Article Text

Abstract
Omega (ω)-3 polyunsaturated fatty acids (PUFAs) are naturally occurring substances that are well tolerated and have been used extensively for the prevention of cardiovascular disease. More recently, ω-3 PUFAs have been recognised to have anticancer activity. There is also evidence suggesting improved efficacy and/or tolerability of conventional cancer chemotherapy when administered with ω-3 PUFAs. The purpose of this review is to (i) describe the mechanisms by which ω-3 PUFAs are thought to have antineoplastic activity, (ii) review published preclinical and clinical studies that support anti-colorectal cancer activity and (iii) summarise current clinical trials investigating the potential therapeutic role(s) of ω-3 PUFAs at different stages of colorectal carcinogenesis, from adenoma (polyp) prevention to treatment of established malignant disease and prevention of cancer recurrence.
- Colorectal neoplasms
- neoplasm metastasis
- fatty acids, Omega-3
- chemotherapy
- cyclooxygenase-2
- colorectal cancer
- cyclooxygenase-2
- fatty acid supplementation
Statistics from Altmetric.com
- Colorectal neoplasms
- neoplasm metastasis
- fatty acids, Omega-3
- chemotherapy
- cyclooxygenase-2
- colorectal cancer
- cyclooxygenase-2
- fatty acid supplementation
Significance of this study
What is already known about this subject?
The two main omega (ω)-3 polyunsaturated fatty acids (PUFAs) found naturally in fish are eicosapentaenoic acid (EPA) and docosahexaenoic acid.
There is strong evidence that ω-3 PUFAs have anti-colorectal cancer (CRC) activity from preclinical studies.
Epidemiological evidence links dietary ω-3 PUFA intake and reduced CRC risk.
Treatment with ω-3 PUFAs is associated with a reduction in measures of mucosal epithelial cell proliferation, a putative biomarker of future CRC risk.
What are the new findings?
EPA has chemopreventative efficacy in patients with familial adenomatous polyposis.
ω-3 PUFAs may improve the efficacy and tolerability of cancer chemotherapy drugs.
Adjuvant treatment with docosahexaenoic acid may improve breast cancer survival and/or reduce cancer-related cachexia.
The mechanisms of action of ω-3 PUFAs probably include reduction of prostaglandin E2 synthesis and/or synthesis of anti-inflammatory resolvins.
How might it impact on clinical practice in the foreseeable future?
An excellent safety and tolerability profile, combined with broader health benefits, make ω-3 PUFAs strong candidate cancer chemoprevention agents.
A role for EPA in ‘sporadic’ colorectal adenoma prevention is currently being evaluated in addition to a potential role in familial adenomatous polyposis patients after colectomy.
A phase II trial is currently evaluating the safety and efficacy of EPA in patients with CRC liver metastases.
ω-3 PUFAs may find a role for ‘tertiary’ prevention of CRC recurrence or as adjunctive chemotherapy for established CRC.
Introduction
There is growing epidemiological, experimental and clinical evidence that omega (ω)-3 polyunsaturated fatty acids (PUFAs) have anti-colorectal cancer (CRC) activity. ω-3 PUFAs may feasibly play a role in several stages of CRC management, from the primary CRC prevention, through to ‘tertiary’ prevention after treatment of CRC and advanced metastatic disease.
In this article, we summarise current knowledge of the mechanistic basis by which ω-3 PUFAs are thought to attenuate CRC, before reviewing the experimental data supporting ω-3 PUFA supplementation for prevention and treatment of CRC. Finally, we review recent clinical data, which now support anti-CRC activity in humans, and summarise current clinical trials of the potential therapeutic roles of ω-3 PUFAs at different stages of colorectal carcinogenesis.
Polyunsaturated fatty acids
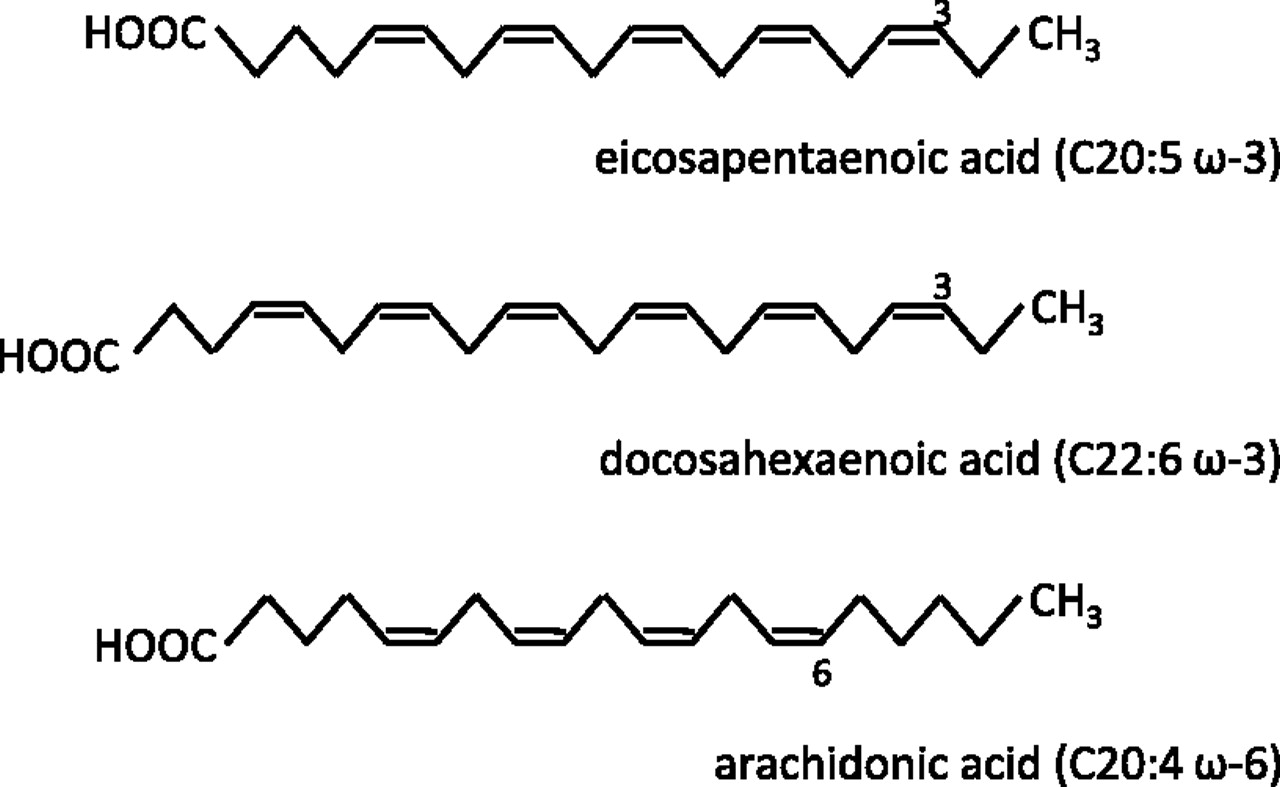
Fatty acids are carbon chains with a methyl group at one end and a carboxyl group at the other. Saturated fatty acids contain only carbon–carbon single bonds, whereas unsaturated fatty acids contain one (monounsaturated) or more (polyunsaturated) carbon–carbon double bonds. Dietary PUFAs of interest are the ω-3 and ω-6 PUFAs, so named by the position of the first double bond at the third and sixth carbon from the methyl (ω) end respectively (figure 1).

