Article Text

Statistics from Altmetric.com
- inflammatory bowel disease
- irritable bowel syndrome
- Crohn’s disease
- protease activated receptors
- ulcerative colitis
- thrombin
- tryptase
Of all the body systems, the gastrointestinal tract is the most exposed to proteinases. Under physiological conditions, digestive proteinases such as trypsin are released in pancreatic ducts and then into the upper gastrointestinal tract, during and after meals. The forms of proteinases released in the pancreatic duct and intestinal lumen are inactive and need to be cleaved to be activated. This cleavage occurs in the gut lumen where a constant balance between proteolytic activity and the presence of proteinase inhibitors persists in order to fulfil the digestive functions of the gastrointestinal tract, but also at the same time to protect mucosal surfaces from exposure to proteolytic enzymes. The intestinal lumen is also constantly exposed to proteases potentially released from the commensal flora, but also in pathophysiological conditions to proteinases released by infectious agents. Thus the importance of signalling pathways activated by proteinases appears to be particularly relevant for the physiology and pathophysiology of the gastrointestinal tract.
PARS: ACTIVATION AND DISTRIBUTION IN THE GUT
Proteinase activated receptors (PARs) are seven transmembrane domain receptors that are activated by specific proteolytic cleavage of their N terminal extracellular domain. This cleavage releases a new motif of amino acids on the N terminal tail which then binds the receptor like an agonist to induce an intracellular signal (see fig 1). The discovery of this family of receptors highlighted a new way of thinking about the role of proteinases, that cannot be seen merely as degradative enzymes, but that can directly and specifically send signals to cells.
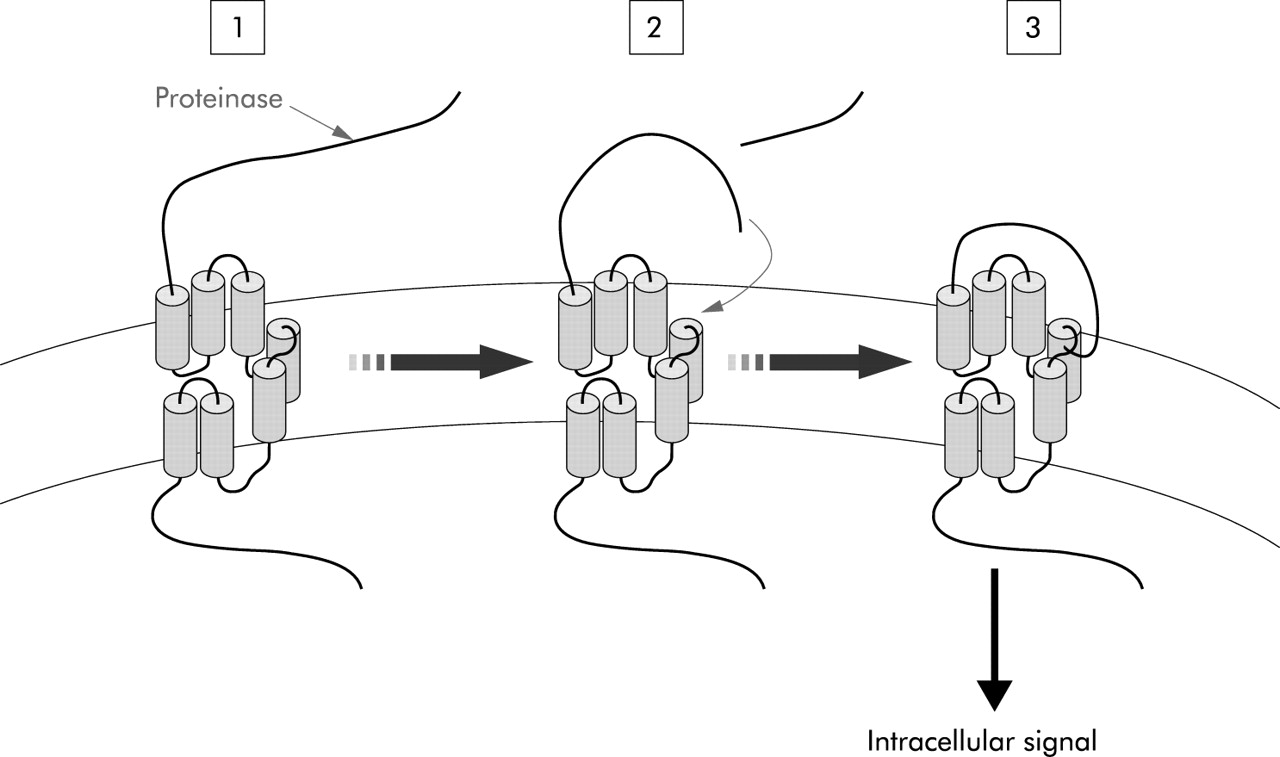
Mechanism of activation of proteinase activated receptors. Proteinases cleave the N terminal domain (1) releasing a new N terminal tail which acts as a tethered ligand that binds the receptor itself (2) to induce an intracellular signal (3).
