Article Text

Statistics from Altmetric.com
In the first half of the 20th century, white adipose tissue (WAT) was mainly viewed as an isolated tissue protecting the organism from heat loss and a passive energy storage compartment. Similarly to other species such as Drososophila melanogaster, it is now well recognised that mammalian fat tissue is not solely a reservoir for excess nutrients but also an active and dynamic organ involved in the development of metabolic syndromes and the regulation of immunity and inflammation. The older anatomical literature repeatedly mentions a close association between adipose tissue and lymphoid organs in various mammals including humans, suggesting a potential role of WAT in the host immune response. Several recent studies indicate that adipocytes could function as macrophage-like cells1 as they express receptors related to the innate immune system and secrete major mediators of inflammation, such as tumour necrosis factor alpha (TNFα). Consistent with this hypothesis,2 the biology of adipocytes is particularly implicated in chronic diseases, such as obesity3 and atherosclerosis.4
This review will focus on the normal and pathophysiological functions of mesenteric WAT (mWAT), which may play an important role in the inflammatory and fibrotic processes in Crohn’s disease, a frequent and complex form of inflammatory bowel disease (IBD).
ENDOCRINE AND IMMUNE FEATURES OF ADIPOCYTES
Long considered as the “anatomists’ Cinderella”,5 mWAT is now recognised as a multifunctional organ. Notably located around organs such as the gut or the lungs, adipocytes may have evolved strategies to drive immune responses to microbial invaders by expressing different innate immune sensors. In addition its function as a storage organ, WAT plays a major endocrine and immune role by expressing several hormones and various mediators (fig 1⇓). To clarify the nomenclature, we will refer to the hormones and immunomodulatory molecules derived from adipocytes as adipormones and adipocytokines, respectively.

Adipormones, adipocytokines and their receptors. The major mediators (A) and receptors (B) expressed in adipocytes are listed.
Adipocyte-derived hormones (adipormones)
Figure 1A⇑ lists the main adipormones, three of which have critical roles in the regulation of inflammation.
Leptin, also named Ob protein, is mainly produced by adipocytes in direct proportion to the fat mass, thus ensuring long-term control of food intake.6 The crystal structure of the human Ob protein revealed a four-helix bundle sharing similarities with the pro-inflammatory interleukin (IL)-6.7 Interestingly, leptin was shown to polarise in vitro the immune response towards a proinflammatory Th1 cytokine profile.8 In addition, several in vivo studies have extended the role of this hormone from energy regulation to pro-inflammatory functions, as illustrated in various experimental models of colitis in mice.9–,11
Unlike leptin, adiponectin is a 244 amino acid protein, exclusively expressed in adipocytes at a level inversely proportional to fat mass. Adiponectin (also known as Acrp30, GBP-28, apM1 and AdipoQ) is an adipormone with anti-atherogenic, anti-diabetic and insulin sensitising properties.12 Furthermore, several results suggest anti-inflammatory properties of adiponectin in macrophages and endothelial cells.13 However, the physiological role of adiponectin in inflammation in the gut remains elusive (see below).14
Resistin, also referred as FIZZ (“found in the inflammatory zone”), is a member of the cysteine-rich secretory protein family. Interestingly, the cellular sources and functions of resistin differ between humans and rodents. Widely expressed in rodent by adipocytes, recent evidence suggests that resistin plays a role in the murine pathogenesis of obesity and diabetes.15 However, the pathophysiological role of resistin in human metabolic syndromes needs to be further clarified. The human resistin is mainly produced by leukocytes and macrophages16,17 and is involved in the modulation of inflammation through an upregulation of chemokines and adhesion molecule production by endothelial cells.16,18 Therefore, it would be worth investigating the possible role of resistin in IBD.