Structure and nomenclature of polyunsaturated fatty acids (PUFAs). Cx:y ω-z refers to the chemical structure where x is the number of carbon atoms, y is the number of carbon–carbon double bonds and z is the position of the first carbon–carbon double bond away from the methyl (ω) end of the hydrocarbon chain. Note that all double bonds are in the cis configuration. Small quantities of conjugated ω-3 PUFAs (which contain alternating single and double carbon–carbon bonds) also exist naturally.
‘Essential’ fatty acids are those which are required for biological processes, but which humans are unable to synthesise and must therefore obtain from dietary sources. The parent ω-3 PUFA α-linolenic acid and parent ω-6 PUFA linoleic acid are both found in vegetable oils. Humans can easily metabolise linoleic acid to form the ω-6 PUFA arachidonic acid (AA). However, endogenous production of the ω-3 PUFAs eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) from α-linolenic acid by humans is so small as to be insignificant.1 Therefore, the main ω-3 PUFAs EPA and DHA are also considered ‘essential’ and are obtained predominantly from cold water, oily fish such as mackerel and salmon.
PUFAs are biologically important, with roles in phospholipid membrane structure and function, as well as cellular signalling and lipid metabolism. PUFAs can be liberated from phospholipid membranes by the phospholipase A2 family of enzymes and are metabolised by three main pathways: (i) the cyclo-oxygenase (COX) pathway, (ii) the lipoxygenase (LOX) pathway and (iii) the cytochrome P450 monoxygenase pathway. Metabolites derived from the ω-6 PUFA AA, such as prostaglandin (PG) E2, are typically proinflammatory and have been linked with initiation and progression of colorectal carcinogenesis, whereas those derived from ω-3 PUFAs (eg, PGE3) are less proinflammatory, and may even have anticancer properties. Excellent reviews are already available on the metabolism of ω-3 PUFAs,2 the implications of inhibition of AA metabolism on cell proliferation3 and the effects of ω-6 and ω-3 PUFA metabolites on colorectal carcinogenesis.4
In addition to their potential role in the treatment and prevention of CRC, ω-3 PUFAs are believed to have a number of other health benefits. ω-3 PUFAs (usually a mix of EPA and DHA as fish oil) are widely available in a ‘nutraceutical’ formulation. Evidence is strongest for cardiovascular benefits of the ω-3 PUFAs.5 There is some evidence of benefit in patients who have had a myocardial infarction5 6 but also uncertainty about whether ω-3 PUFAs have anti- or pro-arrhythmogenic properties.5 6 ω-3 PUFAs have also been variously implicated in treatment of inflammatory bowel disease, rheumatoid arthritis, age-related maculopathy and neurodegenerative diseases.7
ω-3 PUFAs
are ‘essential’ nutrients
are found in large quantities in oily fish
are also widely available as ‘over-the-counter’ nutraceuticals
have broad health benefits, including cardiovascular prophylaxis
may have anticancer properties.
Mechanisms of the antineoplastic activity of ω-3 PUFAs
Current knowledge of the antineoplastic activity of ω-3 PUFAs has been comprehensively reviewed elsewhere.2 4 8 A brief summary concentrating on more recent findings is provided here. The three main antineoplastic activities of ω-3 PUFAs that have been proposed are (i) modulation of COX activity; (ii) alteration of membrane dynamics and cell surface receptor function and (iii) increased cellular oxidative stress. Recently described novel anti-inflammatory lipid mediators derived from EPA and DHA, including resolvins, protectins and maresins, represent a fourth potential antineoplastic mechanism of action of ω-3 PUFAs.
Modulation of COX activity
COX-2-dependent synthesis of PGs in colorectal mucosa, in particular PGE2, is believed to play a critical role in the early stages of colorectal carcinogenesis.9 EPA can act as an alternative substrate for COX-2, instead of AA, leading to a reduction in formation of pro-tumorigenic ‘2-series’ PGs (eg, PGE2) in favour of ‘three-series’ PGs (eg, PGE3) in several cell types including CRC cells (figure 2).2 10 11 PGE3 has anti-tumorigenic activity against human lung cancer cells in vitro11 and inhibits pro-tumorigenic PGE2-EP4 receptor signalling in human CRC cells.10 Recently, a ‘PGE2 to PGE3 switch’ has been demonstrated in colorectal mucosa of rats treated with fish oil.12 However, reduction of PGE2 synthesis and/or generation of PGE3 following EPA treatment remains to be demonstrated in human CRC tissue.