Four members of the PAR family have been described: PAR1, PAR2, PAR3, and PAR4. Each of these receptors can be activated by the same proteinases: for example, thrombin can activate PAR1, PAR3, and PAR4, and trypsin can activate PAR2, PAR4, and to a lesser extend PAR1 and PAR3, or they can be activated by different proteinases: for example, tryptase activates only PAR2 (for review see Vergnolle1 and table 1). Selective agonists are pharmacological tools available to study PAR1, PAR2, and PAR4, but no selective agonist for PAR3 has been described, thus hampering the research on understanding the physiology of this particular receptor. The selective agonists available for PAR1, PAR2, and PAR4 are small peptides based on the amino acid sequence of the proteolytically released N terminal tail of the receptor, which acts as a tethered ligand (for review see Vergnolle1 and table 1).
Characteristics of the four members of the protease activated receptor (PAR) family: PAR1, PAR2, PAR3, and PAR4.
In the gut, PAR1 and PAR2 are expressed on endothelial cells of the lamina propria, on epithelial cells, smooth muscle cells, enteric neurones, fibroblasts, and on some immune cells (neutrophils, mast cells, lymphocytes, monocytes, macrophages). Expression of PAR2 varies in the gastrointestinal tract, with strong expression in the colon and small intestine and weaker detection in the stomach. PAR3 is expressed in the stomach and small intestine although the cell types that express the receptor still have to be identified. PAR4 is highly expressed in the small intestine and colon where immunoreactive protein was found on epithelial surfaces and submucosa (see table 1).2
ACTIVATION OF PARS AND GUT PHYSIOLOGY
Using PAR activating peptides to selectively activate PAR1, PAR2, and PAR4 in gut preparations, different responses have been observed, affecting all aspects of gut physiology (see fig 2 for summary).

{kind=link}
{kind=link}
Consequences of protease activated receptor (PAR) activation on gut physiological functions.
Ion transport
Activation of PAR1 and PAR2 has been shown to provoke chloride secretion in tissues or epithelial cells mounted in Ussing chambers. In the case of PAR1, direct activation of the receptor on enterocytes leads to chloride secretion.3 However, in whole tissues, PAR1 activation inhibited neurally evoked chloride secretion suggesting that PAR1 activation on enteric nerves might modulate ion transport across the epithelium. These results highlight the fact that depending on the cells activated in response to the PAR1 agonist, opposite effects may be obtained. For PAR2, its direct activation on enterocytes mobilises calcium, releases arachidonic acid and prostaglandins,4 and provokes chloride secretion. In whole tissues, chloride secretion was observed in response to PAR2 agonists.5,6 However, some discrepancies subsist on the mechanism involved in PAR2 induced chloride secretion. Results in rat jejunum suggest that nerve activation is not involved but prostaglandin release is a major mediator of PAR2 induced chloride secretion.6 However, in pig ileum and mouse colon, PAR2 induced chloride secretion depends on eicosanoids release and submucosal neurone activation.5,7 These differences may be explained either by different mechanisms existing between species or by differences between the different regions of the gut studied. In contrast with PAR1, chloride secretion resulting from PAR2 activation is always observed and does not depend on the cell type where the receptor is activated. In human mucosa, PAR2 agonists have also been shown to provoke chloride secretion by a mechanism independent of neurone activation but involving prostaglandin release.8
The functional and clinical implications of regulation of chloride secretion by PARs rely on two aspects. Firstly, stimulation of chloride secretion is associated with fluid transport and results in diarrhoea which constitutes an important clinical symptom of inflammatory bowel disease (IBD) or irritable bowel syndrome (IBS). Secondly, one of the functions of increased chloride secretion is to flush away from the mucosal surfaces potential pathogens. In that setting, increased chloride secretion would be protective against infectious diseases of the gut. In both cases, regulation of chloride and ion secretion by PARs has important clinical implications.