Finally, other mediators such as angiotensinogen, plasminogen activator inhibitor-1 (PAI-1), visfatin, adisin and vaspin are produced by adipocytes. Whereas angiotensinogen and PAI-1 are involved in events associated with excess fat, such as increased blood pressure and thrombosis,19 the physiological role of visfatin, adisin and vaspin remains poorly understood.20
Other bioactive molecules secreted by adipocytes: adipocytokines
Cytokines, chemokines and ADAM (a disintegrin and metalloprotease) proteins are critical mediators of the immune and inflammatory responses involved locally in communication, activation and recruitment of cells in tissues. Like macrophages and epithelial cells, adipocytes are able to synthesise inflammatory (TNFα, IL-6, IL-1β, IL-18) and anti-inflammatory (TGFαβ, IL-10, IL-1RA) cytokines, as well as chemokines (IL-8, MCP-1, MIP1α, MIF), growth factors (M-CSF, HGF, VEGF, FGF2, FGF10, NGF) and ADAM proteins. These recent observations have transformed our classical view of fat from being a storing site to being a complex endocrine tissue regulating metabolic functions and inflammation (fig 1⇑).21 TNFα and IL-6 have received greater attention compared to other adipocytokines. In humans, mWAT is an important source of cytokines as it produces about 30% of circulating IL-6, mainly from adipocytes.22 Besides their roles in inflammation, TNFα and IL-6 are associated with decreased body fat mass, insulin sensitivity and lipoprotein lipase expression, supporting the view that inflammation underlies metabolic diseases.23
Adipocytes: sensors of microbial products
Recent advances in the characterisation of the biology of adipocytes have shed new light on the role of WAT in host defence against enteropathogens. Notably, preadipocytes and macrophages shared particular gene expression patterns.1 Furthermore, both cell types possess phagocytic and anti-microbial activity.24 Given the ability of preadipocytes to differentiate into macrophages,24 we hypothesised that adipocytes might regulate innate immunity.
Like macrophages, adipocytes could detect systemic or local Gram-positive and Gram-negative bacteria since they expressed two pattern-recognition molecules (PRMs), namely the Toll-like receptors (TLR)-2 and -4. As shown by the marked lipopolysaccharide (LPS)-induced expression of several genes such as TLR-2 and TNFα, the 3T3-L1 preadipocytes are responsive to the TLR4 ligand LPS.25–,27 Furthermore, human adipocytes express the human CD14 protein, which plays a crucial role in LPS-induced signalling pathways.26 More recently, we found in 3T3-L1 cells and in the mWAT of healthy individuals physiological expression of two cytoplasmic receptors called nucleotide-binding oligomerisation domain proteins (NODs), NOD2/CARD15 and NOD1/CARD4, involved in sensing unique bacterial peptidoglycan motifs derived from Gram-positive and Gram-negative bacteria.28
Similarly to bacteria, viruses may infect adipocytes, as they expressed CD4, CXCR4 and CCR5 receptors targeted by the human immunodeficiency virus (HIV)-1.29 Furthermore, adenovirus 36 enhanced differentiation in mature adipocytes30 and increased adiposity in experimentally infected chickens, mice and marmosets (non-human primates).31 Conversely, certain HIV protease inhibitors known to interfere with HIV’s ability to enter cells altered the differentiating process in adipocytes.32 Taken together, these observations might explain hyperlipidaemia, insulin resistance and changes in fat tissue distribution reported in HIV patients treated with these drugs.33
However, even if visceral adipocytes express microbe-sensing receptors, it is still unknown if and which micro-organisms reach mesenteric adipose tissue and naturally infect adipocytes. To explore this issue, we have recently compared in vivo the rate of bacterial translocation in mesenteric lymph nodes and mesenteric adipocytes in healthy mice and humans. Viable bacteria were found in about 20% of mesenteric lymph nodes and mWAT, showing for the first time the physiological presence of bacteria within mWAT.28 Despite a similar frequency of bacterial translocation between mesenteric lymph nodes and mesenteric adipose tissue, a 17-fold increased number of viable bacteria were found in mWAT compared to mesenteric lymph nodes.28 In other words, about 95% of the total viable bacteria cultured from mesenteric tissues are physiologically located in adipocytes and only 5% are translocating in mesenteric lymph nodes, indicating that adipocytes might be a main reservoir of bacteria in the mesentery.
