Article Text

Abstract
Functional gastrointestinal disorders (FGIDs) are characterized by chronic complaints arising from disorganized brain-gut interactions leading to dysmotility and hypersensitivity. The two most prevalent FGIDs, affecting up to 16–26% of worldwide population, are functional dyspepsia and irritable bowel syndrome. Their etiopathogenic mechanisms remain unclear, however, recent observations reveal low-grade mucosal inflammation and immune activation, in association with impaired epithelial barrier function and aberrant neuronal sensitivity. These findings come to challenge the traditional view of FGIDs as pure functional disorders, and relate the origin to a tangible organic substrate. The mucosal inflammatory infiltrate is dominated by mast cells, eosinophils and intraepithelial lymphocytes in the intestine of FGIDs. It is well established that mast cell activation can generate epithelial and neuro-muscular dysfunction and promote visceral hypersensitivity and altered motility patterns in FGIDs, postoperative ileus, food allergy and inflammatory bowel disease. This review will discuss the role of mucosal mast cells in the gastrointestinal tract with a specific focus on recent advances in disease mechanisms and clinical management in irritable bowel syndrome and functional dyspepsia.
- FUNCTIONAL BOWEL DISORDER
- FUNCTIONAL DYSPEPSIA
- IRRITABLE BOWEL SYNDROME
- NERVE - GUT INTERACTIONS
- MAST CELLS
Statistics from Altmetric.com
- FUNCTIONAL BOWEL DISORDER
- FUNCTIONAL DYSPEPSIA
- IRRITABLE BOWEL SYNDROME
- NERVE - GUT INTERACTIONS
- MAST CELLS
Key messages
Mast cells play a central pathophysiological role in IBS and possibly in functional dyspepsia, although not well defined.
Increased mast cell activation is a common finding in the mucosa of patients with functional GI disorders.
There is a need to implement standardised methods to count mast cells in the GI mucosa and to establish reference ranges of normality.
Evaluation of spontaneous and stimulated mast cell function and activity on GI samples is recommended when available.
More studies are required to fully understand the implication of mast cells in the origin of clinical manifestations of these disorders.
Treatment with mast cell stabilisers offers a reasonably safe and promising option for the management of those patients with IBS non-responding to conventional approaches, though future studies are warranted to evaluate efficacy and indications.
Introduction
Functional GI disorders (FGIDs) are characterised by chronic complaints arising from disorganised brain–gut interactions leading to dysmotility and hypersensitivity. FGIDs diagnosis is made by symptom-based approach using the corresponding Rome criteria. Functional dyspepsia (FD) and IBS are the two most prevalent FGIDs, affecting up to 16–26% of worldwide population.1 ,2 However, despite these figures, their etiopathogenic mechanisms remain unclear, accounting for the lack of diagnostic biomarkers and the paucity of therapeutic options providing satisfactory long-standing clinical remission.3
FGIDs are associated with a high prevalence of psychiatric comorbidities, chronic fatigue and chronic somatic and visceral pain disorders, rendering substantial social, humanistic and direct and indirect healthcare costs.4 Recent observations revealing the presence of low-grade mucosal inflammation and immune activation, in association with impaired epithelial barrier function5 ,6 and aberrant neuronal sensitivity, come to challenge the traditional view of FGIDs as pure functional disorders, and relate the origin to a tangible organic substrate that stimulates the search for innovative diagnostic and therapeutic approaches. Mast cells (MCs), eosinophils and intraepithelial lymphocytes dominate the inflammatory infiltrate in the intestine of FGIDs. MC activation can generate epithelial and neuromuscular dysfunction and promote visceral hypersensitivity and altered motility patterns in FGIDs,7–9 postoperative ileus, food allergy and IBD.10 This review will discuss the role of mucosal MCs in the GI tract with a specific focus on recent advances in disease mechanisms and management in IBS and FD.