Permeability
Recent work has revealed that both PAR1 and PAR2 activation lead to a leakage in intestinal barrier functions, increasing the passage of fluids or even microorganisms across the gut mucosa.9,10 In the case of PAR1, this increased intestinal permeability was mediated by direct activation of the receptor on enterocytes. This PAR1 induced increased permeability was due to initiation of programmed cell death (apoptosis) at the mucosal surface.10 While there is still a large debate on the question of whether or not apoptosis can induce changes in permeability in vivo, this study showed for the first time that blockade of the apoptotic cascade inhibited PAR1 induced increased permeability both in vitro and in vivo.10 PAR2 induced increased intestinal permeability was also mediated, at least in part, by activation of the receptor on enterocytes but was not linked to activation of the enteric nervous system.11 Rearrangements of cytoskeletal organisation of the tight junctions between epithelial cells were observed in response to PAR2 agonists, thereby explaining the passage of macromolecules across the epithelial barrier. In vivo, this PAR2 induced increase in intestinal permeability even allowed the passage of bacteria from the gut lumen to peritoneal organs,9 which could thus participate in a general inflammatory response.
Motility
PAR1, PAR2, and PAR4 have been shown to be expressed on intestinal smooth muscle.2,12 Both PAR1 and PAR2 activation caused contraction of the resting rat and mouse gastric smooth muscle in a prostaglandin dependent manner.13 In the mouse gastric fundus, responses to PAR activation were biphasic, showing relaxation that was able to mask a concomitant contractile effect.14 In longitudinal muscles of the colon, as well as in circular muscles, PAR1 and PAR2 activation inhibited contractile effects.15–17 The effects of PAR4 activation have only been studied in isolated organs to date, and a recent study has shown that PAR4 agonists produced a concentration dependent contractile effect on longitudinal muscle.2 While the effects of PAR agonists on isolated gut organs resulted in opposite effects, whether or not the organ was precontracted, an in vivo study by Kawabata et al definitively established the resultant effects of PAR1 and PAR2 agonists on gut motility.18 In vivo, administration of PAR1 and PAR2 agonists increased gastrointestinal transit18 thus showing that PAR activation can alter motility patterns. The resultant effect of PAR activation on gut motility might depend on the cellular target that is first reached by PAR agonists. Smooth muscle cells have been shown to be responsive to PAR agonists. However, myenteric neurones have also been shown to be responsive to PAR agonists,19,20 and activation of PARs on those fibres might be responsible for motility dysfunctions associated with disease. This is further supported by the fact that most of the contractile responses to PAR agonists were desensitised by neurokinin receptor antagonists, which suggests previous neural activation in response to PAR agonists.2,17,21
Inflammation
Intraluminal administration of PAR1 or PAR2 agonists into the colon of mice provoked an inflammatory reaction characterised by the formation of submucosal oedema and infiltration of granulocytes.9,22 In the case of PAR1, it was shown that this inflammation was not due to activation of sensory nerves or to their release of neuropeptides.22 However, PAR1 induced colitis was partly due to an increase in intestinal barrier permeability but also to direct activation of B and T lymphocytes,22 thereby implicating PAR1 in potential gut immune functions. Activation of PAR1 was able to exacerbate chronic inflammation in a mouse model of IBD.22
PAR2 activation induced colitis in mice was clearly mediated by activation of enteric nerves and subsequent release of neuropeptides: calcitonin gene related peptide (CGRP) receptor antagonists and neurokinin 1 receptor antagonists were able to completely suppress PAR2 agonist induced inflammation.11,23 This result implies that PAR2 activation on enteric neurones provokes the release of neuropeptides such as substance P and CGRP, which in turn provoke an acute inflammatory reaction. However, in the context of chronic gut inflammation, activation of the enteric nervous system is known to have protective effects.24–26 This suggests that PAR2 induced enteric nerve activation could exert anti-inflammatory properties against chronic inflammation. This hypothesis has been confirmed by a study by Fiorucci et al who have shown that systemic treatments with PAR2 activating peptides significantly reduced trinitrobenzene sulfonic acid induced colitis in mice.27 In that study, the authors suggested that the anti-inflammatory effects of PAR2 agonist treatments were mediated by CGRP release.27 As mentioned earlier, PAR2 agonists from the lumen of the colon are also able to increase intestinal barrier permeability, and this effect could, to a certain extent, participate in generation of inflammatory disorders and may counteract the protective effects of enteric nervous system activation in the chronic context of inflammation.
