Article Text

Abstract
Background and aims: Self-renewal and differentiation of intestinal epithelium is a tightly regulated process, whose perturbations are implicated in human colorectal tumourigenesis. The insulin/insulin-like growth factor (IGF) signalling pathway may play an important role in intestinal epithelium homeostasis. Insulin receptor substrate 2 (IRS2) is a poorly characterised component in this pathway.
Methods: Using complementary in vitro and in vivo human and murine models, expression (mRNA and protein levels), localisation (immunohistochemistry) and regulation of IRS2 were investigated in the normal intestine and colorectal tumours. In silico analysis of the human IRS2 promoter was performed together with reporter and chromatin immunoprecipitation assays.
Results: Significant IRS2 expression was detected in the intestine, with specific protein localisation in the villus region of the ileum and in the surface epithelium of the colon. In human HT29 and Caco2 cells, IRS2 mRNA levels increased with spontaneous and induced differentiation, together with CDX2 (caudal-related homeobox protein 2), P21 and KLF4 (Krüppel-like factor 4). Adenoviral infection with human CDX2 induced IRS2 expression in APC- (adenomatous polyposis coli) and β-catenin-mutated cells. On the other hand, IRS2 downregulation was observed in differentiated enterocytes after adenoviral infection with short hairpin CDX2 (shCDX2), in the intestine of CDX2 heterozygous mice and in colorectal tumours of ApcMin/+ and patients with familial adenomatous polyposis (FAP). The human IRS2 promoter region presents several CDX2-binding sites where CDX2 immunoprecipitated in vivo. IRS2 reporters were functionally activated via CDX2 and blocked via a dominant-negative CDX2 protein.
Conclusions: Combining gain- and loss-of-function approaches, an intriguing scenario is presented whereby IRS2 is significantly expressed in the apical intestinal compartment and is directly controlled by CDX2 in normal intestine and tumours.
Statistics from Altmetric.com
The intestinal epithelium consists of a single layer of epithelial cells mainly organised into submucosal invaginations termed crypts of Lieberkühn. At the crypt bottom reside stem cells that self-renew throughout life to generate progenitors that occupy the lower third of the crypt.1 2 In the midcrypt region, cells stop proliferating to differentiate into enterocytes. After reaching the epithelial surface, differentiated cells undergo apoptosis and are shed into the lumen. A tight regulation of the intestinal epithelium homeostasis is required to avoid neoplastic transformation. A body of evidence implies that Wnt signalling is a dominant force in controlling the process of intestinal mucosa renewal.2 One of the key Wnt signalling components is the scaffold protein encoded by the adenomatous polyposis coli (APC) gene. APC is part of a multiprotein destruction complex.3 In the absence of Wnt activators, free cytoplasmatic β-catenin is sequestered by APC into the destruction complex, phosphorylated and targeted for ubiquitin–proteosome degradation. In contrast, in the presence of Wnt cascade activators, or when APC is mutated, β-catenin accumulates in the cytoplasm, shuttles to the nucleus and, in association with the transcription factor T cell factor 4 (TCF-4), induces the expression of genes required for cell proliferation.4 5 APC was identified as a germline mutated gene in familial adenomatous polyposis (FAP),6 an autosomal dominantly inherited disease characterised by the development of colorectal adenomas within the fourth decade of life, with a very high risk of colorectal cancer (CRC).7 Later, somatic APC inactivation was found to occur in the majority of sporadic CRCs.8
Caudal-related homeobox protein 2 (CDX2) is considered to be a downstream mediator of APC tumour suppressor activity in intestinal cells.9 10 The human CDX2 gene is one of the three mammalian homologues of the Drosophila homeobox-containing gene Caudal, whose encoded transcription factor has a primary role during embryonic development, as assessed by the early lethality of Cdx2-deficient mouse embryos.11 From the midge stage onwards, the CDX2 transcription factor becomes selectively expressed in the crypt–villus epithelium of the small and large intestines.12 During embryogenesis, the CDX2 gene determines intestinal identity in the presumptive gut endoderm13 and it later controls the transcription of genes linked to the differentiation of the intestinal epithelium, such as Krüppel-like factor 4 (KLF4),10 sucrase-isomaltase,14 and P21.15 The expression of CDX2 becomes altered in human CRC in relation to the tumour grade.16 17 18 The relevance of these alterations has been uncovered in mice since reduction of Cdx2 expression facilitates tumour progression in models of genetically induced CRC.19 20 Restoration of CDX2 in colon cancer cells has also been shown to reduce cell migration in vitro and dissemination in vivo.21 On the other hand, the function of CDX2 might not be straightforward as an intestinal tumour suppressor since recent reports suggest a context- and cell type-dependent role.22 23 24
Insulin receptor substrate (IRS) proteins mediate the pleiotropic effects of insulin/insulin-like growth factor (IGF) signalling on mitogenic and metabolic processes. IRS proteins act as docking proteins between insulin, IGF-1 and cytokine receptors, and a complex network of intracellular molecules containing Src homology 2 (SH2) domains that are capable of binding IRS proteins phosphorylated at tyrosine residues.25 This results in the activation of several downstream pathways that are variably responsible for cell survival, proliferation, differentiation and metabolic responses. In particular, IRS2-activated pathways may have distinct roles in different organs or tissues, such as in female reproductive function,26 brain growth and metabolic homeostasis.27 Among these functions, IRS2 seems to have a primary involvement in metabolism. Indeed, IRS2-knockout mice have defects in hepatic insulin signalling and show altered growth in only a few tissues such as certain neurons28 and pancreatic β-cells.29 It is presently unknown if IRS2 is expressed in the intestine and how it is regulated. In contrast, it is known that IRS1 is expressed mostly in the intestinal crypts and that its disruption inhibits growth of intestinal epithelial cells,30 31 and confers protection against tumour formation in the ApcMin/+ mouse background.30 Interestingly, genetic IRS1 and IRS2 polymorphisms have been independently associated with CRC risk.32 Recently, a somatic mutation in the IRS2 gene has been identified in human CRC specimens.33
In the present study, we first show that IRS2 is significantly expressed in the villus–epithelial compartment of the adult intestine and that IRS2 is downregulated in APC-mutated intestinal tumours. Then, combining a variety of gain- and loss-of-function approaches, we present a novel and intriguing scenario whereby intestinal IRS2 expression is directly controlled by CDX2 in the normal intestine and colorectal tumours.