The origin, phenotypes and function of GI MCs
MCs are long-lived granulated cells derived from bone marrow myeloid-cell progenitors (CD34+), under the influence of stem cell factor and interleukin (IL)-4, cytokines that also regulate the development of MCs subtypes.11 MC progenitor cells (CD34+, CD13+, c-kit+, FcεRI−) circulate in low numbers in the blood and migrate to locate in close proximity to blood and lymphatic vessels, glands, smooth muscle and nerves. In the tissue, they remain as a homeostatic pool or they complete their differentiation process into mature MCs, as a direct consequence of genetic background, and inflammatory or bacterial-derived molecules released in the local microenvironment, including IL-3, IL-4, IL-9, IL-10, IL-33, CXCL12, transforming growth factor-β, nerve growth factor (NGF),and stem cell factor.12 Intestinal homing of MCs progenitor cells depends mostly on the binding of α4β7 integrin with their corresponding adhesion molecules such as cell adhesion molecule-1 or vascular cell adhesion molecule-1 on the endothelium, although the CXC chemokine receptor 2, expressed on MC progenitors, has been also implicated.13 Mature MCs are particularly abundant in body barriers, ready for optimal interaction with the local environment. In the GI tract, MCs comprise 1–5% of mononuclear cells in the lamina propria and the submucosa, and are also found intraepithelial and deep in the muscle and serosal layers. Based on the anatomical location, human MCs are classified into mucosal MCs and connective tissue MCs, while depending on protease content, MCs are divided in two large subsets: MCT, containing tryptase but little or no chymase, and MCTC, containing tryptase, chymase and carboxypeptidase.12 ,13 MCC, which express chymase but little or no tryptase, also have been described, but they appear to be infrequent.12 ,13 MCT prevail in the intestinal and pulmonary mucosa, near T cells, whereas MCTC are found in the skin and lymph nodes, in addition to the lung and the gut submucosa.11 In the human small intestine, MCT represent ∼98% of all MCs in the mucosa and ∼13% of MCs in the submucosa are MCT.12 Recently, a new phenotype of MCs expressing tryptase and carboxypeptidase A3, but not chymase, has been described in the airway epithelium in asthmatic subjects and in oesophageal samples of patients with eosinophilic esophagitis.14 Heterogeneity of MCs also includes differential content in heparin, cytokines and the receptor for the complement C5a, and the trans-differentiation between subtypes.12 ,13 Therefore, location and granule content will determine the nature of mediators released to the extracellular milieu, accounting for modulation of specific functions in the GI tract.11