Nerves, epithelial layers, or immune cells are probably not the only targets to explain the proinflammatory effects of PAR1 and PAR2 agonists. Numerous studies have demonstrated the effects of PAR1 and PAR2 agonists on endothelial cell permeability,28,29 and PAR activation induced endothelial permeability may also explain, in a large part, oedema formation in response to PAR agonists.
Sensory functions
Because PAR1 and PAR2 are expressed and functional on afferent sensory fibres in the gut,1 a role for these receptors in nociception mechanisms has been suggested. It has been shown that intracolonic administration of subinflammatory doses of PAR2 agonists (trypsin or selective PAR2 activating peptide) provoked longlasting hyperalgesia in response to colorectal distension in rats.30 Furthermore, the same intracolonic treatment provoked nociceptor activation at the spinal level in the superficial laminae (laminae I and II) of the dorsal horn, thereby showing that colonic activation of PAR2 provoked pain.30 Whether this PAR2 induced pain symptom is due to direct activation of PAR2 on sensory neurones is still debatable. However, studies have shown that PAR2 agonists evoked transient depolarisation of submucosal neurones, followed by longlasting hyperexcitability of those neurones.31 Another study has shown that PAR2 agonists were able to cause increases in jejunal afferent firing and intrajejunal pressure in rats,32 thereby demonstrating a direct effect for PAR2 activation on intestinal afferent nerves.
In contrast with PAR2, subinflammatory doses of PAR1 agonists provoked analgesia when injected into the colon lumen or the peritoneal cavity.33,34 The same effect was observed at a somatic level when PAR1 agonists (thrombin or selective PAR1 activating peptide) were injected into the rat paw.35 Whether this effect is due to direct activation of PAR1 on enteric afferents has still to be determined.
Proliferation
Most recently, expression and function of PARs in human colon cancer cell lines has been studied. A study by Darmoul et al has shown that PAR2 was expressed in six of 10 colon cancer cell lines tested. Moreover, trypsin and a selective PAR2 activating peptide induced an important proliferative response in different colon cancer cells.36 PAR1 expression was found in 11 of 14 colon cancer cell lines tested. Similarly, PAR1 expression was detected in human colon tumours but was absent in normal human colonic epithelial cells.37 In colon cancer cell lines, PAR1 was functional, inducing calcium mobilisation and proliferation of cancer cells.37 A recent study indicated that activation of both PAR1 and PAR2 could act in synergy to induce migration of transformed cells.38 Taken together, these results suggest that proteases, through activation of PAR1 and PAR2, should be considered as growth factors and potential actors of mitogenic and metastatic events associated with colon cancer.
CLINICAL IMPLICATIONS
The effects of PAR agonists on all the aspects of gut physiology (fig 2) strongly suggest an important role for these receptors on gut functions and thereby a potential role in disease associated gut dysfunctions. Defining clearly clinical implications for PARs in gut diseases relies on the use of antagonists or inhibitors that would specifically interact with activation of these receptors. For PAR1 and PAR4, such drugs have recently started to be available and have already raised some answers. However, for PAR2, such a drug is still not readily available. Several lines of evidence suggest a role for PARs in IBD, IBS, infectious colitis, colon cancer, and fibrosis.
Inflammatory bowel disease (IBD)
As discussed earlier, acute activation of PAR1 and PAR2 in the colon of mice provoked an inflammatory reaction involving different mediators and different target cells. Does that mean that PAR1 and PAR2 activation could be implicated in inflammatory diseases of the gut such as Crohn’s disease (CD) or ulcerative colitis (UC) (table 2)?
