Article Text

Statistics from Altmetric.com
We read with interest the paper by Derikx et al 1 replicating the association of the minor T allele of single nucleotide polymorphism rs10273639C/T, which is located 408 bp upstream of the translation initiation codon of the cationic trypsinogen (PRSS1) gene, with a protective effect against chronic pancreatitis.2 However, whether rs10273639 is the causal variant or not remains unknown. Resolving this issue is of intrinsic biological interest, and it may also have diagnostic and therapeutic value.
During resequencing of the promoter region of PRSS1 in 287 French Caucasian individuals (see online supplementary material), we found that rs4726576C/A, which is located 204 bp upstream of the translation initiation codon of PRSS1 (figure 1A), is in perfect linkage disequilibrium (LD) with rs10273639C/T (figure 2A). To identify which polymorphism is of potential biological relevance, we performed a two-step luciferase promoter reporter assay (see online supplementary material). First, we sought to establish whether the proximal promoter of PRSS1 is sufficient to drive specific gene expression. We thus constructed two luciferase reporter plasmids in the context of the two major alleles, one encompassing the two polymorphic sites [pGL3-(C-C)] and the other encompassing only the rs4726576 site [pGL3-(C)] (figure 1A). We transfected the two luciferase constructs into rat pancreatic acinar AR42J cells treated with dexamethasone and human embryonic kidney HEK293T cells to assess their activities against the pGL3-basic background. An ∼20-fold increase in gene expression was observed for both constructs in AR42J, whereas no increased expression was observed in HEK293T (figure 1B). There was no significant difference in gene expression in AR42J between the two constructs (p=0.2751), suggesting functional sufficiency of the proximal promoter of PRSS1 in the regulation of basal gene expression. To obtain further supporting evidence for this point, we then investigated whether the pGL3-(C) construct can drive gene expression within intact parenchymal pancreatic tissue in mice. For this purpose, we developed an in vivo electroporation technique coupled to the luciferase assay, based on the intrapancreatic parenchymal injection for gene transfer delivery3 (see online supplementary material and figure S1). The proximal promoter of the PRSS1 gene was also functional in the transfected mouse pancreatic tissues (figure 1B).

Functional analysis of rs10273639 and rs4726576. (A) Upper panel illustrates the human PRSS1 promoter region in the context of the major alleles for rs10273639 and rs4726576. Nucleotide positions are numbered using the A of the translation initiation codon ATG as +1. Lower panel illustrates the pGL3-(C-C) and pGL3-(C) luciferase constructs for PRSS1 promoter activity assay. (B) Left panel, promoter activities of pGL3-(C-C) and pGL3-(C) constructs in relation to the pGL3-basic vector, in AR42J cells treated with 100 nM dexamethasone (Dx) and HEK293T cells. Results are means±SD of three independent experiments. Right panel, promoter activity of the pGL3-(C) construct in the mouse parenchymal pancreatic tissue. Results are means±SD of four injections. (C) Illustration of all possible luciferase constructs with respect to variations at positions c.1-408 and c.1-204. (D) Relative promoter activities of the luciferase constructs (as illustrated in C) in AR42J cells treated with 100 nM dexamethasone. Results are means±SD of at least three independent experiments. **p<0.01. TSS, transcription start site.