Patients and methods
Patients with FAP
Unrelated patients with FAP were recruited for the study after approval by the Ethics Committee of the University “G D’Annunzio” of Chieti. Written informed consent was obtained from each patient before mutation analysis and tissue harvesting. All the included cases presented with a classical FAP phenotype and harboured pathogenetic germline APC mutations.34 More information on patients and gene sequencing are available in the Supplementary methods online.
Mice and cell culture
Heterozygous Cdx2+/− mice11 and wild-type littermates were housed under pathogen-free conditions according to the guidelines of the Ethics Committee of the University Louis Pasteur of Strasbourg.20 ApcMin/+ mice were obtained from the Jackson Laboratory (Bar Harbor, Maine, USA). All mice were fed a standard rodent chow diet in a temperature-controlled room (23°C) on a 12 h light/dark cycle. All the experiments presented in this study have been carried out in male mice. The Ethics Committee of the Consorzio Mario Negri Sud approved this experimental set-up.
HT29 (HTB-38), CaCo2, HeLa and SW480 cells were obtained from ATCC (ATCC-LGC Promochem, London, UK). APC- or LacZ-inducible HT29 cells were a generous gift of Drs KW Kinzler and B Vogelstein,4 and TR4 or dominant-negative TCF-4- (DNTCF-4) inducible Ls174T cells were a generous gift of Dr H Clevers.35 Methodological details on cell culture are available in the Supplementary methods online.
RNA extraction and quantitative real-time PCR (qRT-PCR)
Following tissue and cell RNA isolation and purity check, cDNA was synthesised by retro-transcribing 4 μg of total RNA in a total volume of 100 μl using the High Capacity DNA Archive Kit (Applied Biosystems, Foster City, California, USA). qRT-PCR primers were designed using Primer Express software. qRT-PCR assays were performed as previously described using the ΔΔCT method for relative quantification.36 Methodological details and sequences of validated primers are reported in the Supplementary methods online.
Adenoviral constructs
Human CDX2 adenovirus (AdCDX2) was generated using the Viral Power Adenoviral Expression System (Invitrogen, La Jolla, California, USA). AdLacZ was used as control. Short hairpin CDX2 adenovirus (shCDX2) was generated based on a previously validated sequence against human CDX237 using the BLOCK-iT U6 RNA entry vector kit (Invitrogen). shLacZ was used as a control. Details on the cloning of plasmids, and on purification and propagation of the viruses are reported in the Supplementary methods online. The adenoviral titre was determined by qRT-PCR with specific primers.38
Luciferase reporter constructs
Regions between −2394/−1740, −2269/−1740, −2089/−1740, −2039/−1740 and −1860/−1740 of the pGL3-IRS2 (−2399/+217) plasmid (a kind gift of Dr M Kasuga, Kobe University, Japan) containing the 5′-flanking region of the human IRS2 promoter39 were PCR amplified and subcloned into the polylinker of the promoter-less pGL3-basic vector (Promega, Madison, Wisconsin, USA) using MluI 5′ and HindIII 3′ restriction sites to generate pGL3-IRS2 (−2394/−1740), pGL3-IRS2 (−2269/−1740), pGL3-IRS2 (−2089/−1740), pGL3-IRS2 (−2039/−1740) and pGL3-IRS2 (−1860/−1740) reporter constructs.
Statistical analysis
All results are expressed as the mean (SEM). Data distribution and gene expression statistical analysis was performed using NCSS statistical and power analysis software 2007 (Kaysville, Utah, USA). Multiple groups were tested by one-way analysis of variance (ANOVA) followed by the Fisher least significant difference test for unpaired data. Comparisons of two groups were performed using the Student t test followed by a Mann–Whitney U test where appropriate. A p value <0.05 was considered significant.
Methods used for western blot, immunohistochemistry and immunocytochemistry, alkaline phosphatase measurements, transient transfection and luciferase activity, chromatin immunoprecipitation (ChIP) and gel-shift electromobility shift assays (EMSAs) are available as Supplementary methods online.
Results
IRS2 expression and localisation in the intestine
The first step of our study consisted of the assessment of IRS2 gene expression in the murine gastrointestinal tract and the exact localisation of IRS2 protein in the crypt–villus axis of the intestinal mucosa. Significant IRS2 mRNA levels were found in the entire enterohepatic system (liver, stomach, duodenum, jejunum, ileum and colon), as compared with white and brown adipose tissue that showed the highest IRS2 expression levels (fig 1A). Immunohistochemical analysis of the terminal ileum and proximal colon revealed clear IRS2 cytoplasmic localisation within the differentiated absorptive enterocytes of the villus compartment (ileum) and surface epithelium (colon), together with L-FABP (L-type fatty acid-binding protein) (fig 1B). No IRS2 labelling was observed in the Ki67-stained proliferating progenitor cells of the crypt compartment (fig 1B).