{kind=link}
{kind=link}
The effect of eicosapentaenoic acid (EPA) without or with concurrent aspirin treatment on cyclo-oxygenase (COX) activity. (A) When arachidonic acid (AA), derived mainly from dietary ω-6 polyunsaturated fatty acid (PUFA) linoleic acid in ‘westernised’ diets, is the main substrate for COX-1 and COX-2, the predominant prostaglandin (PG) species in colorectal tissue is PGE2. COX-2 is the predominant COX isoform in dysplastic and malignant colorectal tissue.9 (B) EPA can act as an alternative substrate for both COX enzymes. It effectively inhibits COX-1 activity but modulates the enzymatic activity of COX-2 so that PGH3 is the major product.2 PGH3 is then converted to other three-series PGs such as PGE3 by downstream PG synthases. Although the KM for EPA is similar to AA, the enzymatic turnover for EPA is approximately threefold less than that of AA so that the overall outcome is reduction in COX-2-dependent PGE2 synthesis and simultaneous appearance of smaller quantities of PGE3.2 (C) Aspirin irreversibly acetylates COX-1 and COX-2. This leads to effective inhibition of COX-1 activity. However, acetylated COX-2 can metabolise EPA producing 18R-hydroxyeicosapentaenoic acid (HEPE) and 18S-HEPE instead of PGH3. In turn, HEPEs can be metabolised by 5-lipoxygenase (5-LOX) generating E-series resolvins, in combination with a reduction in tissue PGE2 levels.28 29 D-series resolvins are generated via docosahexaenoic acid metabolism by lipoxygenase-dependent pathways.
It is known that DHA also binds the substrate channel of COX-2 and inhibits COX-2 activity,13 although the biochemistry of DHA metabolism by the COX isoforms and the subsequent production of downstream lipid mediators is poorly understood compared with EPA.2
Alteration of membrane dynamics and cell surface receptor function
There is some evidence that the incorporation of ω-3 PUFAs into cell phospholipid membranes alters the fluidity, structure and/or function of lipid rafts or calveolae.14 These are sphingolipid- and cholesterol-rich microdomains that float freely in the cell membrane. The localisation of cell surface receptors, such as epidermal growth factor receptor (EGFR),15 in lipid rafts is believed to be crucial for downstream receptor signalling, controlling proliferation and apoptosis.16 17
Increased oxidative stress
ω-3 PUFAs may have an antineoplastic effect through alteration in the cellular redox state and increased oxidative stress. PUFAs are highly peroxidisable, which generates reactive oxygen species (ROS) such as the superoxide radical. Many tumour cells display altered cellular pathways for the handling of ROS, including depletion of the major intracellular antioxidant, glutathione. A subsequent elevation of intracellular ROS levels by ω-3 PUFAs has been hypothesised to induce cancer cell apoptosis.18 A potential beneficial interaction between ω-3 PUFAs and dietary fibre leading to induction of colonocyte apoptosis has been elegantly studied by Chapkin and colleagues.19–21 DHA has been shown to potentiate oxidative stress and colonocyte apoptosis induced by the fermentation product short-chain fatty acid butyrate via both intrinsic and extrinsic apoptosis pathways.22–24
The relative contributions of, and interactions between, these three activities to the anticancer properties of ω-3 PUFAs are unclear. For example, ω-3 PUFAs might alter EGFR function by changing receptor behaviour in lipid rafts but EPA might also decrease trans-activation of EGFR by reduction in PGE2 synthesis.25 Activation of peroxisome proliferator-activated receptors by ω-3 PUFAs is also recognised but it is not known whether this occurs directly or via changes in COX metabolism, thereby altering levels of PGD2 or 15-deoxy PGJ2.26
Novel anti-inflammatory lipid mediators
In the presence of aspirin, which irreversibly acetylates the COX enzyme, EPA drives COX-2-dependent production of resolvin (Rv) E1 (5S,12R,18R-trihydroxyeicosapentaenoic acid) via metabolism of 18R-hydroxyeicosapentaenoic acid by 5-LOX (figure 2).27 18R-RvE1 has been detected in plasma of healthy volunteers in ng/ml quantities after aspirin and EPA ingestion.28 More recently, synthesis of the 18S enantiomer of RvE1 has been demonstrated after EPA and aspirin treatment in healthy volunteers.29 The precursors of E-series resolvins may also be produced independently of COX by direct cytochrome P450 metabolism of EPA.30
Metabolism of DHA can produce D-series resolvins via a LOX-dependent pathway to produce 17S-resolvins, or via acetylated-COX-2 leading to 17R-resolvin synthesis.31 DHA can also be metabolised by leucocyte-mediated pathways to produce 17S-docosatrienes termed protectins31 or by macrophage-mediated pathways to produce 14-LOX-derived products termed maresins.32 These newly described families of EPA- and DHA-derived lipid mediators all share anti-inflammatory and inflammation resolution activity in animal models of acute inflammation.32 33
Cell signalling via these novel lipid mediator families is best characterised for RvE1. Both 18R and 18S enantiomers of RvE1 are ligands for ChemR23 and BLT1 G protein-coupled receptors.29 RvE1 has been shown to induce expression of intestinal alkaline phosphatase in human CRC cells in a ChemR23-dependent manner and abrogate chemically induced colitis in mice.34 It is currently not known whether ω-3 PUFA-derived resolvins exhibit antineoplastic activity. However, it is known that ChemR23-dependent RvE1 signalling inhibits nuclear factor κB activation, which is a critical regulator of early-stage colorectal carcinogenesis.35
Although ω-3 PUFAs have recognised immunomodulatory activity, including alteration of T-cell activation and cytokine production,36 the contribution of any effect on host antitumour immune surveillance to the anticancer activity of ω-3 PUFAs has received relatively little attention, to date. In vitro studies have suggested that both EPA and DHA may suppress angiogenesis.37 38 Whether negative regulation of stromal-epithelial cell signalling and/or the angiogenic response contributes to the antineoplastic activity of ω-3 PUFAs in vivo requires further investigation.
ω-3 PUFAs for the prevention of CRC
Epidemiological observations
A link between dietary ω-3:ω-6 PUFA balance and CRC risk originated from epidemiological studies that observed reduced rates of CRC in Greenland and the Far East compared with Western populations.39 While the results of epidemiological studies have been variable, they have tended, in general, to report a small reduction in CRC risk with increasing dietary fish intake, a view supported by the second expert report into food, nutrition and the prevention of cancer by the World Cancer Research Fund and the American Institute for Cancer Research in 2007.40
Interpretation of epidemiological studies has been hampered by heterogeneity in study design. The use of food questionnaires to record dietary intake is subjective, and does not always discriminate between oily fish such as sardines (high in ω-3 PUFAs) and lean fish such as cod (lower ω-3 PUFA content). Moreover, studies do not always discriminate between processed (smoked or salted) and non-processed fish, which may confound observational data on CRC risk.40 A meta-analysis of 19 prospective cohort studies was limited by study heterogeneity and wide variability in both frequency of fish intake and the reporting of outcome measures (eg, grams/day, portions/week, ω-3 PUFA g/day).41 Meta-analysis of CRC incidence data could be performed on only 14 of 19 studies. It demonstrated a small but significant 12% relative risk (RR) reduction in CRC incidence between the highest (1–7 portions or 210–590 g fish per week) and lowest (0–1 portions or 0–100 g fish per week) fish consumption groups. The pooled RR reduction rose to 22% in those studies where the difference in fish consumption between the highest and lowest categories was at least seven portions of fish a month.
