Article Text

Statistics from Altmetric.com
Summary
M cells are highly specialised cells present within the epithelium overlying organised lymphoid follicles of the small and large intestine. They play a central role in the initiation of mucosal immune responses by transporting antigens and microorganisms to the underlying lymphoid tissue. In this way the mucosal immune system encounters the limitless variety of antigens that enter the body through the gut mucosa and reacts by mounting specific mucosal and systemic immune responses.
Despite the role of M cells in mucosal defence many basic aspects of their biology, the most controversial being their origin within the follicle associated epithelium (FAE), still remain the subjects of debate. Recently, new information on the complex interactions of luminal microorganisms, mucosal immune system, and epithelial cells, that are instrumental in the induction of this cell phenotype, have become available. Here, the most novel data and hypotheses on M cell genesis and function in the gut are reviewed and discussed.
The main task of the epithelium overlying mucosal surfaces of the intestinal tract is to provide an effective barrier to the vast majority of macromolecules and microorganisms present in the intestinal lumen. This is achieved by several means. Firstly, the epithelium is formed by cells joined by tight junctions that allow passage of water and ions but provide an effective mechanical barrier to macromolecules.1 ,2 Secondly, mucosal surfaces are covered by local secretions of mucus, secretory IgA antibodies, and by a thick glycocalix.3-6 These features and the closely packed carpet of microvilli present on absorptive cells prevent contact and binding of macromolecules and potential pathogens to the epithelium. On the other hand, the intestinal epithelium must also provide portals through which antigens and microorganisms are delivered to the intestinal immune system in order to induce immune responses. In fact, it is now established that antigenic penetration of epithelial barriers is the first critical step in the generation of protective mucosal and systemic immune responses.7 The ability of the intestinal epithelium to transport antigens and microorganisms is strategically restricted to the FAE8 that overlies the organised mucosal associated lymphoid tissue in the gut.9 To accomplish this task, the FAE has evolved features that distinguish it from the surrounding absorptive epithelium. The most remarkable adaptation is the presence of a relatively small number of highly specialised antigen sampling membranous (M) cells.10
Features of intestinal M cells
MORPHOLOGY AND FUNCTION