Protease activated receptors (PARs) and inflammatory bowel disease
We have recently used PAR1 antagonist treatments in two different mouse models of inflammatory bowel disease. In this study, we showed that blockade of PAR1 activation significantly decreased the mortality rate and severity of inflammatory parameters in mice. Furthermore, we used a genetic approach investigating the effects of colitis induction (by intracolonic administration of trinitrobenzene sulfonic acid or by dextran sodium sulphate treatment) in PAR1 deficient mice. We confirmed that PAR1 deficiency was beneficial to improve clinical signs of inflammation in those two models of IBD.22 Expression of PAR1 was found to be upregulated in biopsies from CD or UC patients, further suggesting an important role for PAR1 activation in IBD. Finally, this study highlighted the fact that PAR1 activation on monocytic cells of the lamina propria is most likely responsible for activating the inflammatory/immune response. Taken together, these results imply an important role for PAR1 in the pathogenesis of experimental colitis, supporting the notion that PAR1 inhibition may be beneficial in the context of IBD. Permeability disorders have been associated clinically with the development of IBD39 as PAR1 activation also led to increased permeability.10 PAR1 induced increased permeability on enterocytes may also participate in the development of IBD. As thrombin is the most recognised activator of PAR1, and because coagulation disorders and thrombosis have been implicated in the pathogenesis of IBD,40,41 thrombin appears to be a good candidate to activate PAR1 in IBD. However, cathepsin G, which is largely released by activated neutrophils, can also activate PAR1 and could account for endogenous PAR1 activation during the course of the disease, thereby amplifying and prolonging the inflammatory response.
While acute PAR2 activation in the colon of mice induced colitis, two separate studies have shown that this inflammatory reaction involves a neurogenic component.11,23 Since in the context of chronic inflammation of the gut, activation of the enteric nervous system is considered to be protective,11,25,26 PAR2 activation in IBD patients might exert an anti-inflammatory effect. This is further suggested by the study of Fiorucci et al who showed that PAR2 agonist treatment reduced inflammatory parameters in a model of chronic colitis in mice.27 PAR2 expression was increased in biopsies from UC patients,27,42 suggesting a role for this receptor in IBD. Whether this role is protective or proinflammatory still needs to be investigated in depth, specifically using PAR2 deficient mice and/or PAR2 antagonists when they are available. However, the last study demonstrated that PAR2 was overexpressed in mast cells in tissues from UC patients and that PAR2 activation in human mast cells induced release of tumour necrosis factor (TNF).42 Thus it can be hypothesised that activation of PAR2 on mast cells and subsequent release of TNF might be implicated in the pathogenesis of UC. In that setting, blockade of PAR2 could be beneficial for the treatment of IBD. Similar to PAR1, PAR2 activation on epithelial cells induced increased permeability,9 and considering the potential involvement of permeability disorders in IBD,39 PAR2 activation in this context may also participate in pathogenesis. Taken together, these studies show that pro- or anti-inflammatory effects could be expected in response to PAR2 activation in the gut, depending on the cell where the receptor is activated. Thereby, in the absence of studies using a PAR2 antagonist, it is difficult to predict any clear advantage for PAR2 related drug in the treatment of IBD. It is interesting to note that an open label pilot study has investigated the effects of a tryptase inhibitor in UC patients, and this study reported beneficial effects for inhibition of tryptase. As tryptase exerts some of its effects through activation of PAR2, this could point to a proinflammatory role for PAR2 activation in IBD. Another enzyme that might be responsible for activation of PAR2 in the colon is trypsin, the activity of which has been shown to be increased in the tissues and lumen of IBD patients.43–45 Better knowledge of the endogenous enzymes that are responsible for PAR activation could contribute to the development of selective protease inhibitors as new therapeutic options for IBD.
Infectious colitis
A recent study has shown that PAR1 agonist induced chloride secretion was altered after nematode infection thereby suggesting that PAR1 might be implicated in infectious intestinal diseases.46 Another fact that supports this hypothesis comes from the observation that proteinases from pathogens could activate PARs. Gingipains, which are arginine specific proteinases released by the buccal pathogen Porphyromonas gingivalis, can activate PAR1, PAR2, and PAR4.47 Although it has never been shown for intestinal pathogens, but because of the strong expression of PARs on gut epithelial mucosa, it is reasonable to think that these receptors could be activated by pathogen proteinases in the gut. Considering the rather “inefficient” mechanism of activation of PARs (these receptors are one shot receptors, activated only once by proteolytic cleavage of their N terminus and then internalised for degradation—reviewed by Ossovskaya and Bunnett48) they can be viewed as a first line of defence against pathogen infection. Their activation by proteases released from pathogens would be part of the innate immune response, organising an immediate inflammatory reaction which would aim at fighting the infection. The fact that PAR2 induced colitis is mediated by a neurogenic mechanism supports this hypothesis because infectious colitis is known to involve enteric nerve activation and release of neuropeptides.49 If PARs are truly activated on gut infection by intestinal pathogens, blockade of PAR activation in the gut could be beneficial in counteracting disproportionate inflammatory responses to pathogens. However, we need to consider the possibility that PAR activation at mucosal surfaces could be a surveillance mechanism for pathogen infection, and its blockade would lower the capacity of the body’s response to infection, thereby rendering the patient more susceptible to infectious diseases.