Since this meta-analysis, two large observational studies have demonstrated significant inverse relationships between ω-3 PUFA intake/levels and risk of colorectal neoplasia. The first, a case–control study of 1872 patients (929 cases of distal CRC and 943 controls), estimated ω-3 and ω-6 PUFA intake from detailed one-to-one dietary interviews.42 The study showed a significant dose-dependent reduction in CRC risk for total ω-3 PUFA intake (OR=0.61 for the highest vs lowest quartile), as well as for EPA and DHA intake individually. In the second study, serum PUFA levels were measured in 861 patients undergoing endoscopy.43 Three hundred and sixty-three patients were diagnosed with a colorectal adenoma(s) and 498 polyp-free patients served as controls. When comparing serum ω-3 PUFA levels (as a percentage of overall PUFA levels), there was a significant reduction in colorectal adenoma risk (OR=0.67) between the first tertile (<1.8% ω-3 PUFA) and the third tertile (>2.3% ω-3 PUFA). By contrast, there was a significant increase in colorectal adenoma risk (OR=1.68) between the first tertile (<58.9%) and third tertile (>62.8%) of serum ω-6 PUFA content.
Animal studies
The differential effect of ω-3 and ω-6 PUFAs has been demonstrated in a number of animal models of early-stage colorectal carcinogenesis relevant to prevention of CRC. These studies are summarised in table 1. Injection of the carcinogens azoxymethane or 1,2-dimethylhydrazine drives development of colorectal aberrant crypt foci (ACF) after 6–8 weeks, before development of colorectal adenomas and adenocarcinomas from 8 months onwards. These lesions have all been widely used as end points for comparison of the effects of ω-3 and ω-6 PUFA supplementation on ‘sporadic’ colorectal carcinogenesis. Alternatively, the effect of ω-3 and ω-6 PUFAs on adenoma number in the ApcMin/+ mouse model of familial adenomatous polyposis (FAP) has been investigated.
In vivo pre-clinical studies of ω-3 PUFA supplementation for the prevention of CRC
Studies of rodents fed an ω-3 PUFA-supplemented diet versus an equivalent ω-6 PUFA-supplemented diet or low-fat control diet controls (table 1) have consistently reported a 20–50% reduction in chemically induced tumour incidence,19 45 47 48 50 52 53 55 56 64 together with a 30–70% reduction in tumour multiplicity, in both carcinogen and ApcMin/+ mouse studies.44 45 50 53 58 59 61 64 Studies using the number of ACFs as the primary end point (table 1) have reported an effect of similar magnitude with ω-3 PUFA supplementation.54–56 59 Reddy and Sugie demonstrated that ω-3 PUFA supplementation reduced chemically induced colonic tumour incidence when given in either the pre- or postinitiation phase compared with ω-6 PUFA supplementation.46 When given in both phases, tumour incidence was further reduced to half that seen in low-fat control animals. Further studies have demonstrated that these effects are directly related to ω-3 PUFA supplementation rather than simply a reduction in ω-6 PUFA intake.53 65 66 While most in vivo studies have compared a mixture of EPA and DHA, attention is drawn to the few studies either directly comparing EPA and DHA,60 or using EPA45 61 64 or DHA49 51 58 as single agents. In general, similar results have been demonstrated with each of the two main ω-3 PUFAs. The effect of fish oil administration on azoxymethane-induced ACF multiplicity in rats has also been tested in combination with the fibre fermentation product butyrate. Crim and colleagues have demonstrated that butyrate decreases ACF number in rats administered fish oil, but not corn oil (rich in ω-6 PUFAs).20
One recent study demonstrated that dietary supplementation with a large amount (6%) of DHA exacerbated colonic inflammation and dysplasia in a mouse model of infection-induced colitis-associated cancer.57 The dose–response relationship between ω-3 PUFA intake and intestinal tumorigenesis in animal models of colitis-associated cancer requires further investigation.
Analysis of mucosal PUFA content has consistently demonstrated incorporation of ω-3 PUFA, at the expense of AA content, in rodents supplemented with ω-3 PUFA compared with controls,45 49 50 together with a reduction in mucosal PGE2,12 45 60–62 reduction in mucosal cell proliferation47 50 54 and an increase in mucosal cell apoptosis 12 19 20 54 62 (table 1).