M cells were first observed by transmission electron microscopy in rabbit appendix.11 They display distinctive morphological features that distinguish them from surrounding enterocytes. The brush border is poorly organised with short irregular microvilli, and the thick glycocalix usually associated with absorptive cells is absent.12 ,13 These adaptations allow material in the intestinal lumen to have easy access to the apical domain of M cells where it is internalised and then transported to the underlying lymphoid tissue. The mechanisms by which M cells take up microorganisms and macromolecules vary according to the nature of this material. Large particles and bacteria induce phagocytosis, which is associated with ruffling of the apical plasma membrane of the M cell and rearrangement of the actin cytoskeleton, which permits active formation of pseudopod-like structures.14 ,15 Viruses and other adherent particles are taken up by endocytosis via clathrin coated vesicles,16 whereas non-adherent material is internalised by fluid phase endocytosis.11 ,17 In any case, internalisation is quickly followed by transport of endocytotic vesicles to the endosomal compartment and then by exocytosis to the basolateral membrane. The biochemical events involved in the intracellular transport of endocytotic vesicles in M cells have not been thoroughly explored but it appears that this is regulated in the same manner as polarised transport observed in other epithelial cells.18 ,19
A typical feature of M cells is that, unlike other intestinal epithelial cells, the basolateral surface is deeply invaginated to form intraepithelial pockets that are in intimate contact with specialised lymphocytes that migrate to this peculiar compartment from lymphoid tissue20 ,21 (fig 1A, B). This modification of the basolateral domain of M cells is believed to be a way of shortening the distance that endocytotic vesicles have to travel to reach immunocompetent areas.16 ,22 Thus M cell pockets provide the first opportunity for contact between antigens, penetrating the epithelial barrier and specialised immune cells. It has been proposed that M cell pockets are the sites where intraepithelial lymphocytes (IEL) can interact early with internalised antigens in an environment sequestered from regulatory elements of the mucosal immune system.22
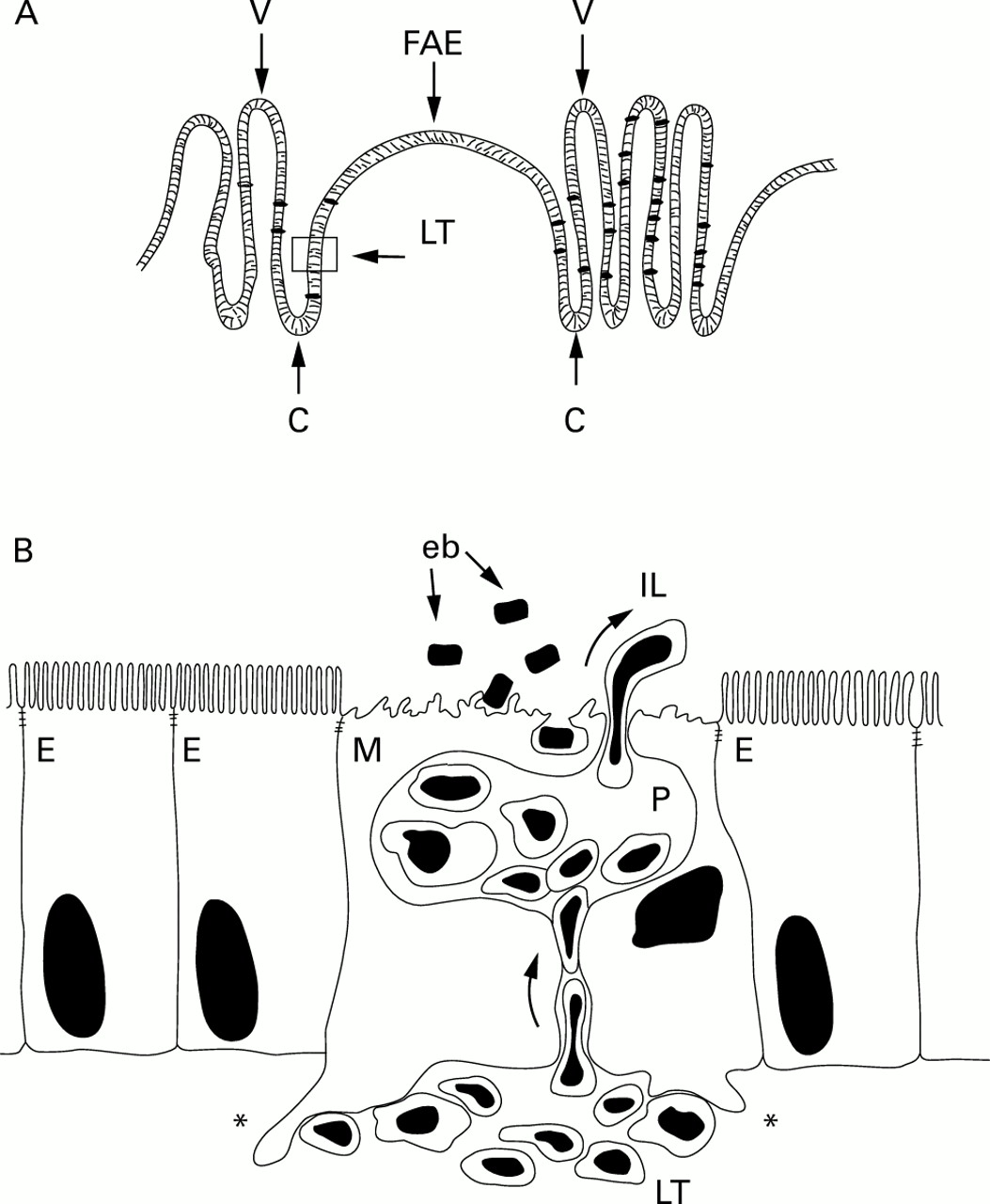
{kind=link}
In Peyer's patches, the lymphoid tissue (LT) is separated from the intestinal lumen (IL) by the follicle associated epithelium (FAE) (A). The cells forming the FAE originate in the closely associated crypts (C) and migrate upwards to the apical region of the dome. Lymphoid follicles are embedded within villi (V). Part of the FAE is indicated in the box (arrowhead) and enlarged (B) to show a typical M cell (M) with adjacent enterocytes (E). The basal membrane of the M cells deeply invaginates forming cytoplasmic pockets (P) harbouring lymphoid cells migrating into them from the lymphoid tissue. M cells also possess basal processes that deeply protrude into the lymphoid tissue (asterisks) where they make contact with cells of the immune system. Enterobacteria (eb) and other antigens adhere to the apical area of the M cells and are subsequently internalised and transported to the mucosal immune system. M cells are also a migration route for lymphocytes moving into the intestinal lumen. The function of these intraluminal cells, the number of which markedly increases after bacterial challenge, remains unknown.