MCs have been viewed, for the most part, as effectors of allergy and anaphylaxis and are best known for their association with pathological conditions such as asthma. However, the advent of MC lines, mouse strains deficient in MCs and the reconstitution of these strains with bone marrow-derived MCs has greatly facilitated the characterisation of various aspects of MC function in vivo and their involvement in several disease states by interacting with a variety of other cells implicated in physiological and immunological responses. In the GI tract, MCs regulate vascular and epithelial permeability, ion secretion, angiogenesis, peristalsis, fibrosis and tissue repair, innate and adaptive immunity, bacterial defence, chemotaxis and nociception.11 Hence, uncontrolled or dysregulated MC activation may interfere with gut homeostasis and generate tissue dysfunction and promote inflammation in diverse GI diseases such as food allergy, IBD, postoperative ileus, autoimmune disorders, cancer and FGIDs.11 However, at the same time, MCs are indispensable for controlling a wide range of pathogenic infections, and for modulation innate and adaptive immune responses.15 Indeed, MCs can be intentionally activated to enhance protective host responses, including the production of high-affinity antibodies and immunological memory, raising the possibility of incorporating MC activators in vaccine formulations to harness the inherent adjuvant activity of MC activation.15
Regulation and activation of MCs
The classical and most effective stimulus for MC activation is cross-linking of cell surface-bound IgE to its high-affinity receptor (FcεRI) by allergen in sensitised individuals.16 This results in a sequence of phosphorylation cascades and activation motifs that leads to intracellular calcium flux, activation of certain transcription factors such as AP-1 (c-FOS, v-Jun), MITF and STAT-5, and MC degranulation and cytokine production.17 MCs also express receptors for IgG (FcγRI), immunoglobulin free-light chains (IgLC), other Ig-associated receptors, complement fractions and toll-like receptors. Moreover, MCs can be activated by neurotransmitters, neuropeptides, growth factors and hormones (table 1), accounting for MC versatility. Upon activation, MCs release newly synthesised (lipid mediators and cytokines) and stored (histamine, heparin, proteases) bioactive substances contained in cytoplasmic lipid bodies and granules (figure 1). Secretion is achieved by IgE-mediated rapid release of all granule contents by fusion of granules and extrusion (anaphylactic degranulation) or by partial or total granule emptying without inter-granule fusion (piecemeal degranulation).18 Neuropeptides, cytokines and microbial products induce piecemeal degranulation as frequently seen in diverse diseases, including IBD, IBS and FD.19
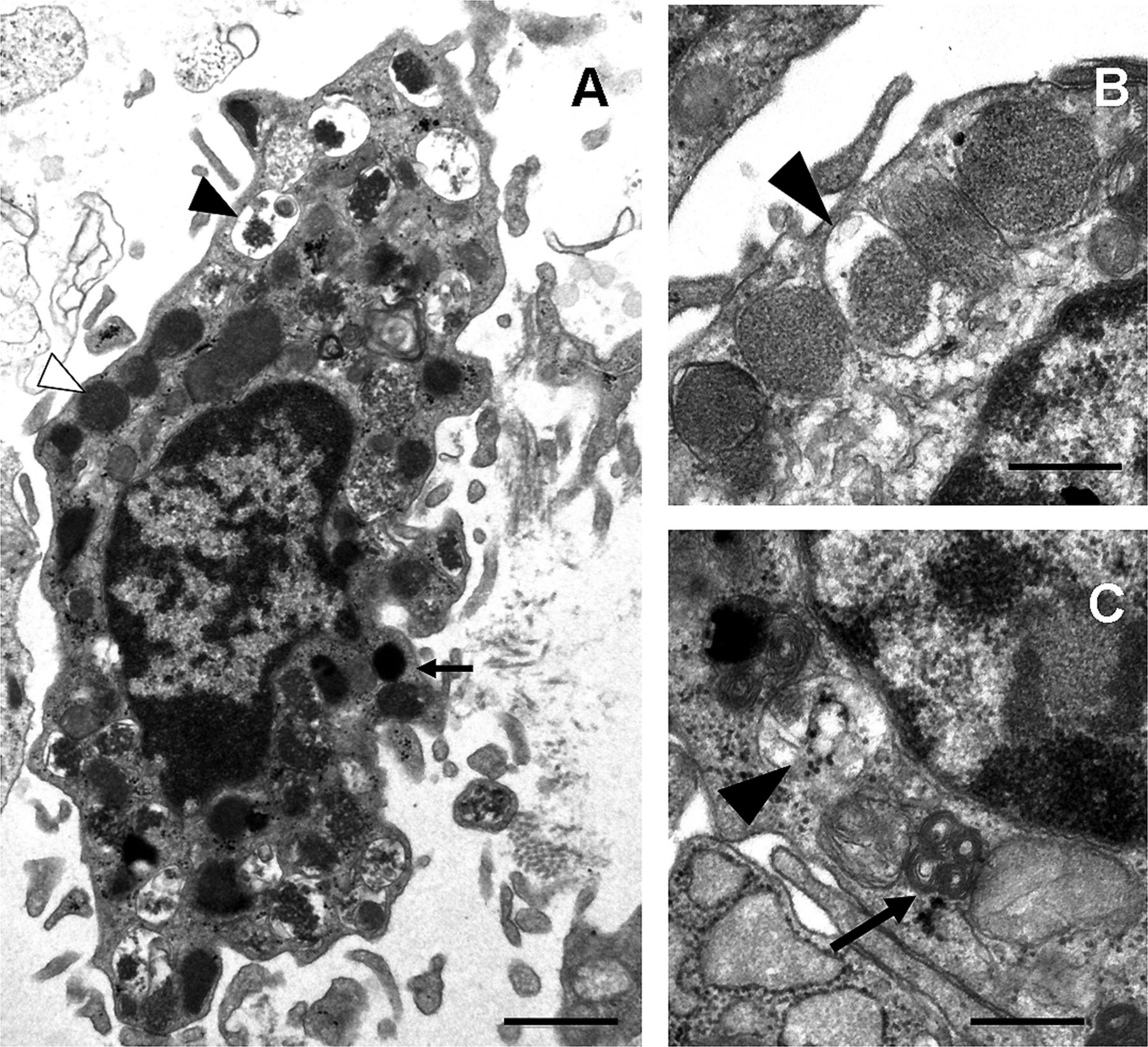
Triggers of mast cell activation

Ultrastructure of human mucosal mast cell. (A) Ultrastructure of an activated mast cell in the intestinal mucosa, with irregular plasma membrane and numerous lipid bodies (arrow) and cytoplasmic granules, displaying piecemeal degranulation. Intact (white arrowhead) and degranulated (black arrowhead) granules are identified. (B and C) High-magnification micrographs of cytoplasmic granules from a mucosal mast cell. Different granule patterns are observed, with crystalloid structure (B) and scrolls (arrow, C). Enlarged empty and partially empty granule containers (black arrowhead) are typical of piecemeal degranulation. Bars: 1 μm (A) and 0.5 μm (B and C).
