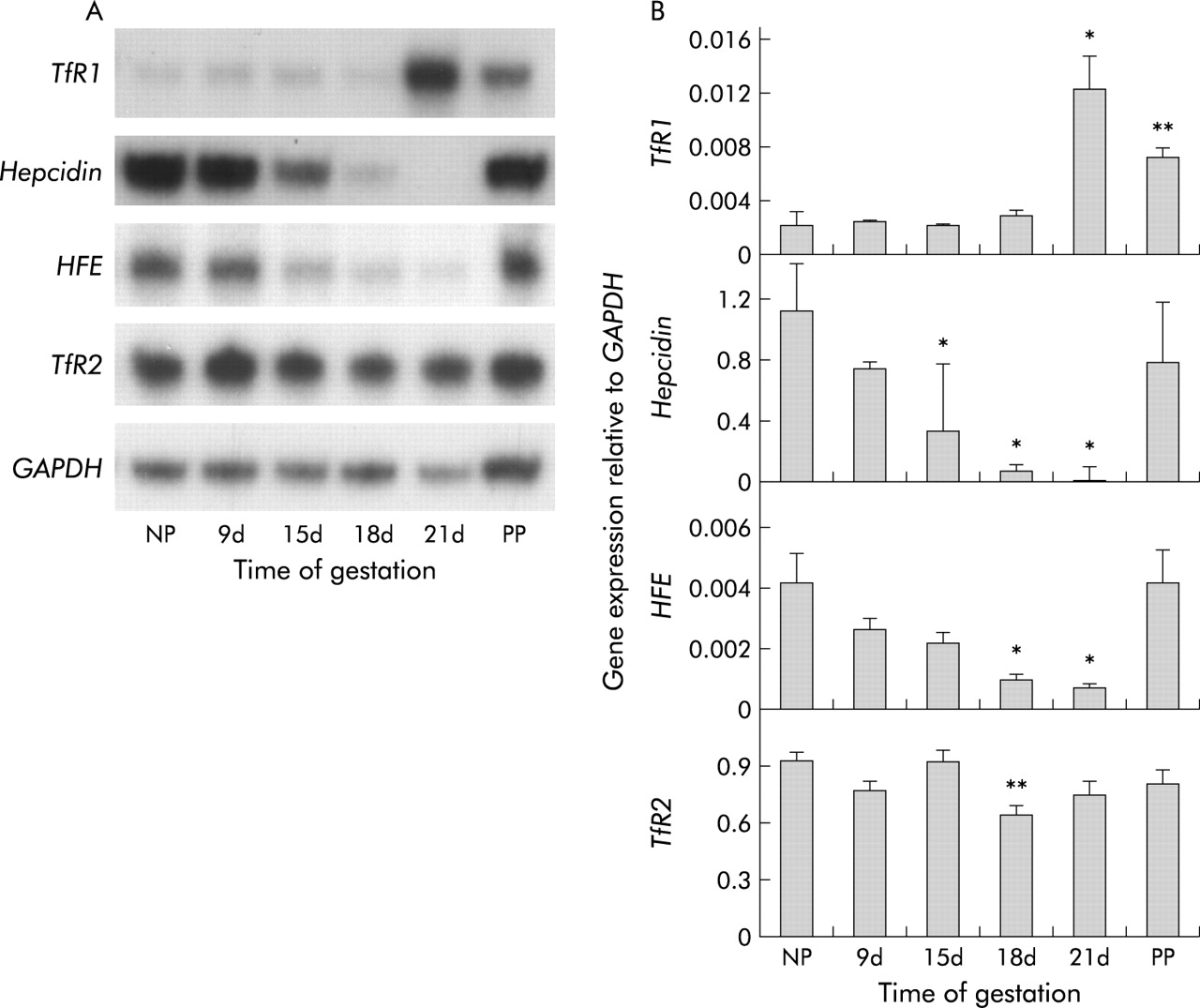
Hepatic expression of transferrin receptor 1 (TfR1), hepcidin, HFE (the gene mutated in the most prevalent form of hereditary haemochromatosis), and TfR2 mRNA. Animals are as described in fig 1. Expression of each gene was measured by ribonuclease protection assay. (A) Representative assays for each gene. (B) Band intensities for each gene were measured by densitometry and graphed as a proportion of glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Data are mean (SEM), n = 4. Significant difference from non-pregnant rats: *p<0.05, **p<0.01.