SIMPLY ANTIGEN SAMPLING CELLS OR SOMETHING MORE?
The phenotype of IEL residing in M cell pockets has been analysed in Peyer's patch tissue of humans and other species.23-25 Although there is remarkable variation between animal species, most of the M cell associated IEL are α/β memory T cells. In the rabbit a distinctive phenotype lacking both CD4 and CD8 was also observed.26 T cells in the pockets are often associated with naive IgM+ and IgD+ B cells and macrophages, whereas IgG+ and IgA+cells have rarely been observed.27
The lymphoid cells harboured within M cell pockets represent all the cell types required to initiate a specific immune response but their true functional significance remains unclear. An immunological role for the M cell associated lymphocytes is suggested by the observation that their number rapidly increased after application of a bacterial challenge.15 ,28 In these circumstances, a large number of cells migrate into the intestinal lumen via M cell gated diapedesis.28 The physiological relevance of M cell regulated passage of lymphocytes into the gut lumen and their role in this location remain to be determined.
A broadly accepted view is that M cells are simply conduits through which antigens reach the immunocompetent areas of the gut associated lymphoid tissue. According to some authors M cells are provided with the cytoplasmic components required to process antigens,29including cathepsin E30 an aspartic proteinase involved in antigen processing by a B cell lymphoma.31 It is notable that expression of major histocompatibility complex class II molecules has been reported on the basolateral domain facing the pockets.27 ,29 ,31 M cells may also have the potential to participate actively in the generation of mucosal immune responses by signalling to lymphocyte by releasing IL-1.32 In addition, M cells possess basal processes that deeply protrude into the lymphoid tissue where they make contact with cells of the immune system.33 Taken together these data would suggest a more active and complex role of intestinal M cells in the early phases of mucosal immune responses.
HETEROGENEITY OF M CELL POPULATION
M cells were discovered more than 25 years ago11 but despite intensive investigations a clear cut M cell marker, independent of species and location in the gut, has not been found. For example, in the rabbit, but not in other species so far tested, M cells express the intermediate filament protein vimentin34-36 that is typical of cells of mesenchymal origin. In the past, M cells have been mainly identified by the absence of hydrolytic enzymes, such as alkaline phosphatase, that are abundant in the brush border of enterocytes.37-39 However, the reliability of alkaline phosphatase as a specific negative marker has been questioned by several authors on the basis of the extreme variability in the content of this enzyme in all FAE cells.18 ,40 More recently it has been reported that mouse M cells display a different distribution of the actin associated protein villin.41 This is restricted to the apical region of enterocytes but in M cells it is diffusely distributed in the cytoplasm reflecting a differentially organised cytoskeleton and probably a decreased structural rigidity.
The heterogeneity of M cell populations is also well exemplified by the highly differentiated expression of polysaccharides of membrane bound glycoproteins and glycolipids. The use of a large panel of lectins to stain FAE cells has revealed that in some cases M cells display a different glycosylation state compared with neighbouring enterocytes.42 ,43 In mice, for example, small intestine Peyer's patch M cells predominantly express α(1–2)-fucose that can be detected by Uleus europeaus (UEA-1) lectin.33 ,43 ,44 The pattern is different in caecal patches where M cells are stained by the same lectin but also express other terminal saccharides.42 ,45 In humans, the glycosylation pattern is different from that of other species as M cells preferentially display the sialyl Lewis A antigen.46
Membrane bound glycoconjugates are believed to play an important role in microbial-M cell interactions.47 In the light of this consideration the extreme variability of these molecules on the M cell surface has been interpreted as an effective way to generate a broad repertoire of M cell surface binding molecules to recognise bacteria borne lectins.22 ,47
M cell specific glycoconjugates have been successfully used as targets for oral and nasal delivery of antigens and particulate carrier.48 ,49 However, regional differences in the glycosylation pattern of M cells could influence the outcome of an antigen challenge. Studies in sheep have shown that the location in the gut in which antigen transport takes place may have profound effects on the immune responses.50 Both systemic and mucosal immune responses were observed when the vaccine vector was delivered to jejunal, but not ileal, Peyer's patches. In the latter case only a systemic response was observed.