Irritable bowel syndrome
Several aspects of gut physiology are involved in the generation of IBS symptoms: altered motility patterns, inflammatory mediator release, loss of intestinal barrier integrity, ion transport dysfunctions, altered nociceptive functions, and even psychological factors (for review see Mayer and Collins50). With the exception of the latter, PAR activation has been shown to be able to modify all of these functions (table 3).
Protease activated receptors (PARs) and irritable bowel syndrom
As previously discussed, several studies have reported the effects of PAR agonists on motor functions of the gastrointestinal tract, suggesting that PAR activation could participate in motor dysfunctions associated with IBS symptoms.
The putative role for microinflammation in tissues of IBS patients has been largely discussed and is supported by numerous studies.51 As both PAR1 and PAR2 activation have been shown to provoke an inflammatory reaction that was rather discrete compared with inflammatory models of IBD, and that did not last longer than 48 hours,9,22 this bout of inflammation might serve as a basis for generation of IBS symptoms. Development of IBS is often associated with a previous history of infectious colitis.49,51–53 The potential role for PAR1 and/or PAR2 activation as initial inflammatory events for the development of IBS symptoms is even more supported by the fact that PARs can be activated by pathogen proteinases.
Loss of intestinal barrier integrity has been associated with IBS in child and adult patients.53,54 This suggests that intestinal barrier dysfunction may play a role in the generation of IBS symptoms. As both PAR1 and PAR2 activation provoked an increase in intestinal barrier permeability, here again this could serve as a basis for generation of IBS symptoms.
IBS is often associated with either diarrhoea or constipation. Electrolyte transport across the intestinal barrier regulates water movements in the intestinal lumen, thereby participating in the generation of diarrhoea symptoms. Both PAR1 and PAR2 activation modify chloride secretion,3,6 suggesting that both receptors could be implicated in secretory dysfunctions associated with IBS. However, in contrast with PAR2, PAR1 activation in whole intestinal tissues was shown to inhibit neurally evoked chloride secretion, indicating a differential role for PAR1 activation depending on the cells that are activated. Thus in IBS, PAR2, but not necessarily PAR1, may participate in generation of diarrhoeal symptoms.
The stronger evidence for the role of PARs in generation of IBS symptoms relies on the properties of PAR activation on visceral nociceptive functions. Both cellular and animal model approaches have shown that PAR2 activation provoked longlasting visceral hypersensitivity and hyperexcitability of enteric neurones,30–32 suggesting an important role for PAR2 activation in visceral hypersensitivity states. The consequences of PAR2 activation coincide with all of the pathophysiological factors associated with generation of IBS symptoms (altered motility, microinflammation, increased permeability, increased chloride secretion, pain symptoms), strongly suggesting PAR2 activation in IBS. Although PAR1 activation altered motility and intestinal permeability and can induce release of inflammatory mediators, PAR1 activation also provoked analgesia, suggesting an inhibitory role for PAR1 in pain symptoms associated with IBS. These conflicting results on the role of PAR1 may be explained by the different cellular targets involved in each function.
Here again the question of the endogenous protease responsible for PAR activation in the setting of IBS is crucial. Tryptase appears as an obvious candidate for activation of PAR2 considering the involvement of mast cells in IBS. Recently, a study by Barbara et al has shown that in IBS patients, the distance between mast cells and enteric nerves was significantly reduced compared with control patients.