As recent studies have suggested that the hepatic molecules hepcidin, HFE, and TfR2 play key roles in the regulation of iron absorption, we investigated their expression during pregnancy. There was a progressive decline in hepcidin expression (fig 2) which reached statistical significance by day 15 and a minimum of 1.9% of the non-pregnant level (p<0.05) on day 21. After birth, hepcidin expression rapidly returned to near control levels. Hepatic expression of both HFE and TfR2 decreased as pregnancy progressed (fig 2). HFE expression followed a similar pattern to hepcidin and was significantly decreased to 18% of non-pregnancy levels (p<0.05 compared with non-pregnant controls) on day 21 before returning to control levels by 24–48 hours after birth. The reduction in TfR2 expression was less striking, with a decline to 70% of the control value on day 18 of pregnancy (p<0.01).
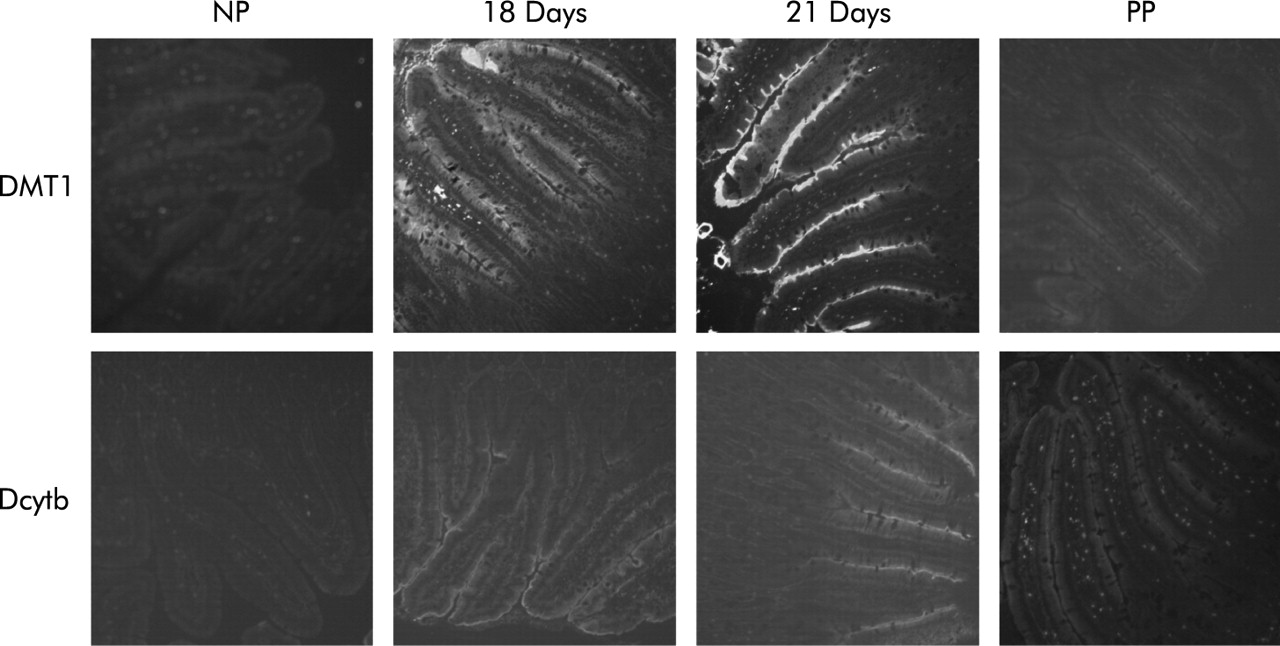
As an increase in maternal intestinal iron absorption has been well described during pregnancy,1,6 we investigated expression of duodenal iron transport molecules during gestation. Expression of mRNA for the brush border iron transporter DMT1 increased as pregnancy progressed (fig 3), peaking at 7.9 times (p<0.05) and 2.4 times (p<0.01) non-pregnant levels for the IRE and non-IRE forms, respectively, on gestation day 18, but declining thereafter. Western blot analysis showed DMT1 protein levels to be similarly upregulated (8.1-fold over non-pregnant control animals; p<0.05) on day 18 at the protein level and reduced to control levels by 24–48 hours post-partum (fig 4). Immunofluorescence showed an increase in DMT1 on the brush border on days 18 and 21 compared with both non-pregnant (fig 5) and 24–48 hour post-partum rats. Expression of the brush border ferrireductase Dcytb was also increased in the latter half of pregnancy. At day 15, mRNA expression rose to 2.9 times non-pregnant levels, remained relatively constant until day 21 (3.3-fold increase on day 18 and 3.1-fold increase on day 21 over non-pregnant animals; p<0.05) before declining after birth (fig 3). Immunohistochemistry showed little Dcytb staining in non-pregnant animals but distinct brush border staining on days 18 and 21. This was greatly reduced by 24–48 hours after birth (fig 5).