All these observations fuelled speculation about the potential roles of mesenteric mWAT in the development of Crohn’s disease by reacting to the microbial environment and by initiating and/or promoting local inflammatory reactions by autocrine and/or paracrine modulation of adipocytes. Alternatively, mWAT might control visceral inflammation through interactions with mesenteric lymph nodes and/or disseminated infiltrating cells.
MACROSCOPIC AND HISTOLOGICAL CHARACTERISTICS OF MESENTERIC FAT IN CROHN’S DISEASE
Although pathological surgical specimens display phenotypic variation, Crohn’s disease is often recognised on the macroscopic appearance of intestinal lesions. Surgical assessment of the intestine in Crohn’s disease revealed that the mesentery is often thickened and stiff, with WAT overgrowth.34–,36
Considered as a hallmark of Crohn’s disease, fat-wrapping is defined as an mWAT extension from the mesenteric attachment and partially covering the small and large intestinal circumference in association with loss of the bowel-mesentery angle (fig 2A⇓).34–,36 Dr Burrill B Crohn himself mentioned this characteristic feature of mesenteric adipose tissue as a consistent symptom of the disease.37 The prevalence of fat abnormalities in Crohn’s disease has not been formally assessed by population-based studies. In a consecutive and unselected group of 27 intestinal resections performed on 25 patients with Crohn’s disease confirmed by histology, fat-wrapping was identified in 12 of 16 ileal resections and in seven of 11 large bowel resections.34 It correlated with transmural inflammation, and there was a significant relationship between fat-wrapping and other connective tissue changes, including fibrosis, muscular hypertrophy and stricture formation. Finally, a retrospective review of 225 small intestinal resections suggested that fat-wrapping is a hallmark of Crohn’s disease. More precisely, there was evidence of fat-wrapping in 31 of 58 cases, but it was never observed in other conditions, including ischaemia or infarction, Meckel’s diverticulum, carcinoma or lymphoma, perforation of various causes, radiation enteritis and carcinoid.34
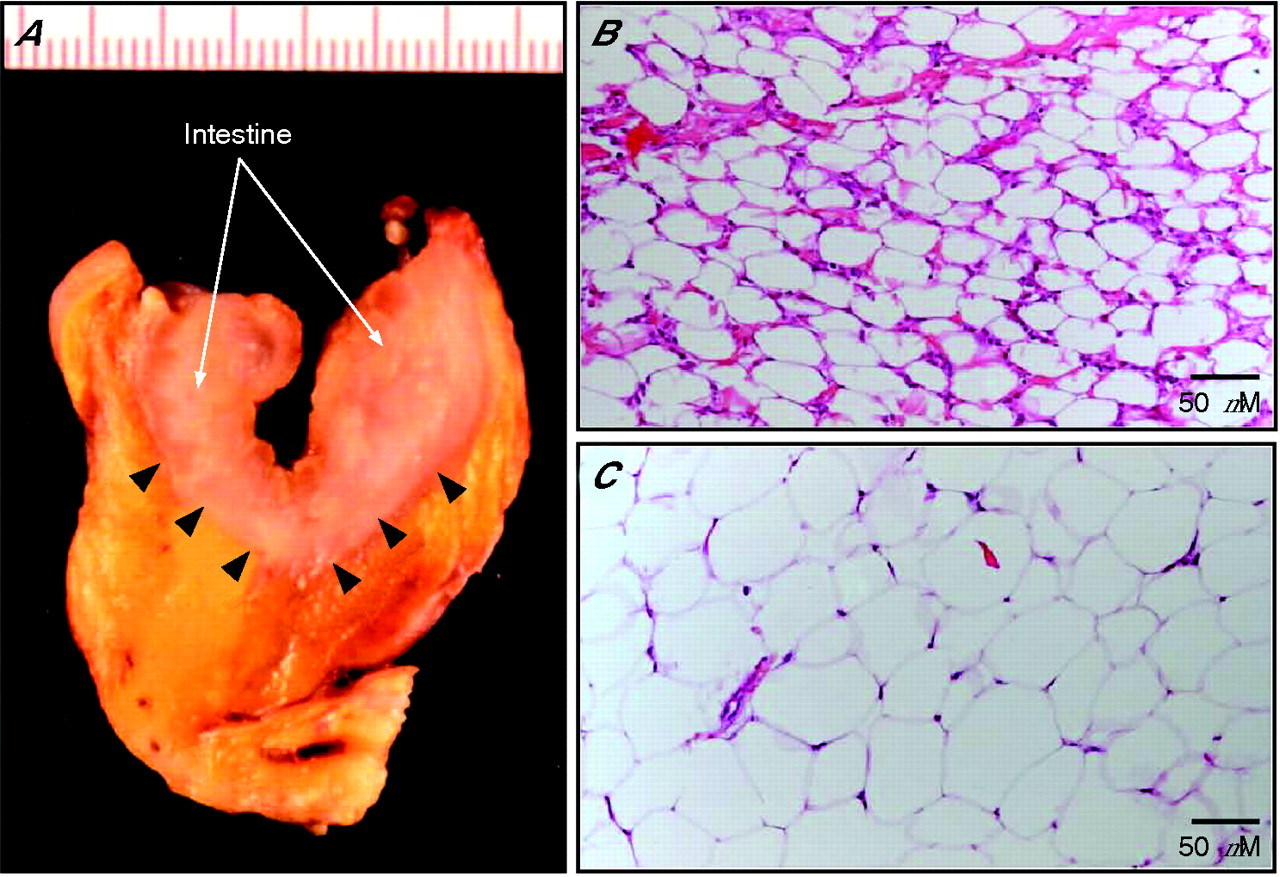
The macroscopical and histological hallmarks of “creeping fat” in Crohn’s disease. (A) Surgical specimen showing white adipose tissue that partially covers the intestinal circumference in association with loss of the bowel-mesentery angle (arrowheads). Histological examination with hematein eosin safran (HES) staining shows more (approximately 150 v 80 cells/slide in a healthy control) and smaller (50 v 80 μm in a healthy control) adipocytes in Crohn’s disease (B) compared to healthy conditions (C).