ω-3 PUFAs in the treatment of CRC
In vitro studies
Many in vitro studies have explored the antineoplastic activity of ω-3 PUFAs against human CRC cells, with both EPA and DHA treatment being associated with reduced cellular proliferation67–74 and increased apoptosis.68 69 72 75 It remains unclear from the few studies comparing EPA and DHA whether there is any significant difference in anti-proliferative and/or pro-apoptotic activity. Calviello et al reported a more pronounced reduction in cell number with EPA than with DHA,71 whereas Chen and Istfan have reported a lower cell number with DHA than with EPA, and that only DHA induced apoptosis.72 Both ω-3 PUFAs have been shown to reduce COX-2 expression and PGE2 production.10 69 71 76 ω-3 PUFAs also reduce cell growth of COX-negative CRC cells, suggesting that the anti-CRC activity of ω 3-PUFA occurs via both COX-dependent and independent mechanisms.67 Increased membrane fluidity73 and lipid peroxidation,73 74 as well as reduced vascular endothelial cell growth factor, β-catenin, peroxisome proliferator-activated receptor γ (PPARγ), BCL-2, matrix metalloproteinase levels and reduced extracellular signal-related kinase-1/2 signalling, have all been demonstrated in human CRC cell lines treated with ω-3 PUFAs.17 69 71 72 75 77
Animal studies
The effect of ω-3 and ω-6 PUFA supplementation on the growth of human CRC cell lines grown as xenograft tumours in immunocompromised mice has been studied widely (table 2). There has been a consistent 40–60% reduction in xenograft size in rodents supplemented with ω-3 PUFAs compared with controls.67 71 81 Similar findings have been reported for studies of rodent CRC cell allograft tumours (table 2).65 78 79 82 84 Similar to data from chemoprevention models, the ω-3 PUFA content of tumours increased, at the expense of AA, in animals supplemented with ω-3 PUFA,67 79 80 82 together with a reduction in expression of COX-2,65 71 reduction in tissue PGE2 levels71 and a reduction in tumour vascularity.71 Conjugated EPA (figure 1) has been shown to have antitumour activity against DLD-1 human CRC cell tumours in nude mice, which was associated with increased lipid peroxidation.83
In vivo pre-clinical studies of ω-3 PUFA supplementation for the treatment of CRC
Models of CRC metastasis
Fewer preclinical studies have investigated the effect of ω-3 PUFA supplementation on the development of metastatic disease. Two studies have modelled liver metastasis by injection of CRC cells into the portal vein or superior mesenteric vein, and a third by injection into the spleen. Iwamoto et al demonstrated a 40% reduction in number and 44% reduction in size of liver metastases in rats supplemented with EPA (9.5% w/w as the ethyl ester) compared with controls. This was associated with a reduction in the tumour cell proliferation index (PI), but no change in the apoptosis index (AI), and downregulation of vascular cell adhesion molecule 1 (VCAM-1). Rats fed a diet high in ω-6 PUFA showed a threefold increase in number and 1.5-fold increase in the size of metastases.85 A more recent study of an EPA/DHA mixture (1.96% fish oil; EPA:DHA ratio 3:2) demonstrated a 70% reduction in incidence and 50% reduction in size of hepatic and extra-hepatic metastases, as well as a 30% reduction in tumours expressing VCAM-1.86
By contrast, Griffini et al demonstrated a 10-fold increase in liver metastases in rats supplemented with an EPA/DHA mixture (20% fish oil v/w; EPA:DHA ratio approximately 3:2) compared with 5% soybean controls, and a fourfold increase compared with those animals supplemented with 20% safflower oil (high in ω-6 PUFA).87 This was associated with an increase in liver:body weight ratio and an increase in mitotic tumour cells in the ω-3 PUFA group. These results are difficult to reconcile with the other preclinical data, but the 20% fish oil preparation used in this study is a much higher dose than that used in any other in vivo study, and is far in excess of any clinically attainable dose in humans.
Two further studies have modelled the effect of ω-3 PUFA supplementation on the development of pulmonary metastases, by measuring lung colonisation after injection of CRC cells into the tail vein.78 79 One demonstrated that supplementation of either EPA or DHA (0.1 ml aliquots daily of 98% pure EPA or DHA ethyl ester) was associated with significantly fewer lung colonisations at 12 days compared with controls (54% and 58% fewer colonies, respectively).79 The other study found that while supplementation with high-fat safflower oil (24.7% by weight for 30 days preinoculation) caused a fivefold increase in the number of pulmonary colonies, there was no difference in low (5%) or high (24.7%) ω-3 PUFA intake on the number of metastases compared with controls.78
Pharmacology, safety and tolerability of ω-3 PUFAs in humans
Bioavailability of ω-3 PUFA preparations
The bioavailability of ω-3 PUFAs is highly variable and is dependent on the form of the ω-3 PUFA, the mode of administration and the timing of administration—for example, with food. Dietary PUFAs are absorbed primarily in the small intestine. ω-3 PUFAs in the free fatty acid (FFA) form are absorbed more efficiently than those in the triglyceride or ethyl ester form.88 89 Lawson and Hughes demonstrated >95% absorption of an oral dose of EPA and DHA (as measured by plasma PUFA content) when given to healthy volunteers in the FFA form, whereas absorption in the triglyceride and ethyl ester forms was approximately 65% and 21%, respectively.89 These differences are believed to be due to the ability of enterocytes to directly absorb ω-3 PUFAs in the FFA form, whereas absorption of those in the ethyl ester and triglyceride forms requires prior hydrolysis by pancreatic lipase.89 Pancreatic lipase hydrolyses fatty acids from the three positions of the triglyceride backbone at different rates.90 Bioavailability is therefore further complicated by variation in the distribution of fatty acids in the sn-1, sn-2 and sn-3 positions in fish oils from marine mammals, marine fish and processed fish oil supplements.91 92 Co-absorption with other dietary fats, absorption as micelles rather than direct absorption through the intestinal wall and the variable action of gastric lipase and gastric absorption can also affect bioavailability. These variables make it difficult to obtain direct comparisons between dose and outcomes in different clinical (and animal) studies.
Duration of ω-3 PUFA supplementation
There is considerable variation in the duration of ω-3 PUFA supplementation in clinical studies, ranging from 4 weeks to 6 months. These are summarised in table 3. Studies have demonstrated that maximal incorporation of orally administered ω-3 PUFA into plasma and colorectal mucosa occurs within 3 weeks.103 104 Hillier et al gave fish oil (3.2 g EPA+2.2 g DHA daily) to 11 patients with active inflammatory bowel disease and took colonic biopsy specimens at 3, 6 and 12 weeks.104 After 3 weeks, mucosal EPA content had increased sevenfold and DHA content had doubled. These levels were maintained but did not increase further throughout the 12 weeks' study period. By contrast, AA content did not fall significantly until after 6 weeks of treatment. Colonic mucosal PGE2 content fell to 45% of baseline values after 3 weeks of fish oil supplementation, and fell only slightly further to 43% of the baseline value at 12 weeks. Similarly, in eight healthy volunteers taking 1.4–4.2 g daily EPA for 12 weeks, a significant, maximal increase in EPA content of plasma, platelets, neutrophils, monocytes and lymphocytes was observed at 2 weeks, whereas AA content fell significantly only by week 12.103
Clinical studies of ω-3 PUFA treatment on colorectal mucosa biomarkers and polyp number
These results support the hypothesis that the increased mucosal EPA and DHA content, rather than the reduction in ω-6 PUFA content, contributes to the reduction in PGE2 levels. Tissue ω-3 PUFA incorporation in humans plateaus after a few weeks. The optimal duration of ω-3 PUFA treatment required to produce maximal antineoplastic effects remains to be established.