Duodenal expression of divalent metal transporter 1 (DMT1) (iron responsive element (IRE) and non-IRE forms), duodenal cytochrome b (Dcytb), hephaestin (Hp), and iron regulated mRNA (Ireg1) mRNA. Animals are as described in fig 1. Expression of each gene was measured by ribonuclease protection assay. (A) Representative assays for each gene. (B) Band intensities for each gene were measured by densitometry and graphed as a proportion of glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Data are mean (SEM), n = 4. Significant difference from non-pregnant rats: *p<0.05, **p<0.01.
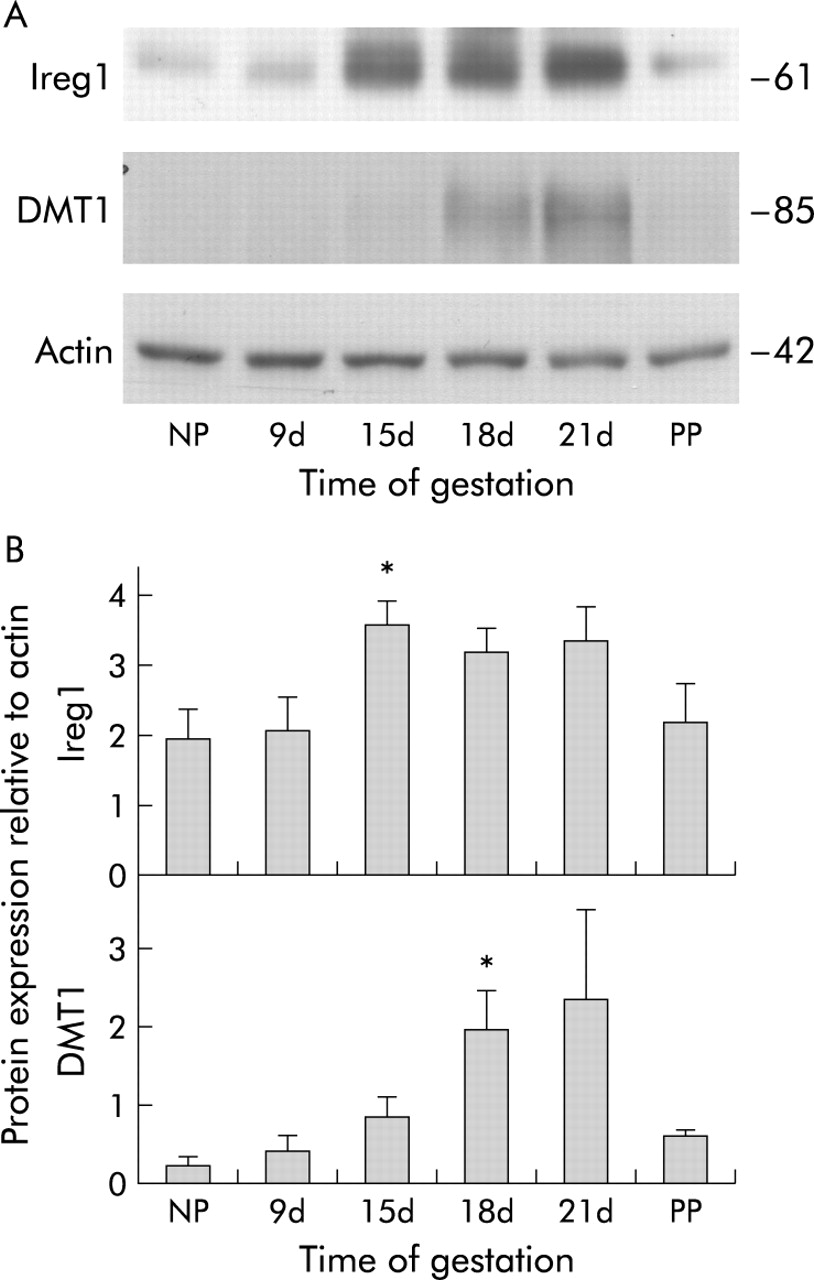
Expression of divalent metal transporter 1 (DMT1) and iron regulated mRNA (Ireg1) protein in the duodenum. Animals are as described in fig 1. The level of each protein was determined by western blotting. (A) Representative western blots for each protein. (B) Band intensities for each protein were measured by densitometry and graphed as a proportion of actin. Data are mean (SEM), n = 4. Significant difference from non-pregnant rats: *p<0.05.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Localisation of duodenal cytochrome b (Dcytb) and divalent metal transporter 1 (DMT1) by immunohistochemistry. Duodenal sections shown are from non-pregnant (NP) rats, pregnant rats at gestation days 18 and 21, and rats 24–48 hours post-partum (PP). Immunofluorescent staining was carried out using antibodies specific for Dcytb or DMT1. Sections were studied from four animals at each time point and representative sections are shown. Original magnification 100×.
Ireg1 mRNA expression showed a small increase in the latter stages of pregnancy but, unlike DMT1 and Dcytb, the change was not significant (fig 3). However, the Ireg1 protein level was increased on day 15 to 1.8 times non-pregnant controls (p<0.05) and this trend continued with 1.6-fold and 1.7-fold increases over control at days 18 and 21, respectively (p = 0.06). Protein expression returned to control levels after birth (fig 4). Hp mRNA expression was unchanged for the duration of pregnancy and after birth (fig 3).
DISCUSSION
The growing fetus has a high demand for iron which must be met from maternal sources. As the amount of iron transferred to the fetus throughout the course of pregnancy exceeds the amount of stored maternal iron under normal circumstances, there must be a compensatory increase in maternal iron absorption to provide the extra iron needed. In this study, we have undertaken a preliminary characterisation of this process at the molecular level.