The measurement of fat distribution has become an important and challenging issue in the field of obesity. Numerous techniques such as anthropometric indices, ultrasonography, computed tomography, magnetic resonance imaging and dual x ray absorptiometry have been developed to assess visceral fat, which seems to be the fat most strongly associated with metabolic disorders.38–,40 Using magnetic resonance imaging, we confirmed and quantified in vivo significant intra-abdominal fat accumulation in patients with Crohn’s disease. Interestingly, fat accumulation was identified at the onset of disease but was not affected by its duration or activity.41
Histological analysis revealed abnormalities in the mWAT of patients with Crohn’s disease,34,36 including marked macrophage and T cell infiltrates, fibrosis, perivascular inflammation and thickening of vessels. Furthermore, visceral adipocytes are significantly smaller, resulting in a fourfold increased number of adipocytes throughout the mesentery of patients with Crohn’s disease compared to controls (fig 2B⇑,C). Taken together, these observations indicate that mesenteric obesity is a common and specific feature of Crohn’s disease and may be due to hyperplasia rather than hypertrophy of the mesenteric adipocytes.
MESENTERIC FAT AND INFLAMMATION IN CROHN’S DISEASE
As regards these circumstantial observations, the role of fat tissue in Crohn’s disease has so far been underestimated. Of more than 6000 papers on Crohn’s disease published in the last 20 years, less than 0.2% of them have mentioned the term “adipose tissue”. In 1999, we showed that mWAT in Crohn’s disease specifically expressed TNFα mRNA, but not the mRNA of several other proinflammatory cytokines. Using immunohistochemical analysis and in situ hybridisation, adipocytes were identified as the main cellular source of TNFα within the mWAT. The absence of detectable TNFα mRNA in the mesentery of controls indicated that this cytokine was not constitutively expressed at this site.41 More recently, the work reported by Yamamoto and colleagues extended our observations by showing increased production and release of adiponectin by adipocytes in hypertrophied mWAT of patients with Crohn’s disease as compared to patients with ulcerative colitis and controls.42 Finally, it must be stressed that in the studies of both Desreumaux and Yamamoto, the increased production of mediators by abdominal fat was certainly underestimated, as the concentrations were expressed per milligram of total protein or number of cDNA per β-actin cDNA molecule.41,42 However, these results did not take into account that the abdominal fat area in Crohn’s disease is composed of a global fourfold increased number of adipocytes as compared to controls.