Factors and mechanisms underlying MC activation in the gut
Food antigens as trigger for MC activation
The majority of patients with FGIDs consider their symptoms to be related to meals. For example, >60% of patients with IBS report the onset or worsening of symptoms after meals, within 15 min in 28% and within 3 h in 93% of these patients.20 ,21 Classically, in food allergy, MCs are activated by food antigen-dependent cross-linking of antigen-specific IgE to FcεRI. Although some patients with IBS have a higher incidence of atopy,22 food allergy has not been convincingly associated to FGID pathogenesis. Of note, adverse reactions to food, including some types of food intolerance, may occur through IgG-mediated sensitisation of MCs, but the role for these IgG-mediated immune reactions remains to be established.20 ,22 When candidate food antigens are directly applied to the duodenal mucosa of IBS patients with suspected food intolerance through an endoscope, it caused immediate epithelial breaks, increased intervillous spaces and increased intraepithelial lymphocyte numbers in the intestinal mucosa,23 and an individualised exclusion diet improved symptoms in 74% of patients at 1-year follow-up. The underlying mechanism and the potential role for MCs requires further study. On the other hand, the response to food is also partly regulated by neuroendocrine factors including peripheral serotonergic responses.24 Although MCs can secrete and synthesise serotonin from tryptophan and serotonin is a chemotactic molecule for MCs,25 and some adverse reactions to diet in FGIDs involve foods containing serotonin, including cheese, meat, soya beans, cereals, nuts and vegetables,26 the role of MCs in such responses, if any, is mostly ignored. Finally, spice intake correlates directly with the likelihood of developing IBS in females.27 Spicy foods contain capsaicin, the natural ligand of transient receptor potential vanilloid 1 (TRPV1) receptors on nociceptive afferent C-fibres. The increased density of sensory fibres expressing TRPV1 receptors reported in patients with FGIDs and visceral hypersensitivity,28 the genetic polymorphism of TRPV1 gene in FD,29 the potential TRPV1 sensitisation in patients with IBS,30 the close proximity of MCs to TRPV1 expressing sensory nerve fibres and the ability of capsaicin to modulate MCs31 all suggest that transmission of pain signals, including those generated by spicy foods, may be enhanced in FGIDs. In contrast, desensitisation of afferent terminals by a high capsaicin diet seems also plausible, as one study reported beneficial effects on abdominal bloating and pain in response to the ingestion of encapsulated red pepper for 6 weeks in IBS.32
The role of infections
Post-infectious (PI)-FGIDs represent common entities in daily clinical practice. Infectious gastroenteritis is associated with an increased risk for FD and IBS; however, the mechanisms leading to chronicity remain unknown.33 MCs are potential regulatory linkers between innate and adaptive immunity and have been demonstrated to play critical roles in host defence, participating in effective immune responses to a number of bacterial, parasitic, viral and fungal pathogen products.15 Antibody titres against bacterial flagellin are increased in patients with IBS and are even higher in PI-IBS.34 Recently, increased mucosal Ig production and upregulation of germline transcripts and Ig genes have been identified in diarrhoea predominant IBS (IBS-D) together with increased proximity between MC and plasma cell, suggesting MC activation by Ig.35 Whether FGID individuals may become sensitised to food and microbial antigens during an acute infection and subsequently develop antibodies that will activate MCs upon antigen exposure remains to be established.