As evidence of increased maternal-fetal transfer and mobilisation of maternal iron, we observed a drop in transferrin saturation and hepatic non-haem iron, and an increase in TfR1 expression as pregnancy progressed. This reduction in transferrin saturation and maternal iron stores was accompanied by an increase in the expression of the brush border iron transporter DMT1 and the iron reductase Dcytb in the duodenum, with maximum levels being observed in the last few days of gestation. One other group has examined DMT1 mRNA expression during pregnancy and they also found highest levels in the latter stages of gestation.19 Like DMT1 and Dcytb, expression of the basolateral iron exporter Ireg1 was upregulated in late pregnancy but the increase was only significant at the protein level. Expression of all three molecules fell sharply after birth. There was also an increase in transferrin saturation at this time, thus strengthening the relationship between increased systemic iron demand and maternal transporter expression. There were no significant changes in expression of the iron oxidase Hp.
The progressive increase in expression of intestinal iron transport molecules as pregnancy advances and the sharp fall in expression after birth are consistent with previous studies showing that iron absorption reaches a maximum in the latter stages of gestation1,6,7,20 and rapidly declines after birth.1,6 These studies have used a variety of techniques to demonstrate absorption changes, including 59Fe absorption through duodenal loops (rats), uptake of 59Fe from the diet (rats), and stable isotope studies (humans). The observed regulation is also consistent with numerous previous investigations showing that increased iron demands and depleted iron stores are associated with increased expression of the intestinal iron transport machinery.11,21 Other studies have highlighted the fact that while many iron transport molecules are regulated transcriptionally, they can also be regulated translationally and at the cellular level by iron dependent changes in subcellular localisation.18,22 For example, the latter mode of regulation could be important for Ireg1 and Hp, as recent data have indicated that the basolateral transfer step of iron absorption is likely to play a key role in how the intestine responds to changes in systemic iron demand.23,24 Indeed, in this study we found little regulation of Ireg1 at the mRNA level but a significant increase when protein was studied, suggesting that post-transcriptional mechanisms are most important in the regulation of this molecule during pregnancy.