Insulin receptor substrate 2 (IRS2) expression levels and localisation in mice tissues. (A) IRS2 mRNA expression was measured by quantitative real-time PCR using cyclophilin as a reference gene. Results are expressed as the mean (SEM). The mean Ct of ileum samples is reported in number. (B) Paraffin-embedded terminal ileum (upper panel) and proximal colon (lower panel) specimens were immunostained with IRS2 antibody to determine the expression and localisation of IRS2 protein. Immunostaining of Ki67 and L-type fatty acid-binding protein (L-FABP) was used as a marker of the proliferative crypt and differentiated villus compartments, respectively (×400 magnification). WAT, white adipose tissue; BAT, brown adipose tissue.
Changes in IRS2 expression during intestinal cell differentiation
Since IRS2 is solely expressed in the differentiated apical compartment of the intestinal epithelium, we analysed the behaviour of IRS2 expression in a human CRC cell line during spontaneous and induced in vitro differentiation. As a first step, IRS2 mRNA and protein expression was evaluated in HT29 cells in preconfluent, confluent and postconfluent states.40 Under these conditions, a clear induction of IRS2 protein expression was observed together with the upregulation of CDX2 protein (fig 2A).40 Alkaline phosphatase (ALP) activity was used as a surrogate marker of enterocyte differentiation (fig 2B,C). A net increase in IRS2 mRNA levels was observed during confluence-induced growth arrest and enterocyte differentiation (fig 2B), which was accompanied by increased expression of genes that are known to be associated with the degree of intestinal differentiation, such as CDX2, KLF4 and P21. Since in vitro postconfluence growth also increases cellular metabolic requirements, in order to know if the induction of CDX2 and IRS2 is related to differentiation or increased metabolic requirements, we studied SW480 colon cancer cells that are not able to differentiate after postconfluence.41 Interestingly, there was no increase in ALP activity in postconfluent SW480 cells with no induction of CDX2, IRS2 and P21 mRNA levels (Supplementary fig S1). Thus, in the postconfluent state, SW480 cells increase their metabolic requirements without overexpressing CDX2 and IRS2.

Insulin receptor substrate 2 (IRS2) expression levels during confluence-induced growth arrest and sodium butyrate (NaB)-induced differentiation in HT29 cells and after confluence in Caco2 cells. (A) Expression levels of IRS2 and CDX2 (caudal-related homeobox protein 2) in HT29 cells (preconfluence, confluence and postconfluence; vehicle and NaB treated) were observed by western blot. Alkaline phosphatase (ALP) activity was analysed as a marker of differentiation in HT29 cells (preconfluence, confluence and postconfluence (B); vehicle and NaB treated (C)). Quantitative real-time PCR (qRT-PCR) was performed to measure IRS2, CDX2, KLF4 (Krüppel-like factor) and P21 mRNA levels in HT29 cells (preconfluence, confluence and postconfluence (B); vehicle and NaB treated (C)). (D) qRT-PCR was performed to measure IRS2, CDX2, KLF4 and P21 mRNA levels in Caco2 cells at different time points after confluence. Values shown represent the mean (SEM) for four independent determinations performed in duplicate. Cyclophilin was used as a reference gene and values were normalised to data obtained from preconfluent or vehicle-treated cells. Different lower case letters indicate significant statistical difference (b different from a, p <0.05; c different from a and b, p <0.05, analysis of variance).
Treatment with sodium butyrate (NaB) at 1 mM concentration promotes differentiation of intestinal cell lines. Also, CDX2 expression is stimulated in HT29 cells by NaB treatment at a dose that promotes cell differentiation.42 Here we show that IRS2 mRNA (fig 2C) and protein (fig 2A) levels significantly increased in HT29 cells after 24 h treatment with 1 mM NaB. Upregulation of IRS2 was accompanied by increased CDX2, KLF4 and P21 mRNA levels and ALP activity (fig 2C). As a further confirmation, an increase in CDX2, IRS2, KLF4 and P21 mRNA levels was also observed in Caco2 cells during differentiation (fig 2D).
The CDX2–IRS2 axis regulates IGF responses during intestinal cell differentiation
Since IRS2 is presumably part of the IGF signalling pathways in the intestine, we decided to study IGF-1-induced activation of extracellular signal-regulated kinase (Erk) in preconfluent (low CDX2–IRS2 expressing) and postconfluent (high CDX2–IRS2 expressing) HT29 colon cancer cells. We therefore treated preconfluent and postconfluent HT29 cells with 100 nmol/l IGF-1 for 0–30 min and we measured total and phosphorylated forms of Erk1 and Erk2.43 Representative immunoblots are reported in fig 3A. Also, we performed a fluorescence-activated cell sorting (FACS) analysis of the same cells with and without treatment with 100 nmol/l IGF-1. Interestingly, when the levels of endogenous IRS2–CDX2 were low (ie, preconfluent HT29 cells), IGF-1 was able to phosphorylate both Erk1 and Erk2 to a certain extent and to induce a strong increase in cell proliferation, as shown by the significant amount of cells in S-phase (from 13.49% to 57.52%, vehicle vs IGF-1, p <0.01, fig 3B). On the other hand, in postconfluent HT29 cells that express high levels of CDX2 and IRS2, there is a basal increase in total and phosphorylated Erk1 and 2 as compared with preconfluent HT29 cells. Furthermore, under these conditions, IGF-1 was unable to induce any changes in total and phosphorylated Erk or in cell proliferation, as shown by no changes in the amounts of cells in phases G0G1, G2M and S (fig 3B). Taking these data together, we conclude that when CDX2 induces IRS2 expression in postconfluent HT29 colon cancer cells, IGF-1 fails to stimulate phosphorylation of Erk1 and 2 as well as to induce cell proliferation. These findings are consistent with the idea that in the intestine IRS2 is expressed in differentiated low proliferating enterocytes, where it may counteract IGF-1-induced proliferation.