Colon cancer
Proteinases in general, and trypsin-like proteinases in particular, are extremely abundant in the vicinity of tumours. Moreover, most of the colon cancer cell lines express both PAR1 and PAR2, activation of which is known to induce proliferation in those cells.36,37 Thus it is reasonable to believe that PAR1 and PAR2 can be activated on transformed epithelial cells during colon cancer, and that this activation might participate in the proliferation of cancer cells (table 4). In the case of PAR1, because thrombin and selective PAR1 activating peptide led to a twofold increase in cell motility of wounded colon carcinoma cells,37 this suggests further that PAR1 activation could be involved in metastatic events. In that setting, inhibition of PAR1 and/or PAR2 activation would have beneficial effects against tumour progression in colon cancer. Whether or not PAR activation participates in proliferation and metastasis in animal models of colon cancer has still to be investigated to provide clear answers on the role of PARs in colon cancer. Procoagulant activity and platelet activation have been shown to enhance the ability of tumour cells to spread via the circulation, and colonise other organs inducing metastasis.55 Because PAR1, PAR3, and PAR4 are all implicated in thrombin signalling to platelets, a recent study using PAR deficient mice has investigated their role and the role of platelets in the formation of lung metastases after injection of melanoma cells.55 The authors reported that PAR1 and PAR2 deficient mice were not protected against metastasis. However, PAR4 deficiency conferred some protection against metastasis in this model. Similar studies would be necessary in a model of colon cancer to definitively conclude on the potential effects of PARs on proliferation and metastasis mechanisms.
Protease activated receptors (PARs) and colon cancer
Fibrosis
The presence of activated mast cells in the gut correlates with fibrotic disorders such as collagenous colitis, intestinal strictures observed in CD, or radiation induced fibrosis. As tryptase, the major protein present in mucosal mast cell granules and released on mast cell activation, can signal through PAR2, it is likely that PAR2 is activated in fibrotic disorders of the gut. One study performed with mucosal biopsies collected from cystic fibrosis patients supports this hypothesis. In that study, the authors observed that cystic fibrosis tissues were no longer able to activate chloride secretion in response to PAR2 stimulation, suggesting that the receptor had been desensitised.8 Activation of PAR2 has been shown to reproduce proliferative effects of tryptase on lung fibroblasts and myocytes, thereby suggesting an important role for this receptor in lung fibrosis. Given the strong resemblance between lung and gut mucosal physiology, a role for PAR2, which has been shown to be present on enteric fibroblasts and smooth muscle cells, could also be suggested in gut fibrotic disorders.
Fibrotic disorders are also associated with excessive deposition of extracellular matrix proteins and ongoing coagulation cascade activity. Proteinases of the coagulation cascade such as thrombin, factor VIIa, and factor Xa can activate the two thrombin receptors PAR1 and PAR4. Activation of PAR1 by thrombin and factor Xa has been shown to stimulate expression of connective tissue growth factor in fibroblasts.56 Moreover, in a model of radiation induced fibrosis in the gut, PAR1 was overexpressed in smooth muscle cells and its expression correlated with fibrotic damage.57
Taken together, these experimental approaches suggest a role for PAR1 and PAR2 in fibrotic disorders associated with gut pathologies such as radiation injuries, collagenous colitis, or stricture formation observed in IBD. However, such a role for PARs in gut fibrosis has still to be delineated and clearly established by investigating the consequences on gut fibrosis of receptor blockade.
CONCLUDING REMARKS
There have been substantial advances in our understanding of the role of proteinases as signalling molecules to cells. Particularly in the gut, which is the organ most exposed to proteinases, it appears that proteinases, through activation of PARs, can interfere with most of the physiological functions of the gut. PAR activation can participate in inflammatory reactions, be protective to mucosal surfaces, send or inhibit nociceptive messages, modify gut motility or secretory functions, and stimulate cell proliferation and motility. It is still difficult to clearly define the role of PARs in human gut diseases, mostly because of the lack of pharmacological tools to efficiently block in vivo activation of these receptors. However, studies that combined in vitro and in vivo approaches using pharmacological and genetic tools clearly pointed to a role for PAR1 in IBD. Strong evidence also supports a role for PAR2 in symptoms associated with IBS. In vivo approaches are still needed to fully investigate the role of PARs in fibrosis, colon cancer, or even infectious colitis.
Another important aspect that needs to be investigated to propose a therapeutic approach involving PAR related drugs would be to define which proteases are responsible for PAR activation in the setting of a particular disease.
In conclusion, it appears that recent advances in basic science point to a crucial role for proteinases and their receptors in gut functions. We need to consider proteinases not only as digestive or degradative enzymes in the gut, but as signalling molecules that actively participate in several clinical symptoms of gut diseases.
Acknowledgments
This work was supported by grants from the Crohn’s and Colitis Foundation of Canada, by the Canadian Association of Gastroenterology, and by the Canadian Institute for Health Research. Nathalie Vergnolle is an Alberta Heritage Foundation for Medical Research Scholar and a Canadian Institute for Health Research New Investigator.
REFERENCES
Footnotes
-
Conflict of interest: None declared.