{kind=link}
{kind=link}
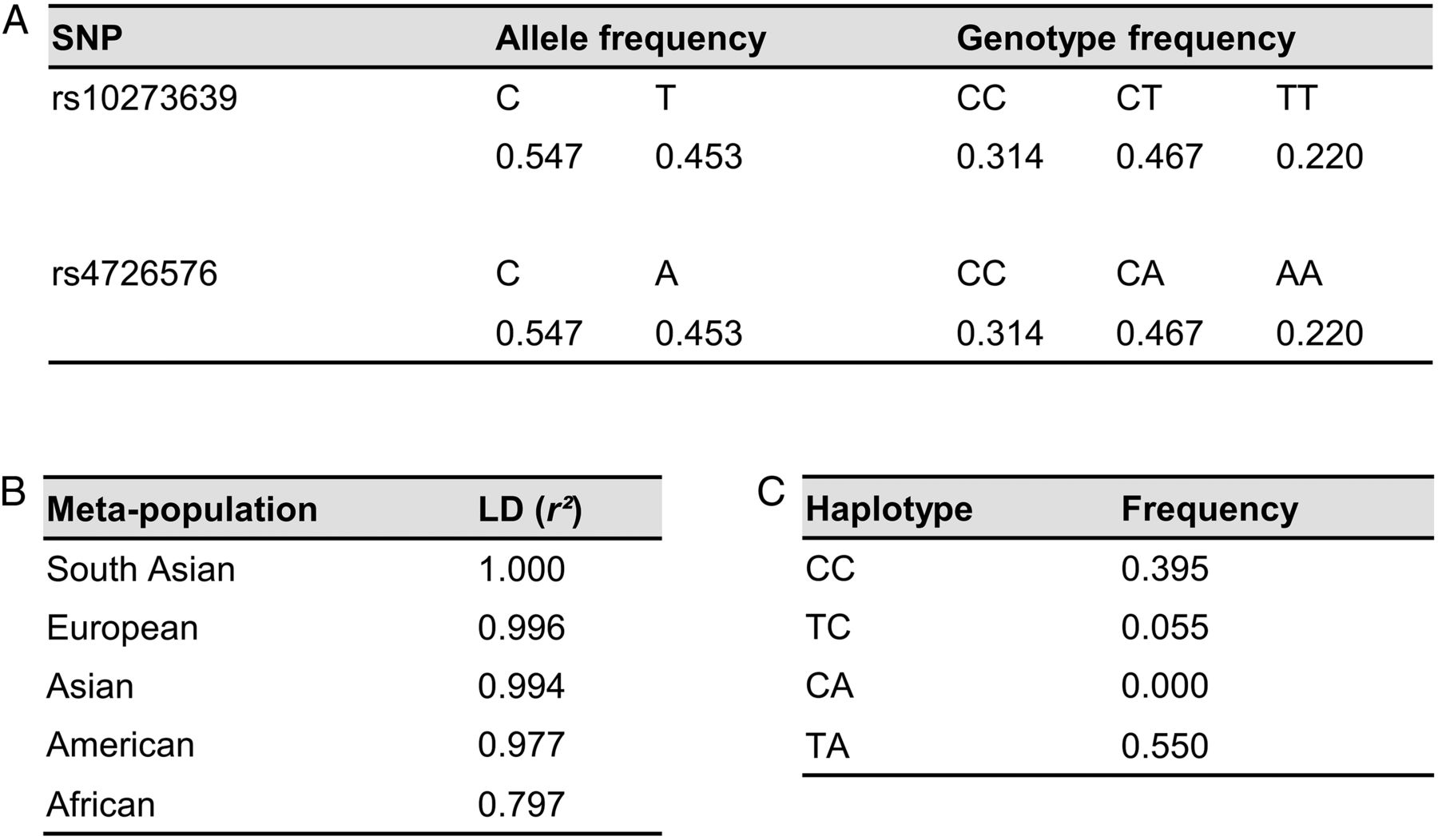
Linkage disequilibrium (LD) and distribution of rs10273639 and rs4726576 in different populations. (A) Allele and genotype frequencies of the two single nucleotide polymorphisms (SNPs) in 287 French individuals of European ancestry. rs10273639 and rs4726576 were calculated to be in perfect LD (r2=1). (B) LD data between the two SNPs in five meta-populations. (C) Haplotype frequencies for SNP pair [rs10273639 rs4726576] in the African meta-population.
In the second step, we constructed luciferase reporter plasmids corresponding to the other three possible haplotype configurations by reference to the pGL3-(C-C) plasmid as well as to the minor A allele of rs4726576 by reference to the pGL3-(C) plasmid (figure 1C). Subsequent reporter gene assays demonstrated that it is the minor A allele of rs4726576 that was associated with reduced gene expression (figure 1D). In other words, it is the rs4726576 A allele rather than the rs10273639T allele that underlies the protective effect at the PRSS1–PRSS2 locus.
The identification of a polymorphism other than the one identified in the original studies as the causal variant may have diagnostic implications since the two variants of interest may not always be in perfect LD. We calculated LD between rs4726576 and rs10273639 from the 1000 Genomes project Phase 3 data and found that the degree of LD varies significantly among five meta-populations (figure 2B). The problem is most significant in the African meta-population. Based on our current finding, the TC (ie, rs10273639T in cis with rs4726576C) carriers, who accounted for 5.5% of subjects (figure 2C), should be assigned to carry a risk allele. However, they would have been assigned to carry a protective allele if the minor T allele of rs10273639 was thought to be causal.4
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
Footnotes
Contributors AB, J-MC and CF designed the overall project. AB performed in vitro experiments. MS performed mouse experiments. EM performed resequencing. EG analysed genotype data. AB, MS and J-MC drafted the manuscript. All authors revised and approved the final manuscript.
Funding This work was supported by grants from the Conseil Régional de Bretagne, the Association des Pancréatites Chroniques Héréditaires, the Association de Transfusion Sanguine et de Biogénétique Gaetan Saleun and the Institut National de la Santé et de la Recherche Médicale (INSERM), France.
Competing interests None.
Patient consent Obtained.
Ethics approval Ethical Committee of the University Brest; Animal Care and Experimentation Committee of Kagoshima University.
Provenance and peer review Not commissioned; internally peer reviewed.