Activation of extracellular signal-regulated kinase (Erk) by insulin-like growth factor-1 (IGF-1) in HT29 cells after confluence. (A) Stimulation of Erk in preconfluent and postconfluent HT29 cells was studied in the absence or presence of 100 nmol/l IGF-1 for the indicated times. Erk1/2 phosphorylation and protein were evaluated by immunoblotting with antibodies specific to total or phoshorylated Erk1/2. (B) Fluorescence-activated cell sorting (FACS) analysis in preconfluent and postconfluent HT29 cells with or without the 100 nmol/l IGF-1. Preconfluent cells treated with IGF-1 showed an accumulation in S-phase; conversely, postconfluent cells showed an accumulation in G0G1.
Downregulation of IRS2 expression in colorectal tumours from patients with FAP and ApcMin/+ mice
Enterocyte differentiation from proliferative crypt precursors is a homeostatic process where Wnt signalling and APC status represent the dominant factors in controlling cell fate. FAP is a classic model of CRC, where the earliest phase is exclusively initiated by and dependent on APC gene mutations. FAP adenomas grow within the top crypt–villus compartment retaining a “crypt progenitor-like” phenotype generated by the loss of APC and the subsequent accumulation of the β-catenin protein at high levels.2 We therefore studied IRS2 and CDX2 expression levels in colorectal adenomas obtained by endoscopic resection from patients with FAP with molecularly defined germline and somatic APC mutations.6 IRS2 and CDX2 mRNA levels were reduced in adenomas as compared with matched normal mucosa (fig 4A). Also, IRS2 protein was decreased in the aberrant crypt foci and colon tumours of ApcMin/+ mice as revealed by immunohystochemical analysis, where Ki67 was used as control for the proliferative compartments (fig 4B).
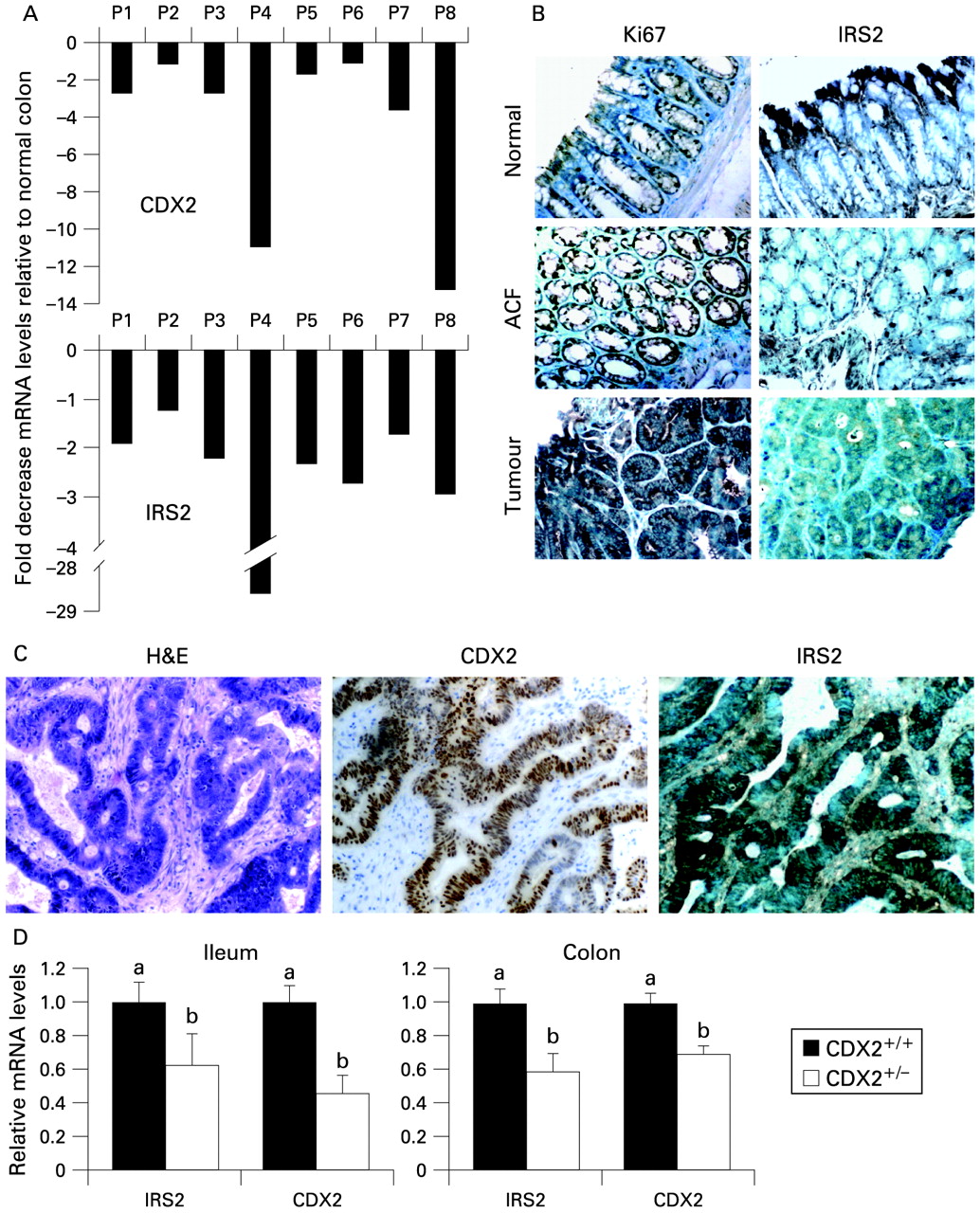
(A) Insulin receptor substrate 2 (IRS2) and caudal-related homeobox protein 2 (CDX2) expression in colon adenomas from patients with familial adenomatous polyposis (FAP). Quantitative real-time PCR (qRT-PCR) was performed on total RNA from colorectal adenomas and adjacent normal mucosa samples obtained from patients with FAP. Values shown represent the mean (SEM) of four independent determinations performed in duplicate. Cyclophilin was used as a reference gene and values were normalised to data obtained in the normal mucosa (plotted as 0 on the x-axis). (B) Paraffin-embedded specimens from normal colonic mucosa, aberrant crypt foci (ACF) and colon tumours of ApcMin/+ mice. Sections were immunostained with the IRS2 antibody to determine expression and localisation of IRS2 protein in normal tissue and tumours. Immunostaining of Ki67 protein was used as a marker of the proliferative cell compartment (×400 magnification). (C) Immunohistochemical analysis of sections of human colorectal tumours with certain CDX2 expression. Representative fields of sections of human colon tumours stained for CDX2 and IRS2 (×160 magnification). (D) IRS2 expression was measured in the ileum and colon of CDX2+/− mice tissues and corresponding wild-type mice. qRT-PCR was performed to measure IRS2 and CDX2 mRNA levels. Values shown represent the mean (SEM) for two independent determinations performed in duplicate. Cyclophilin was used as a reference gene and values were normalised to data obtained from wild-type mice. Different lower case letters indicate significant statistical difference (b different from a, p <0.05, t test).