In addition to the production of TNFα, mWAT is known to produce adiponectin. Interestingly, several studies indicated that adiponectin might have anti-inflammatory properties in vitro and ex vivo.43,44 Although the effect of adiponectin on TNFα expression in adipose tissue has not yet been studied, this adipormone suppressed both TNFα secretion and signalling in macrophage/endothelial cells.45,46 Therefore, as hypothesised by Yamamoto et al,42 the increased secretion of adiponectin in the mesentery of patients with Crohn’s disease could be a TNFα-mediated counter-regulatory mechanism. Abnormal adiponectin concentrations might thus result in unregulated production of TNFα and an increased risk of developing Crohn’s disease lesions such as internal fistula.42 However, this theory must be approached cautiously since the anti-inflammatory roles of adiponectin are still a matter of debate. Indeed, different studies demonstrated a pro-inflammatory effect of this mediator on human placental and adipose tissue explants,47 macrophages and THP-1 cell lines,48 and on the HT-29 colonic epithelial cell line.49 In the latter study, globular adiponectin promoted inflammation through increased expression of IL-8, GM-CSF and MCP-1 and a synergistic effect on IL-8- and GM-CSF-induced IL-1β processing.50 In vivo, preliminary data suggested that adiponectin may play distinct roles in adipose tissue51 and the intestine14 during inflammatory processes. More recently, adiponectin-knockout animals seemed to be protected from dextran sodium sulphate (DSS) and trinitrobenzene sulfonic acid (TNBS)-induced colitis.14 Definitive conclusions about the role of adiponectin in chronic intestinal inflammation cannot be drawn, but we can hypothesise that the local production of this mediator by mWAT in Crohn’s disease might enhance the local release of inflammatory mediators and initiate and/or promote damage to the intestinal mucosa.
EMERGING QUESTIONS ABOUT MESENTERIC ADIPOSE TISSUE IN CROHN’S DISEASE
What are the links between the intestinal/biological characteristics of Crohn’s disease and mesenteric WAT hypertrophy?