In order to investigate how the body signals the small intestine to absorb more iron during pregnancy, we investigated expression of the liver derived regulatory peptide hepcidin. Hepcidin expression decreased progressively throughout pregnancy, reaching its lowest level just before parturition, then returned to near normal levels soon after birth. The inverse relationship between expression of hepcidin in the liver and iron transport proteins in the duodenum observed in this study is consistent with a growing body of evidence suggesting that hepcidin acts as a repressor of intestinal iron absorption. Mice and humans with abrogated hepcidin expression develop iron overload14,25 whereas mice overexpressing the gene develop iron deficiency anaemia.26 Similarly, experimentally induced iron overload results in increased hepcidin expression and reduced gut transporter expression while iron deficiency has the opposite effect.11,27 Our data suggest that hepcidin is a key messenger controlling intestinal iron absorption during pregnancy.
Factors leading to alterations in expression of hepcidin are poorly defined but recent evidence suggests that HFE and TfR2 are potential regulatory molecules. Mutations in either of these genes lead to iron overload subsequent to increased iron absorption.12,13 Furthermore, hepcidin expression is greatly diminished in HFE associated haemochromatosis, despite elevated iron stores,28 and the same response would be predicted in TfR2 associated iron overload as patients with HFE, hepcidin, and TfR2 mutations all have very similar phenotypes. Thus HFE (and possibly TfR2) appear to be upstream regulators of hepcidin expression and are required for hepcidin to respond appropriately to changes in body iron levels. Consequently, it was of some interest that we observed a decrease in expression of HFE, and to a lesser extent TfR2, mRNA levels during pregnancy. We recently proposed a mechanism by which HFE and TfR2 could respond to variations in transferrin saturation and subsequently influence hepcidin expression.15 As maternal iron levels and transferrin saturation are reduced during pregnancy, this is a potential mechanism for influencing hepcidin levels. However, other factors altering HFE and/or TfR2 expression could have a similar effect. In this regard it is notable that Batey and Gallagher1 found that iron absorption was still elevated in pregnant rats maintained on an iron loaded diet, albeit not to the same levels as found in animals on a control diet. This suggests that both iron dependent and iron independent factors are involved in determining the absorption response during pregnancy. It is possible that a placenta derived factor could play an important role in this regulation as the placenta is critical for iron delivery to the fetus.
How HFE and TfR2 themselves are regulated is unclear. Previous studies have suggested that expression of HFE is increased in intestinal cells lines with iron loading29 and decreased in the intestine in iron deficiency.30 However, the range of regulation was much smaller than the range of expression we observed in the liver during pregnancy. It is possible that this is due to tissue specific regulation, and that HFE in the liver is far more responsive to changes in tissue iron content than is HFE in the gut. Alternatively, there may be a pregnancy specific factor involved. In support of this possibility, earlier studies have failed to find evidence of TfR2 regulation in the liver when body iron load is varied,31 while we saw a significant decrease in TfR2 expression in late gestation. A candidate pregnancy specific factor is oestrogen. During pregnancy, oestrogen is initially secreted from the corpus luteum and later the placenta.32 Oestrogen levels progressively increase during pregnancy, peaking just prior to parturition, the same time that iron absorption is maximal, then dropping dramatically soon after birth (when the placenta is expelled), again similar to absorption.32,33 In addition, when oestrogen was administered to ovariectomised rats at concentrations similar to those found in late pregnancy, iron absorption was increased.34 Whether oestrogen plays a role in the regulation of HFE and/or TfR2 remains to be determined.
In conclusion, the changes in iron absorption that occur during pregnancy are consistent with the increase in duodenal iron transporter expression and the decrease in hepcidin expression observed in this study. Decreased expression of HFE and TfR2 is also likely to be important during gestation and may underlie the hepcidin response. Whether these molecules respond to maternal iron levels and transferrin saturation, a pregnancy specific factor, or a combination of both, will require further investigation.
Acknowledgments
This work was supported in part by grants from the National Health and Medical Research Council of Australia and the Human Frontier Science Program (grant RGY0328/2001-M).