CDX2 is expressed in a heterogeneous pattern in some CRCs in relation to their degree of differentiation,18 22 and sometimes amplified.23 Thus, we analysed the expression of IRS2 in human colon cancer samples that presented CDX2 expression heterogeneously. Intriguingly, there was also a high correspondence between CDX2 and IRS2 levels in human colon tumours with certain CDX2 expression (fig 4C), thus supporting our hypothesis of IRS2 being a target of CDX2.
Downregulation of IRS2 mRNA levels in CDX2+/− mice
To confirm further that IRS2 is a direct target of CDX2, we analysed the IRS2 and CDX2 mRNA levels in the intestine of CDX2+/− mice. While homozygous CDX2−/− mice are embryonic lethal, CDX2 heterozygous mice have a compromised CDX2 function, develop gastric heteroplasia in the intestine, and are hypersensitive to CRC upon treatment with a DNA mutagen20 and under genetic predisposition in the ApcMin/+ background.19 Here we show that IRS2 expression levels are lower in the terminal ileum and proximal colon of CDX2+/− mice as compared with corresponding wild-type mice (fig 4D). Interestingly, IRS2 levels were positively correlated with CDX2 levels in all the examined samples.
CDX2 and IRS2 mRNA levels are upregulated in HT29 cells with APC-inducible expression and unchanged in Ls174T cells with DNTCF4-inducible expression
Since IRS2 expression is significantly decreased in mouse and human tumours carrying an APC mutation, we decided to investigate if re-expression of a wild-type APC in an APC-mutated cell line would restore IRS2 expression. Like most CRC cell lines,44 HT29 cells have no functional APC protein, while two C-terminally truncated isoforms of approximately 100 and 200 kDa, expressed by mutated APC alleles, are present. Inducible expression of full-length APC in HT29 cells results in a significant decrease of cell proliferation via induction of cell death through apoptotic pathways.45 In line with previous reports,9 10 when APC expression is re-established in HT29-APC cells via ZnCl2, CDX2 (fig 5A) and KLF4 and P21 (data not shown), mRNA levels are significantly increased. Under these conditions, there is a net upregulation of IRS2 expression levels (fig 5A), accompanied by an increase in P21 mRNA levels (data not shown). HT29-β-Gal cells treated with ZnCl2 exhibited no significant difference in CDX2 and IRS2 mRNA levels (fig 5A). A net increase in IRS2 protein levels in HT29APC cells after ZnCl2 was observed via immunocytochemical analysis (fig 5B). In contrast, in Ls174T cells that present a mutated β-catenin with transcriptionally activated β-catenin–TCF-4 complexes, the induction of DNTCF4 decreased β-catenin–TCF-4 target genes and increased P21 mRNA levels (data not shown),35 but did not alter the mRNA levels of CDX2 and IRS2 (fig 5B). These data underscore the necessity for a functional cytoplasmic APC programme for the expression of IRS2.