Previous studies mostly suggested that hypertrophy of mWAT and its inflammatory changes could participate in the pathogenic process of Crohn’s disease both at the intestinal and systemic levels.
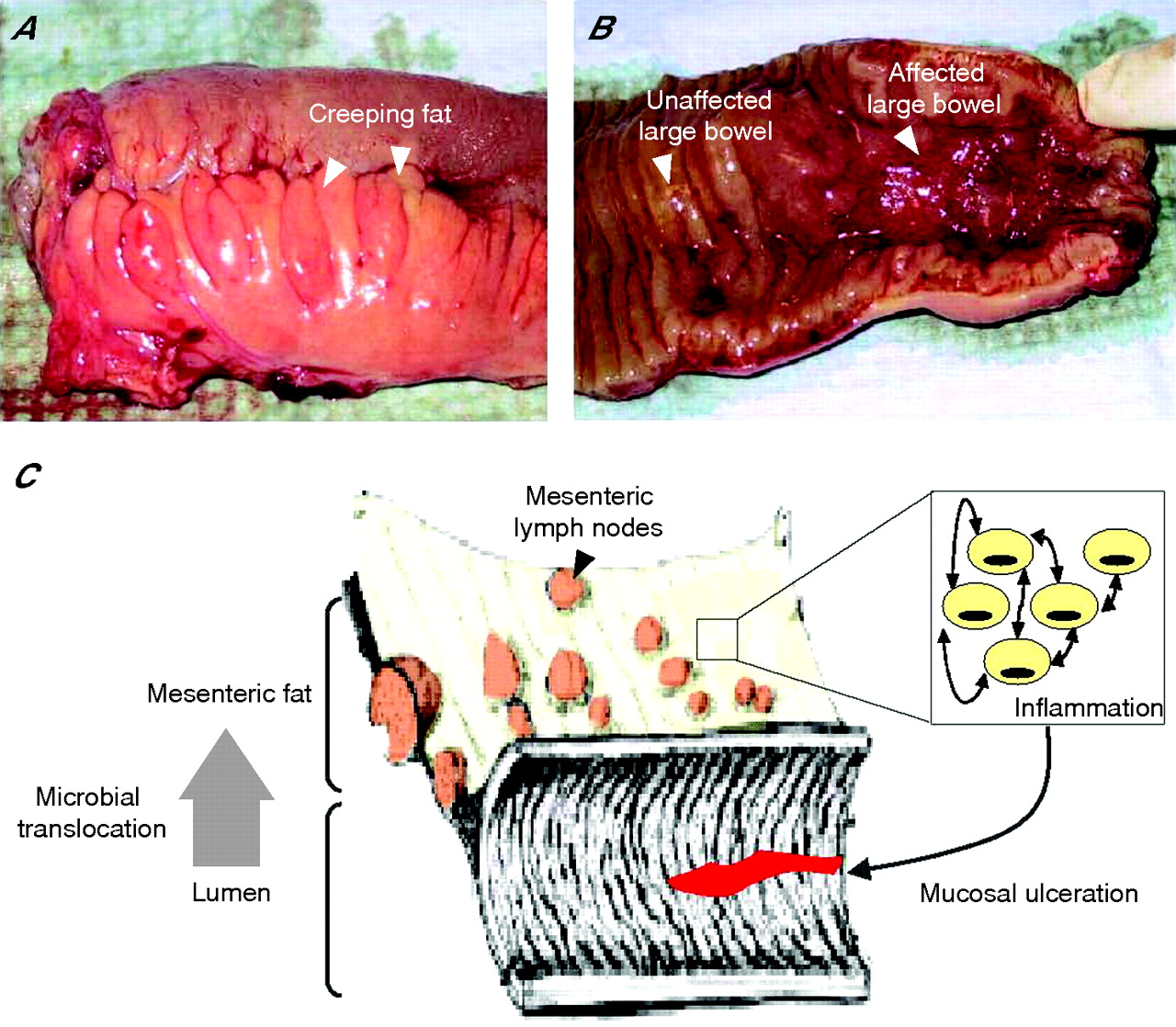
In Crohn’s disease, patchy or linear mucosal ulcerations are located primarily along the mesenteric attachment.37,52 This is in marked contrast to other infectious and inflammatory bowel disorders. In intestinal tuberculosis, ulcerations are usually oriented transversally rather than longitudinally. In infectious conditions mimicking Crohn’s disease (Salmonella typhi, Shigella sp or Yersinia pseudotuberculosis and enterocolitica), the mucosal ulcerations are linear, near Peyer’s patches, and parallel with the long axis of the intestine along the anti-mesenteric border.52 Thus, the axial polarity and the predominance of the ulcerations beneath the attachment of the mesentery are characteristic of Crohn’s disease. A causal link between TNFα synthesis by the mesentery and the particular location of mucosal ulcerations along the mesenteric border may be suggested (fig 3⇓).41 Similarly, fat-wrapping was correlated with transmural inflammation and other connective tissue changes including fibrosis, muscularisation and stricture formation.34 However, further work is now required to formally assess whether or not mesenteric fat hypertrophy is associated with a more aggressive subtype of Crohn’s disease.
{kind=link}
{kind=link}
{kind=link}
A physiopathological model for fat hypertrophy in Crohn’s disease. In chronic inflammatory diseases, white adipose tissue (WAT) hypertrophy is associated with increased production of numerous adipocytokines, such as TNFα. (A) A resected small bowel affected by Crohn’s disease with fat-wrapping (arrows), commonly known as “creeping fat”. (B) In Crohn’s disease, the mucosal ulcerations predominate beneath the attachment of the mesentery. (C) Taking these facts together, we proposed that mesenteric fat may control mucosal ulcerations observed in patients with Crohn’s disease by promoting an inflammatory environment.