Adverse effects
ω-3 PUFA formulations are generally very well tolerated. Table 3 summarises the adverse effects reported in human studies of the effect of ω-3 PUFA supplementation on colorectal mucosal parameters and colorectal polyps. Study withdrawal due to EPA intolerance in a recent phase III trial of EPA-FFA 2 g daily was only 3.4%.102 In a previous phase II trial with the same preparation, the commonest gastrointestinal adverse event, diarrhoea, was associated with the EPA 2 g daily dose of EPA (14.0%), but not the 1 g dose (2%, cf. placebo 7.8%), suggesting dose de-escalation as a potential strategy for maintaining ω-3 PUFA treatment if diarrhoea does occur.64
The excellent safety and tolerability profile of ω-3 PUFA supplementation is supported in large studies of the effects of ω-3 PUFAs in cardiovascular disease. In the GISSI prevention study, 2836 patients were given 0.85 g ω-3 PUFA for 3.5 years.105 The commonest side effects were gastrointestinal disturbance (4.9%) and nausea (1.4%) Only 3.8% of patients discontinued the drug. Gastrointestinal symptoms are typically mild diarrhoea, indigestion, belching or a ‘fishy’ taste.106 107 In a study using a higher dose of 6.9 g EPA+DHA daily for 6 months, gastrointestinal disturbance was reported by 7% of 275 patients, but also by 8% of patients taking placebo (corn-oil) capsules.108
For more detail, the reader is recommended to review concise overviews of the safety and toxicology of EPA and DHA by Kim et al92 and Lien.109
Risk of bleeding
ω-3 PUFAs have antiplatelet activity, thought to be due to a reduction in COX-1-dependent production of thromboxane A2 from AA in platelets, leading to reduced platelet activation and aggregation.110 However, the concern that ω-3 PUFAs may increase bleeding has not been realised clinically, even when co-administered with aspirin. A recent review highlighted that in over 4000 patients taking ω-3 PUFAs undergoing carotid endarterectomy, percutaneous transluminal coronary angioplasty or coronary artery bypass grafting, excess bleeding was virtually non-existent.106 111
Environmental contaminants
While it has been suggested that fish oils may contain potentially toxic levels of heavy metals (eg, mercury), environmental toxins and fat-soluble vitamins, all of which can accumulate in fish, there is no evidence to support this in practice.106 112 The extraction and purification methods used in the production of fish oil supplements result in negligible or undetectable levels of such toxins,106 112 and so fish oil preparations given in daily doses up to 2 g/day are classified by the US Food and Drug Administration (FDA) as ‘generally regarded as safe.’
Translational studies of human colorectal biomarkers and ω-3 PUFA content
Mucosal biomarker studies
The long natural history of colorectal carcinogenesis in humans precludes the use of CRC incidence as a primary end point in clinical intervention studies and a 3–5 year intervention period is generally required for adenoma (polyp) prevention trials. Therefore many investigators have measured the effect of ω-3 PUFA administration on putative mucosal biomarkers of future CRC risk, such as epithelial cell mitosis frequency in microdissected whole crypts or immunohistochemistry for the Ki-67 ‘proliferation’ antigen. The design of such studies and their main findings are summarised in table 3.
We identified eight studies of oral ω-3 PUFA supplementation in patients with previous ‘sporadic’ colorectal adenomas (table 3), in which colorectal mucosal biopsy specimens were obtained at endoscopy before and after ω-3 PUFA supplementation. Two studies used EPA-FFA, whereas the others used a fish oil mixture containing EPA and DHA in varying proportions in either the ethyl ester or triglyceride form. In six of eight studies a 13–70% reduction in mucosal epithelial cell proliferation index was observed compared with the respective placebo group.64 93 94 96 97 100 In those studies comparing different doses of ω-3 PUFA, a dose-dependent reduction in the PI was observed.94 96
By contrast, two studies demonstrated no change in PI following administration of 2.4 g ω-3 PUFA daily for 12 weeks98 or low-dose ω-3 PUFA (400 mg DHA+100 mg EPA/day) for 1–2 years.99 The latter study did demonstrate a 50% increase in AI and increased expression of the pro-apoptotic protein BAX. Although in vivo preclinical work suggests that ω-3 PUFAs may have anti-CRC activity via an increase in apoptosis, AI has been measured in only two other studies, which demonstrated a significant increase in AI after 3–6 months' treatment with EPA 2 g daily.64 100 One further study noted a more modest 16% reduction in PI after 28 days supplementation in healthy volunteers.95 In those studies measuring mucosal PUFA content, significant increases in mucosal DHA and EPA, together with a reduction in mucosal AA, were observed in all but one study (table 3).
Polyp prevention studies
Patients with FAP have a heterozygous germline mutation in the Adenomatous Polyposis Coli (APC) gene. Following somatic loss of the second APC allele, multiple colorectal adenomas develop, and prophylactic colectomy is advised in order to prevent CRC. Patients who undergo total colectomy with ileorectal anastomosis require regular endoscopic surveillance of the remaining rectum, which remains at risk of further polyps. Studies of potential chemopreventative agents in these patients allow polyp size and number to be used as end points over a relatively short (6–12 months) period of time.
A recent phase III randomised, double-blind, placebo-controlled trial investigated treatment with EPA-FFA 2 g daily for 6 months in patients (n=58) with FAP who had previously undergone colectomy and ileorectal anastomosis.102 Rectal polyp multiplicity and size were measured by blinded video-endoscopic assessment of a tattooed area at baseline and at 6 months. There was a 22.4% reduction in polyp number in the EPA group compared with placebo (p=0.01), a similar magnitude of reduction to that seen with the selective COX-2 inhibitor celecoxib.113 In keeping with previous studies, a significant increase in mucosal EPA levels was seen. The demonstration of chemopreventative efficacy of EPA-FFA in patients with FAP has led to funding of a randomised, placebo-controlled trial of EPA-FFA in patients who have had ‘sporadic’ colorectal adenomas removed and who require further colonoscopic surveillance (http://www.eme.ac.uk/funded_projects/).