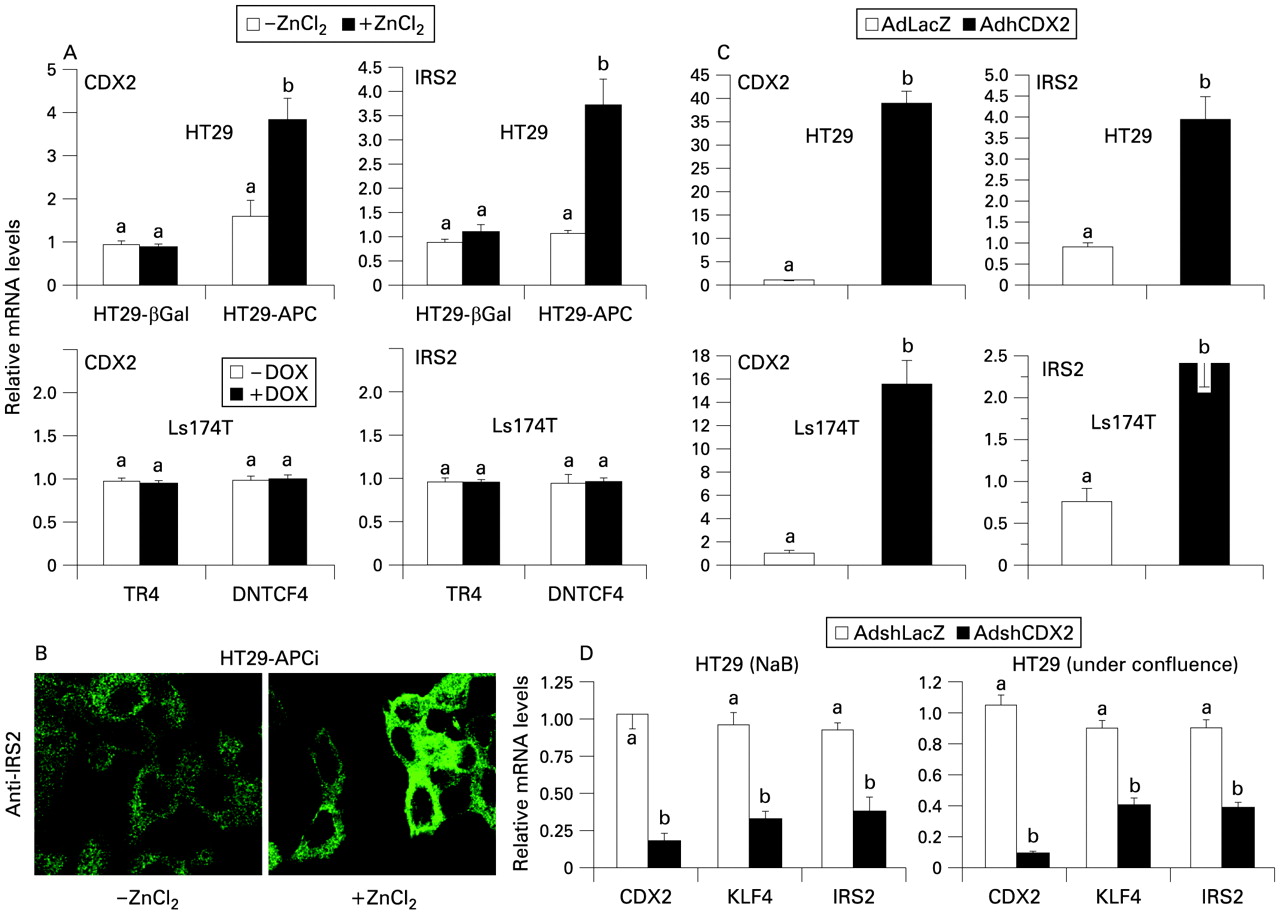
(A) HT29 adenomatous polyposis coli (APC)-inducible and LacZ-inducible cells (upper panel) were treated with 120 μM ZnCl2 or vehicle for 24 h. Ls174T TR4 and dominant-negative T cell factor 4 (DNTCF-4)-inducible cells (lower panel) were treated with 1 μg/ml doxycyclin (DOX) or vehicle for 24 h. Quantitative real-time PCR (qRT-PCR) was performed to measure IRS2 and CDX2 mRNA levels. Values shown represent the mean (SEM) for four independent determinations performed in duplicate. Cyclophilin was used as a reference gene and values were normalised to data obtained in vehicle-treated cells. (B) Immunostaining was performed to visualise IRS2 protein in HT29-APC-inducible cells treated with ZnCl2 or vehicle (×1260 magnification). (C) Expression of insulin receptor substrate 2 (IRS2) in HT29 (upper panel) and Ls174T (lower panel) cells after adenoviral caudal-related homeobox protein 2 (CDX2) infection. qRT-PCR was performed to measure IRS2 and CDX2 mRNA levels 48 h after infection with 100 multiplicity of infection (MOI) of CDX2 or control LacZ adenoviruses. Values shown represent the mean (SEM) for four independent determinations performed in duplicate. Cyclophilin was used as a reference gene and values were normalised to data obtained from control LacZ adenovirus-infected cells. (D) IRS2, Krüppel-like factor 4 (KLF4) and CDX2 expression levels were measured in HT29 cells treated with sodium butyrate (NaB) and under confluence after infection with short hairpin (sh) CDX2 or shLacZ control adenoviruses. qRT-PCR was performed to measure IRS2 and CDX2 mRNA levels. Values shown represent the mean (SEM) of four independent determinations performed in duplicate. Cyclophilin was used as a reference gene and values were normalised to data obtained from adenovirus shLacZ-treated cells. Different lower case letters indicate significant statistical difference (b different from a, p <0.05, t test).
Increased IRS2 expression in HT29 and Ls174T cells via adenoviral CDX2 infection
To prove if CDX2 was also able to induce IRS2 expression in the absence of a functioning APC programme, we evaluated expression of IRS2 in APC-mutated HT29 cells after infection with CDX2 or control LacZ adenoviruses. Induction of CDX2 expression in HT29 cells was correlated with a significant upregulation of IRS2 (fig 5C), and KLF4 and P21 mRNA levels (data not shown). Also, the forced CDX2 expression via AdCDX2 in preconfluent HT29 cells is linked to increased IRS2 protein levels (Supplementary fig S2). Interestingly, the same results are obtained in β-catenin-mutated Ls174T cells under the same conditions (fig 5C), thus underlining the independence of CDX2 transcriptional activity from the cytoplamic APC and nuclear β-catenin–TCF-4 status of the cell.