C-reactive protein (CRP) is one of the acute phase proteins that increase during systemic inflammation. Unlike ulcerative colitis, active Crohn’s disease is commonly associated with a significant CRP increase.53 Interestingly, Yamamoto et al reported an inverse correlation between adiponectin concentrations in hypertrophied mesenteric tissue and serum CRP levels in patients with Crohn’s disease.44 Unfortunately, no data were available in their study regarding systemic adiponectin concentrations and CRP production within the mesenteric adipose tissue. In coronary atherosclerosis, an inverse association between adiponectin and CRP levels has been observed in both plasma and adipose tissue.50,51 The as yet unexplained difference in CRP production between Crohn’s disease and ulcerative colitis needs to be further investigated but might be explained by the specific fat accumulation associated with Crohn’s disease. Primarily using cultures of human adipocytes and biopsy specimens of mesenteric and subcutaneous adipose tissues taken from IBD patients and controls, we found important expression of CRP mRNA and protein by adipocytes, and a 80- and 1450-fold increase in CRP concentrations in hypertrophied WAT of patients with Crohn’s disease compared to patients with ulcerative colitis and controls, respectively, suggesting that mWAT may be responsible, at least in part, for the elevated CRP plasma levels observed in patients with Crohn’s disease.54 This hypothesis is reinforced by recent data published by Colombel et al reporting a significant correlation between serum CRP levels and increased mesenteric fat density assessed by computed tomography enterography in patients with Crohn’s disease.55
Leptin might also be implicated in the pathophysiology of Crohn’s disease56 through stimulation of CRP production.57 Indeed, physiological concentrations of leptin stimulated the hepatic expression of human CRP.57 In parallel, CRP was capable of inhibiting the functions of leptin by direct binding,57 indicating a potential regulatory feedback loop.
What explains fat accumulation in the mesentery of patients with Crohn’s disease, as opposed to other sites?
Crohn’s disease shares with the HIV-associated adipose redistribution syndrome (HARS) the peculiarity of selective expansion of intra-abdominal adipose tissue while other sites such as the limbs, buttocks, and face are depleted.58 HARS develops gradually after several months of HIV infection, both in untreated patients and in those taking protease inhibitors and nucleoside reverse transcriptase inhibitors.59 Pond suggested that changes in adipose tissue distribution in both cases may implicate preferential interactions between the immune system and perinodal adipocytes.60 Briefly, the polyunsaturated fatty acids derived from perinodal adipocytes might activate adjacent lymph node lymphoid cells. In turn, prolonged and frequent stimulation of immune cells might lead to selective enlargement of lymph node-containing fat depots. Since Crohn’s disease selectively affects the intestinal lymphoid tissue, selective growth of perinodal adipose tissue would be limited to the mesentery. The striking correlation observed in Crohn’s disease between fat-wrapping, lymphoid aggregates and transmural inflammation may support Pond’s hypothesis.34
A role for growth hormone has also been suspected in mesenteric fat accumulation in Crohn’s disease. This hormone can modulate adiposity since growth hormone deficiency is associated with increased central adiposity.61 Consistently, reduced growth hormone levels have been reported in patients with Crohn’s disease.62 Furthermore, Katznelson and colleagues63 demonstrated in Crohn’s disease an inverse correlation between serum growth hormone concentration and intra-abdominal fat accumulation.63 Whether reduction of intra-abdominal fat explains the therapeutic efficacy of growth hormone in Crohn’s disease is not known.64
Mesenteric fat accumulation in Crohn’s disease may develop as the long-term consequence of chronic intestinal inflammation and its subsequent overproduction of growth factors and anti-apoptotic family proteins such as M-CSF,65 insulin-like growth factor-I (IGF-I),66 Bcl-x(L) and Bax proteins.67 Paracrine effects of M-CSF secreted from macrophages within the mesenteric adipose tissue may also participate in WAT hyperplasia in Crohn’s disease. However, since fat accumulation in the mesentery is an early event in the course of Crohn’s disease, not correlated with duration and intensity of intestinal lesions, this hypothesis remains unlikely.