M CELLS AS THE MAIN PORTAL OF ENTRY FOR ENTEROPATHOGENS
In contrast with the positive role of M cells in the initiation of mucosal immune responses, their capacity for antigen sampling can facilitate invasion by potential harmful intestinal microorganisms.51 The risk is mitigated by the fact that these pathogens are directly delivered to areas of the immune system fully equipped to cope with such an emergency. Nevertheless, some microorganisms exploit M cells as an entry site to breach the mucosal barrier and establish local and systemic infections.52 ,53The most dramatic evidence of such ability is provided by studies performed on Salmonella,54 ,55 Shigella,56 andYersinia 57 in different animal models. A detailed analysis of the various mechanisms used by these bacteria is beyond the scope of this article but has recently been reviewed elsewhere.58
Origin and fate of intestinal M cells: facts and hypotheses
Intestinal epithelial cell maturation and differentiation is a well known phenomenon which is completed in a geographically well organised migration.59-61 The epithelium of each follicle derives from surrounding crypts, each crypt being a clonal unit,62 ,63 which are characterised by two distinct axes of migration and differentiation. Cells located on the villus side of the crypt differentiate into absorptive enterocytes, goblet, and enteroendocrine cells. Cells on the FAE side of the crypt move onto the dome, acquiring features of follicle associated enterocytes and M cells.7 The final differentiation of intestinal epithelial cells takes place as they migrate in vertical bands to the apical extrusion zone of the villus and FAE.59 Within this scenario two different theories on M cell genesis in the FAE have been formulated. Although it is accepted that enterocytes and M cells have a common precursor it has been postulated that M cells may originate in the crypts as a distinct cell lineage from stem cells via an independent differentiation programme or from enterocytes on interaction with the local lymphoid microenvironment.
Initially, the hypothesis that M cells are derived from undifferentiated crypt cells was based on ultrastructural studies and on the utilisation of 3H thymidine as a proliferation marker. 3H thymidine labelled cells were observed in crypts adjacent to the dome13 and within 24 hours they migrated at the dome periphery where they acquired morphological and structural features of M cells.13 ,64 The most recent experimental evidence supporting this idea mostly comes from studies on differential expression of glycoconjugates on M cell membranes. It has been demonstrated, in both the rabbit and mouse, that lectin labelled M cells were not restricted to the dome of the FAE but were also detected in crypts.33 ,65 These data indicated that a subpopulation of crypt cells is predetermined as M cells before attaining their morphological and functional features. More recently another morphological and histochemical study has provided further evidence that supports this view.66 Here it was also determined that the random distribution of the sites where lymphocytes invade the FAE did not correlate with the organisation of M cells in radial strips that emerged from the dome associated crypts. These data led the authors to conclude that M cell formation is restricted in specialised dome associated crypts and that FAE associated enterocytes do not have the potential to convert into M cells.
In contrast, data from other groups point in a different direction. Originally, lymphocyte induced M cell formation was proposed by Smith and colleagues.39 ,67 Recently, an elegant in vitro co-culture system has been established in which murine Peyer's patch derived lymphocytes convert human intestinal Caco2 cells into functional M cells.68 Although it has been highlighted that due to the lack of terminal differentiation it is difficult to consider these adenocarcinoma cells as differentiated enterocytes,69 this model provides clear cut demonstration that lymphoepithelial cross talk plays a central role in controlling the genesis of differentially specialised epithelial cell populations. In addition, in vivo studies have provided evidence that bone marrow transplantation in SCID syngeneic mice induced the formation of M cells, thereby demonstrating that M cell genesis is immunoregulated.68 ,70 Furthermore, our group has reported that short term exposure (1–3 hours) to a non-intestinal bacterium induced dramatic alterations of the FAE that included a marked increase in the number of fully operational M cells in topographically restricted areas of the FAE.15 ,71 ,72 In this case the brevity of the experimental procedure demonstrated that the appearance of M cells, under certain experimental conditions, can take place in time scales too short to be explained by migration of M cells originated in crypts.22
Is it possible, in the light of these apparently conflicting data, to reconcile the two theories? One possibility is that the rapid increase in M cells after microbial challenge could be explained as short term differentiation of predetermined M cells.65 According to this hypothesis, these immature pre-M cells would elude detection because of lack of well defined ultrastructural and histochemical markers and certain antigenic challenges would provide the ultimate stimulus to drive their differentiation into mature operational M cells.65 The presence of cells within the FAE with the ability of rapidly becoming antigen sampling cells would explain the changes in the FAE following bacterial challenge. It also might be interpreted as an emergency system to boost immune responses by increasing the amount of antigenic substrate in the inductive sites of the mucosal immune system.71 ,72