CDX2 is needed for IRS2 expression during intestinal cell differentiation
To verify if the induction of IRS2 expression during enterocyte differentiation depends on CDX2 levels, we infected HT29 cells with shCDX2 or shLacZ adenoviruses. The shCDX2 adenovirus that we generated was able to induce an 85% decrease in endogenous CDX2 mRNA levels (fig 5D). Significant downregulation of both KLF4 and IRS2 mRNA levels was observed after NaB- or confluence-induced differentiation (fig 5D), thus proving the necessity of CDX2 for IRS2 expression during intestinal cell differentiation.
Direct activation of the IRS2 reporter via CDX2 and binding of CDX2 to the IRS2 promoter
CDX2 is a DNA-binding transcription factor, which recognises TTTAT or ATAAA motifs in the promoter region of its target genes.46 We therefore performed an in silico analysis of the human IRS2 gene promoter that identified several putative CDX2 binding sites (fig 6A). We then generated a series of reporters of the IRS2 gene promoter including a progressively decreasing number of the putative binding sites. pGL3-basic was used as a control. Details of the reporter constructs are given in the Patients and methods section and in fig 6A. Potential CDX2 binding sites were proved to be functionally active in HeLa cells (fig 6B) after co-transfection with CDX2 plasmids. The full-length pGL3-IRS2 (−2399/+217)39 presents a 2.5-fold induction by CDX2, yet with a very high basal activity (data not shown). Importantly, a dominant-negative CDX2 mutant expression plasmid was able to block activation of IRS2 reporters by CDX2 (fig 6C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Transactivation of insulin receptor substrate 2 (IRS2) reporter assay by caudal-related homeobox protein 2 (CDX2). (A) Shown is a schematic representation of the IRS2 gene promoter. Putative CDX-binding elements (CBS1, CBS2, CBS3, CBS4) are depicted. Transcriptional activation of the different reporter constructs was measured in HeLa cells in the presence of wild-type full-length human CDX2 (B) or dominant-negative mutant CDX2 (C) expression plasmids or control (pCDNA4b). pGL3-basic was used as a negative control. Different lower case letters indicate significant statistical difference (b different from a, p <0.05; c different from b and a, p <0.05; d different from c, b and a, p <0.05, analysis of variance). (D) A chromatin immunoprecipitation assay was performed in preconfluent (low CDX2 expressing) and postconfluent (high CDX2 expressing) HT29 cells and in preconfluent HT29 cells after transfection of CDX2 expression plasmid or control (pCDNA4B). Experiments were performed with and without the CDX2 antibody. Quantitative real-time PCR (qRT-PCR) was used to measure the amount of DNA that was pulled down during the experiments. Different lower case letters indicate significant statistical difference. Experiments were performed with and without the CDX2 antibody. qRT-PCR was used to measure the amount of DNA that was pulled down during the experiments. Different lower case letters indicate significant statistical difference (b different from a, p <0.05, t test). (E) Gel-shift electrophoretic mobility shift assays were performed to show the physical interaction between CDX2 protein and IRS2 promoter DNA. Synthetic oligonucleotides representing CDX2 putative binding sites on the human IRS2 gene promoter are shown. Note in bold the CDX core-binding sequence (ATAAA). Nuclear extract (5 μg) from HeLa cells infected with adenovirus (Ad) CDX2 was incubated with the 32P-labelled IRS2 oligonucleotide probe alone (lane 2) and in the presence of 50, 100 or 500 unlabelled IRS2 competitor oligonucleotides (lanes 3–5). In lane 1, only 32P-labelled IRS2 oligonucleotide was added. Samples were loaded on a 5% acrylamide gel.
To prove that CDX2 binds in vivo the IRS2 promoter motifs that are functionally active in the reporter assay, we performed a ChIP assay in postconfluent (endogenous CDX2 expressing) HT29 cells. As shown in fig 6D, significant precipitation of CDX2 was observed in the specific regions of the IRS2 gene promoter. Specific validated qRT-PCR primers were used together with a series of controlled settings (no antibody, aspecific antibody, primers amplifying the region of the IRS2 gene outside the promoter or far inside the promoter). We recognise that the promoter motifs are located within 200–400 bp, thus decreasing the specificity of the pull-down for the single motifs, for which DNA segment length should be at least 600–700 bp. Thus, we decided to present the data as the mean of the copies obtained by the different primer couples. We also performed a ChIP assay in preconfluent (virtually no CDX2 expressing) HT29 cells after infection with AdCDX2 and confirmed the data obtained on endogenous CDX2 with postconfluent cells. Finally, we performed a gel-shift EMSA using the four oligonucleotide probes and we proved that at least two of the four present a strong binding with CDX2 with an elegant competition with the same unlabelled probe (fig 6E). Taken together, the data obtained in the reporter assay, ChIP and gel-shift EMSAs support the compelling hypothesis that IRS2 is a direct target of CDX2.
Discussion
In the present study we show that IRS2 is significantly expressed in the intestine where it is localised in the differentiated absorptive enterocytes of the apical villus–epithelial compartment. IRS2 expression is highly regulated during the differentiation of intestinal epithelial cells via a direct mechanism involving the transcription factor CDX2.