The peroxisome proliferator-activated receptor γ (PPARγ) is a crucial regulator of adipocyte proliferation and differentiation. Given that PPARγ is over expressed in mesenteric adipocytes in patients with Crohn’s disease compared to controls,68 PPARγ stimulation in mesenteric tissues may lead to an increased number of small adipocytes,69 as this dysregulation of PPARγ expression was observed specifically in mesenteric tissues and not the subcutaneous WAT of patients with Crohn’s disease and controls. PPARγ might thus link Crohn’s disease to WAT hyperplasia. However, the underlying mechanisms leading to PPARγ activation in WAT remain poorly investigated in IBD. PPARγ expression is classically downregulated by factors such as fasting and insulin-deficient diabetes,70 while it is positively regulated by obesity and a diet rich in fatty acids.71 These factors are probably not involved in the upregulation of PPARγ expression in Crohn’s disease, as intra-abdominal fat accumulation is not associated with changes in subcutaneous WAT. Local activation of PPARγ is more likely responsible for mesenteric fat hypertrophy in IBD patients. Besides dietary factors, PPARγ expression is modulated by several bacterial stimuli.72 Given the epithelial barrier defects in Crohn’s disease leading to an increased intestinal permeability and bacterial translocation, the intestinal flora may directly regulate the mass of mWAT. Indeed, we observed that mesenteric adipocytes were colonised by luminal bacteria and that 3T3-L1 cells produced lipogenic mediators such as PPARγ.28 Therefore, exposure of mesenteric adipocytes to intestinal bacteria may contribute to mesenteric WAT hypertrophy through increased PPARγ expression. In addition, the gut microbiota may indirectly lead to local activation of PPARγ, as conventionalisation of adult germ-free mice with a normal microbiota rapidly produces a 60% increase in body fat content despite reduced food intake.73 Mechanisms involve the promotion of monosaccharide absorption from the gut lumen and also the selective suppression of fasting-induced adipocyte factor, a member of the angiopoietin-like family of proteins physiologically expressed in epithelial cells. If intestinal flora are an important environmental factor that affect energy harvest from the diet and energy storage in the host, possible impaired expression of the fasting-induced adipocyte factor in Crohn’s disease will need further attention, a condition where several studies have reported abnormalities in the composition of luminal flora.
CONCLUSION AND PERSPECTIVES
In conclusion, the mesenteric fat can no longer be considered a simple bystander in Crohn’s disease, as it may contribute to the increased CRP production previously reported in metabolic disorders. Furthermore, mesenteric fat could also influence the gut barrier function by promoting the innate immune response to the gut flora. However, the origin of mesenteric fat hypertrophy in Crohn’s disease is still unknown. Lifestyle changes, which have been shown to modulate fat distribution, could be involved in the development of fat hypertrophy, as well as a more general interplay between environmental and genetic factors. Notably, genes involved in the control of fat distribution, such as the recently described Gpc4 and Thx15, could be considered as potential candidate genes.74 Finally, an appraisal of the correlation between mesenteric adipose tissue and intestinal inflammation in obese patients may help towards a better understanding of the pathophysiology of chronic inflammatory disorders such as obesity and Crohn’s disease, which may share some common aetiological pathways.
Acknowledgments
We are grateful for the support of grants from UCB, Sanofi-Aventis, the Institut de Recherche des Maladies de l’Appareil Digestif, the Association Francois Aupetit, the Institut Universitaire de France, the Centre Hospitalier et Universitaire de Lille, the Région Nord-Pas de Calais and the Crohn’s & Colitis Foundation of America.
We are grateful to Professor Karel Geboes for fruitful discussion.
REFERENCES
Footnotes
Competing interests: None declared.