Only one further polyp prevention study was identified.101 In this small study, five patients who had previously undergone colectomy for FAP or had >30 colorectal polyps removed endoscopically were given 2.2 g DHA+0.6 g EPA daily for 1–2 years.101 No significant change in polyp number was seen. One patient developed proximal CRC, a second patient developed lung cancer and a third patient developed endometrial cancer before termination of the study. The published report does not make it clear whether this study was stopped prematurely and gives no indication of the planned sample size for the trial.
Clinical trials of ω-3 PUFAs in patients with CRC
Single agent treatment
Despite strong in vitro and animal model evidence for direct anti-CRC activity of ω-3 PUFAs, no published studies have yet investigated the antineoplastic effect of ω-3 PUFAs in patients with primary or metastatic CRC. The authors are currently recruiting patients into a phase II trial of the safety and efficacy of oral EPA-FFA 2 g daily in patients awaiting surgery for colorectal cancer liver metastases (CRCLM) (NCT01070355). Primary tissue biomarker end points (tumour cell proliferation, apoptosis and angiogenesis) are supported by mechanistic analyses investigating whether a PGE2-PGE3 switch occurs in CRC tissue. The authors are aware of only one further trial, registered on http://clinicaltrials.org/ (NCT00942292), investigating the effects of a parenteral fish oil preparation on tumour angiogenesis in CRCLM. ω-3 fish oils are also being investigated for the treatment of advanced prostatic cancer (NCT00996749), and in the prevention and treatment of breast cancer (NCT01282580, NCT00627276).
Adjuvant therapy with traditional chemoradiotherapy
DHA has been shown to potentiate the pro-apoptotic properties of 5-fluorouracil against several CRC cell lines, with a reduction in the expression of the antiapoptotic proteins BCL-2 and BCL-XL.114 Moreover, the addition of a fish oil mixture (55% EPA, 45% DHA) to 5-fluorouracil synergistically reduces CRC cell proliferation.115 Similar results have been demonstrated in rodent CRC models.116 These findings complement those of combination fish oil treatment with other chemotherapeutic drugs in vitro and in vivo for other solid tumour and haematological malignancies.117
We identified only one published human study of the anticancer effects of combining ω-3 PUFAs with chemotherapy. A phase II study evaluated addition of 1.8 g DHA daily to an anthracycline-based chemotherapy regimen for metastatic breast cancer. Patients were dichotomised into two groups based on high or low DHA incorporation into plasma phospholipids. The high DHA-incorporation group had a significantly longer time to disease progression (median 8.7 months vs 3.5 months) and overall survival (median 34 months vs 18 months).118
There are a small number of in vitro studies demonstrating that both EPA and DHA potentiate the cytotoxicity of ionising radiation in various cell lines, including CRC cells.119 A retrospective review of 143 patients, who had been prescribed ω-3 fish oil (0.9 g EPA, 1.5 g DHA daily) for 18 weeks following radiotherapy for brain metastases, found reduced radionecrosis (3.5% vs 14.1%) and improved overall survival (median survival 88.8 weeks vs 54.1 weeks) compared with the 262 patients who had not been prescribed fish oil.120 While un-blinded and non-randomised, this study nevertheless demonstrated a clear survival advantage linked to ω-3 PUFA supplementation.
One further study demonstrated improved tolerability of chemoradiation for oesophageal cancer in patients taking ω-3 PUFA supplements, with a reduction in the incidence of grade 2–4 neutropenia, diarrhoea and pharyngitis compared with those not taking supplements.121 There is a clear need for further human studies to evaluate the role of ω-3 PUFA supplementation in improving the efficacy and/or tolerability of chemotherapy and radiotherapy. Two ongoing studies are investigating the combination of fish oil (either enteral or parenteral) and gemcitabine in patients with advanced pancreatic cancer (study identifiers NCT01019382 and JPRN-UMIN000003658). A DHA-paclitaxel compound is also being investigated for the treatment of metastatic prostate (NCT00024414), pancreatic (NCT00024375) and CRC (NCT00024401).
ω-3 PUFAs as nutritional treatment
While the potential anti-CRC benefits of ω-3 PUFAs as either single agent or combination treatment are yet to be established in human studies, the role of ω-3 PUFA supplementation for preventing cancer-related cachexia has been more widely investigated.
There have been two systematic reviews of the effects of ω-3 PUFAs on cancer cachexia and outcome. A Cochrane review of the benefit of EPA for treating cancer cachexia identified 59 potential trials, of which only five were randomised controlled trials (RCTs).122 Patients in these studies generally had pancreatic, gastrointestinal or lung cancers. No trial has studied CRC patients exclusively. The Cochrane review concluded that there were insufficient data to determine whether EPA supplementation was beneficial in the treatment of cancer cachexia, and identified a clear need for a large well conducted RCT. Four of the five trials demonstrated either stabilisation of weight loss, or weight gain, following EPA supplementation, but this was no greater than the weight changes associated with the isocaloric control diet. However, one study did demonstrate a significant dose-dependent association between EPA supplementation and weight gain when taking into consideration the poor tolerance of up to 18 capsules/day by some patients.123 Of the studies reporting survival data, one demonstrated improved survival in the EPA-supplemented group versus placebo (median 390 days vs 165 days),124 but neither of the two studies comparing EPA supplementation with an isocaloric control intervention demonstrated a survival benefit of EPA.123 125
Colomer et al published a systematic review including the above RCTs and an additional 12 prospective observational studies.126 Many of these studies were in patients with advanced pancreatic cancer. They concluded that there was fair evidence (grade B) to recommend ω-3 PUFA supplementation in patients with solid upper gastrointestinal and pancreatic tumours, with the benefits of increased weight, appetite and quality of life, as well as lower postoperative morbidity.