The process of intestinal mucosa self-renewal and differentiation is tightly controlled by a series of events that involve several regulatory transcriptional pathways.2 The intestinal epithelium is organised in crypt–villus functional units; these units are organised in a dynamic scenario in which progenitor cells are generated from the stem cell population at the crypt bottom and migrate along the crypt axis until exfoliation as terminally differentiated cells at the villus tip. The mechanisms controlling cell transitions from one compartment to the next are complex and involve transcription factors that switch compartment-specific genes on and off. CDX2 has been shown to confer positional information along the crypt–villus axis, inducing the transcription of genes associated with intestinal epithelium differentiation. Multiple pieces of evidence indicate that the Wnt cascade and the APC programme act as dominant forces controlling cell migration and differentiation along the crypt–villus axis.2 Mutations in this cascade inevitably result in development of CRC. Expression of CDX2 has been found to decrease with tumour grade in human CRC16 and in mouse and rat models of intestinal tumourigenesis.11 16 Nevertheless, it has been reported that Cdx2 expression is maintained but heterogeneous in most human colon cancers,18 22 with levels of expression related to the degree of differentiation of the tumours.17 Knowledge of the factors that control cell fate in the intestine will contribute to the discovery of the nutritional and metabolic hits that drive the downstream elements of activation of the constitutive Wnt pathway during CRC progression.7
A growing body of evidence implies IGF-1, IGF-2, IGF-binding proteins (IGFBPs) and insulin in CRC progression.47 48 49 50 The IRS proteins are central mediators of insulin/IGF signalling and are implicated in basic cellular functions such as growth, survival and metabolism.25 IRS1 is considered a marker of active IGF-1 signalling pathways in tumours.50 IGF-1 receptor-induced activation of IRS1 regulates subcellular localisation and activity of β-catenin, activating β-catenin/TCF-4 target genes, such as cyclin D1.51 Reductions in crypt IRS1 levels increase apoptosis of crypt stem or progenitor cells, protect against β-catenin-driven intestinal tumours and reduce Sox9, a β-catenin/TCF-4 target gene. Indeed, disruption of IRS1 inhibits growth of colon cancer cells31 52 and leads to a protection against tumour formation in the background of ApcMin/+ mice.30 While IRS1 seems to be primarily involved in the antiapoptotic and trophic actions of insulin and IGF, IRS2 might have a preferential role in metabolism.47 Nothing is currently known about its expression and regulation in intestinal epithelia.
Using gain- and loss-of-function models, we provide evidence for a direct regulation of IRS2 expression via CDX2. Interestingly, induction of IRS2 expression by CDX2 appears to be independent from APC. On one hand, re-expression of a wild-type APC protein in APC-mutated HT29 cells45 is able to induce the expression of CDX2 and IRS2. On the other hand, there is no restoration of CDX2 and IRS2 expression after inhibition of β-catenin/TFC-4 transcriptional activity via DNTCF-4 in β-catenin-mutated Ls174T cells.35 These findings point to an APC-dependent mechanism as the basis of the regulation of CDX2 expression in intestinal cells. Nevertheless, expression of CDX2 via adenoviral infection is able to activate the transcription of IRS2 as well as KLF4 in both cellular models, thus proving the independence of CDX2 transcriptional activity from the APC or β-catenin/TCF-4 status of the cells.
Here we present a novel scenario for the regulation of IRS2 in the intestinal architecture. In the crypt–villus axis, the IRS2 protein is specifically localised in the terminally differentiated surface compartment. IRS2 expression is almost lost in colorectal adenomas initiated by APC mutation. Furthermore, IRS2 expression directly correlates with the degree of spontaneous or induced enterocytic differentiation in CRC cells and with growth arrest, IRS2 being a direct target of the intestine-specific transcription factor CDX2. One could speculate that as proliferative progenitors migrate along the crypt axis to differentiate into mature enterocytes, CDX2, by inducing IRS2 expression, could contribute to switching insulin/IGF-related signalling pathways towards differentiation and metabolism. Future studies aimed at disclosing cell type-specific activities of IRS proteins via the generation of intestinal-specific transgenic mice are needed to verify this intriguing possibility.
Acknowledgments
We are indebted to Drs J Auwerx, F Beck, H Clevers, M German, F Giorgino, M Kasuga, K W Kinzler, A V Lee, S Perrini, K Schoonjans and B Vogelstein for their valuable tools. We thank A Di Santo, V Evangelista, G Lo Sasso, N Martelli and S Manarini for their valuable help during the study.
REFERENCES
Supplementary materials
Web only appendix 58:9;1250
Files in this Data Supplement:
Footnotes
Funding A Moschetta is funded by a Start Up Grant 2005–1007 of the Italian Association for Cancer Research (AIRC, Milan, Italy), by the European Research Council Starting Independent Grant-IDEAS 2008, by the Italian Ministry of Health and Education (FIRB-RBID08C9N7), by the European Community's Seventh Framework Programme FP7/2007–2013 under Grant Agreement No 202272 (LipidomicNet) and by the University of Bari. A Morgano is a fellow of the G d’Annunzio University Oncology Program. MP is supported by the Rosario Samanin Fund. M-TV is supported by the Association pour la Recherche sur le Cancer, France.
Competing interests None.
See Commentary, p 1179
Ethics approval The Ethics Committees of Consorzio Mario Negri Sud (Santa Maria Imbaro, Chieti, Italy), University “G. D’Annunzio” (Chieti, Italy) and University Louis Pasteur (Strasbourg, France) approved the study protocols for ApcMin/+ mice, patients with FAP and CDX2+/− mice, respectively.
▸ Additional methods, figures and a table are published online only at http://gut.bmj.com/content/vol58/issue9