The theory of lymphocyte induced M cell formation raises a question. How can lymphoid cells transform an enterocyte into an M cell? Indeed, while the genesis of M cells from undifferentiated crypt cells reflects the conventional differentiation pathway of other intestinal epithelial cells, the mechanism by which lymphocytes would induce the M cell phenotype remains a matter of speculation. The Caco2 co-culture system has provided clear evidence that B cells play a major role in the conversion of enterocytes.68 More recently, the role of B cells in the formation of both FAE and M cells has been confirmed by in vivo experiments involving B cell knockout mice.73 At this time, however, the biochemical and molecular events involved are unknown. It was proposed that lymphocytes must deliver two different signals to enterocytes, one with the ability to disassemble the brush border and the second to trigger transcytosis.68 This hypothesis was based on the observation that the sole disorganisation of the brush border in Caco2 cells, via suppression of villin by specific antisense RNA,68 ,74 was not sufficient to trigger transcytosis. Nevertheless, cells with morphological and functional features of M cells that were not in contact with lymphocytes were observed in other experimental systems.72 ,75 In one such case72 rapid appearance of operational M cells after bacterial challenge in vivo was preceded by an increase in the number of cells displaying intermediate phenotype between enterocytes and M cells. Most of these cells, probably transitional stages, did not harbor lymphocytes, thereby suggesting that under certain conditions the stimulus provided only by a soluble molecule(s) may suffice to induce the M cell phenotype without the need for physical interactions with lymphocytes.
Interestingly, M cells are characterised by their peculiar distribution in the FAE. They are relatively abundant in the periphery but totally absent at the apical region.13 ,72
The observation that M cells are never observed in the apical region of the FAE poses a new question. What is their fate? In view of the highly dynamic nature of the FAE it has been hypothesised that M cells may revert back to enterocytes as they reach the apical area of the FAE.41 This is also supported by the observation that no apoptotic cells were found at the sites where M cells are located76 thereby suggesting that M cells are not preferentially extruded at the periphery. Thus M cells may represent a transient and reversible phenotype.
The situation appears to be different in the rabbit model. Rabbit M cells have the advantage of expressing the intermediate filament vimentin34-36 and are easily identified within the FAE. Consistent with the distribution of M cells already observed in other animal species, vimentin positive cells are restricted at the periphery of FAE and are totally absent at the apical area.72Immunohistochemical analysis of the rabbit FAE revealed the presence of M cell debris in the lumen over the dome shoulders, where desquamation of M cells was also observed.77 This strongly suggests that M cells are extruded at the level of the shoulders. Furthermore, if they were to revert back to enterocytes as they migrate towards the apex of the dome, one would expect to observe an intermediate phenotype in the close vicinity of the apical, M cell free area. Instead, vimentin positive cells stop abruptly at the dome shoulders72 suggesting that, at least in the rabbit model, M cells are probably extruded at the periphery rather than at the top of the dome.
Concluding remarks
M cells are important in regulating access of microorganisms and antigens to areas of the intestinal immune system equipped to generate protective immune responses. However, much remains to be learnt of the biology of this cell. The importance of finding the best way to induce protective mucosal immune responses is highlighted by the accepted view that the systemic immune response is not adequate to fight the vast majority of pathogens to which we are exposed throughout life. It is believed that over 95% of human pathogenic microorganisms target host cells after crossing epithelial barriers. For this reason it would be highly desirable to induce specific immunity at the site of invasion. The need to potentiate immunological defence at the mucosal level is further stressed by the fact that in adult individuals the area of the mucosal surfaces reaches the impressive size of 400 m2.78 A better understanding of the mechanisms involved in the recognition and transepithelial transport of antigens, along with the molecular basis of M cell formation within the FAE, will represent an important step forward in the design of new strategies to improve oral delivery of biologically active compounds to the intestinal immune system.
Acknowledgments
I am indebted to M J Taussig for his friendly and valuable support. I also wish to thank P J Kilshaw and J P Kraehenbuhl for helpful comments and discussions. The work in our laboratory has been funded by the Ministry of Agriculture Food and Fisheries of the United Kingdom and by the Biotechnology and Biological Sciences Research Council.
Abbreviations
- FAE
- follicle associated epithelium
- IEL
- intraepithelial lymphocytes