We have identified only one published (uncontrolled) study in patients with CRC. Treatment of 23 patients with advanced CRC awaiting chemotherapy was started with oral supplement drinks containing a total of 2.18 g EPA+0.92 g DHA per day for 4 weeks before and 5 weeks after irinotecan-based chemotherapy (FOLFIRI). There was a significant improvement in weight (mean increase 2.5 kg) during the period of ω-3 PUFA supplementation before chemotherapy, despite a fall in total protein and energy intake in this time, with weight gain persisting after the start of chemotherapy.127
More recently, a RCT demonstrated that patients undergoing oesophagectomy for cancer who received EPA-enriched enteral nutrition for 5 days pre- and 21 days postoperatively conserved fat-free mass compared with patients receiving standard enteral nutrition. Total weight loss was 1.2 kg in the EPA group versus 1.9 kg in the control group (p=0.03). The EPA group also demonstrated an attenuated stress response (measured by serum tumour necrosis factor α, interleukin 8 (interleukin 10 levels) compared with controls, although there was no difference in postoperative complications.128
Animal studies have shown that ω-3 PUFA supplementation impairs the host response to mycobacterial infection.129 130 This raises the question whether the anti-inflammatory and immunoregulatory effects of ω-3 PUFA supplementation might increase susceptibility to infection in immunocompromised patients. This complex topic was comprehensively examined in a review in 2002.131 Subsequent preclinical data have been conflicting.129 130 132–135 Moreover, results from limited human observational studies investigating fatty acid intake and risk of community-acquired pneumonia are inconclusive.136 137 Long-term observational studies of ω-3 PUFA intake have not revealed an increase in infectious adverse events.136 137 Further trials of ω-3 PUFAs for CRC treatment and/or prevention should examine whether ω-3 PUFAs alter host susceptibility to infection, particularly in immunocompromised patients with cancer who may require surgery.
Is there a difference in anti-CRC efficacy between EPA and DHA?
It is clear from the preclinical ‘prevention’ studies that combination EPA and DHA treatment has anti-CRC activity. There is only one ‘head-to-head’ comparison, which demonstrated that EPA had greater efficacy than DHA.60 However, the two preclinical ‘treatment’ studies that have been published did not show any difference between EPA and DHA.71 79 As far as we are aware, there is no published report comparing EPA and DHA in patients with, or at-risk for, CRC. It is also unclear whether an EPA/DHA mixture would give additional benefits over and above equivalent doses of either ω-3 PUFA alone.
In addition, a note of caution should be made of the two studies that demonstrated negative effects of high-dose fish oil in a model of CRCLM (12% EPA/8% DHA mix)87 and a model of infection-induced, colitis-associated cancer (6% DHA),57 in conflict with the majority of ω-3 PUFA studies. The mechanistic basis of a possible negative effect of high-dose ω-3 PUFAs is not clear and warrants further investigation with particular emphasis on the safe maximal dose of either EPA or DHA as single agents, or in combination with other chemotherapeutic agents.
Any comparison between the efficacy of EPA and DHA may also be confounded by interconversion between these two ω-3 PUFAs in vivo. EPA to DHA conversion can occur, via docosapentaenoic acid, in a two-stage process involving elongase and desaturase activity (with docosapentaenoic acid–EPA conversion being the limiting step). However, limited evidence from clinical trials suggests that there is no significant EPA–DHA conversion in human colorectal mucosa.100 102 By contrast, administration of DHA in humans has been shown to result in an increase in EPA content in plasma phospholipids.138 This may be as high as 12% during chronic dosing with DHA and is thought to occur via β-oxidation of the fatty acid.138 The possibility that DHA treatment could result in an eventual increase in EPA content of normal and/or neoplastic colorectal mucosa requires investigation.
Conclusion
ω-3 PUFAs are likely to have multifaceted roles in both prevention and treatment of CRC. The excellent tolerability and safety profile of ω-3 PUFAs combined with other health benefits, particularly cardiovascular, make ω-3 PUFAs an attractive candidate for prevention and treatment of CRC (and other cancers).
There is a wealth of evidence to support the antineoplastic effects of ω-3 PUFAs, with both in vivo models and human biomarker studies demonstrating a clear reduction in mucosal proliferation and an increase in mucosal apoptosis, as well as mucosal ω-3 PUFA incorporation at the expense of ω-6 PUFA. The role of ω-3 PUFAs in prevention of CRC is further supported by the recent demonstration of chemopreventative efficacy of EPA-FFA in patients with FAP.102
Strong in vivo evidence also supports a role for ω-3 PUFAs in the treatment of CRC, with studies consistently demonstrating tumour ω-3 PUFA incorporation and a reduction in tumour size. Phase II and phase III evaluations of ω-3 PUFAs are justified to establish their safety, tolerability and antineoplastic activity in patients with established CRC. Ongoing studies are investigating the effect of ω-3 PUFAs on tumour growth and vascularity in CRCLM. These studies should eventually inform the design of large, multicentre phase III evaluation of ω-3 PUFAs in patients with CRC, both as neoadjuvant/adjuvant treatment in patients undergoing surgery with curative intent and in the management of incurable disease. ω-3 PUFA treatment should be investigated, as a single agent and in combination with other chemotherapeutic agents, for treatment of CRC. Evaluation of combination therapy with ω-3 PUFAs is already underway in other cancers, including breast and pancreatic cancer.
A third potential role of ω-3 PUFAs as nutritional treatment in patients with CRC has also been identified, with reported improvements in cancer-related cachexia and perioperative outcomes in other cancers.
Improved understanding of the mechanisms of anti-CRC activity of EPA and DHA will lead to identification and validation of predictive therapeutic response biomarkers for use in future clinical trials and help to determine the best anticancer agents to use in combination with ω-3 PUFAs for prevention and/or treatment of CRC.
References
Supplementary materials
Gut's education editor Mairi McLean talks to Mark Hull about the paper in the Gut podcast
Footnotes
Search strategy and selection criteria: Data for this review were identified by searches of Medline, PubMed and relevant articles using the MESH search terms ‘fish oils’, ‘fatty acids’, ‘colorectal neoplasms’ and ‘liver neoplasms’. Only papers published in English, or including an English abstract, between 1975 and 2010 were included. Ongoing clinical trials were identified by searches of publicly available Clinical Trials Registries (including http://ClinicalTrials.gov/, Current Controlled Trials, WHO International Clinical Trials Registry Platform, National Cancer Institute and NIHR Clinical Research Network Portfolio) and pharmaceutical company registries (including the International Federation of Pharmaceutical Manufacturers and Associations Clinical Trials Portal, Pfizer Clinical Trials and GlaxoSmithKline Clinical Trials Register).
Competing interests MAH has received an unrestricted scientific grant from, and performed paid consultancy work for, SLA Pharma AG, which produces a formulation of eicosapentaenoic acid.
Provenance and peer review Commissioned; externally peer reviewed.