The role of stress
Chronic stress may also lead to MC activation. In preclinical studies, several types of stresses and stress mediators such as corticotropin releasing hormone (CRF) and related peptides have been shown to modulate ion and water secretion as well as intestinal and colonic paracellular and transcellular permeability, primarily via nerve–MC interactions.36 ,37 Similarly, stress-induced rectal hyperalgesia could be prevented and reversed by administration of an MC stabiliser.38 Other studies have confirmed and extended this paradigm to the human intestine. Santos et al39 showed that a cold stress increased jejunal MC tryptase and histamine release along with intestinal water secretion, and intestinal permeability, with larger responses in women with moderate levels of background stress.40 CRF has been shown to enhance transcellular uptake of macromolecules in human colonic mucosa via CRF-R1 and CRF-R2 receptors located on subepithelial MCs.41 More recently, acute psychological stress (public speech) has been shown to increase small intestinal permeability in humans.42 This effect could be reproduced by peripheral administration of CRF, and blocked by the MC stabiliser disodium cromoglycate (DSCG). Preclinical models showed that chronic stress can induce substance P (SP) release by efferent nerves in the periphery, leading to CRF expression and release by intestinal eosinophils. Eosinophil-derived CRF was then capable of activating MCs, resulting in jejunal epithelial barrier dysfunction.43 SP, NGF and sex steroids also induce the release of vasoactive mediators from MCs, contributing to chloride secretion, barrier dysfunction, hyperalgesia, diarrhoea, inflammation and motility changes.44 ,45
MC infiltration in the GI tract in FGIDs
Since the description by Weston et al46 in 1993 on the infiltration of the terminal ileum by MCs in IBS, numerous studies evaluated MC numbers in the GI mucosa of FGIDs (table 2). It is interesting to note here that the presence of low-grade intestinal inflammation in the gut of these patients also involves an increase in intraepithelial T lymphocytes, and less consistently, enterochromaffin cells, plasma cells, B lymphocytes, neutrophils and other immunocytes.47 ,48
Studies describing mast cell infiltration and activation in FD and IBS and potential correlation with symptoms
MCs have been identified by metachromatic stains such as Giemsa or toluidine blue, but these methods have been replaced by immunohistochemistry (antibodies for c-kit (CD117) or tryptase)49–51 because it is more sensitive and specific. MC counts are comparable with both stains, yet CD117+ cells display a more stable membranous staining, whereas tryptase+ cells display cytoplasmic staining that could be influenced by cell degranulation.52 FGID biopsies contain singly dispersed MCs with no aggregates.52 When elevated MC counts are detected, it may be helpful to exclude systemic mastocytosis by staining for the low-affinity receptor for IL-2 (CD25).52 A reference range for significant increased MC counts is still lacking. This is partly due to the absence of agreement and standardisation on the methodology used to count MCs, to differences in patient and control selection, inter-individual variation, location of the biopsy, the relatively small cohort numbers for the majority of individual studies and to other uncontrolled potential confounding factors (box 1) (see Nasser et al for a detailed review). The great variation in reporting mean mucosal MC numbers in the GI tract makes the interpretation of discriminatory cut-off values very complicated and currently uninterpretable according to some pathologists.53 MC counts have been found to be normal, increased or decreased in IBS (table 2). However, although the numbers vary across studies and segments, the analysis of >1000 IBS biopsies detects a mean, modest 1.2-fold to 2.5-fold increase in MC numbers throughout the entire GI tract.54 ,55 This is also true for cases of chronic undefined diarrhoea, mostly studied in the upper small bowel and left colon, to the point that some pathologists debate the convenience of coining the term mastocytic enterocolitis for this clinical–pathological association.56 A significant finding is that mucosal MC ‘hyperplasia’, when present, is not limited to the lower small intestine57 ,58 and colon59 ,60 but also involves the duodenum,61 the jejunum62 and the rectum.63 While there is discrepancy in IBS, available studies in FD reveal that MC numbers are significantly increased in the antrum and corpus of Helicobacter pylori-negative FD,64 ,65 and in the duodenum of patients with FD (table 2).9 ,61 ,66 ,67 Moreover, increased MCs have been recently reported in the oesophagus of patients suffering from non-cardiac chest pain.68 Even so, it is hard to dismiss the physiological relevance of such ‘modest’ increases because, on the one side, similar incremental changes in leucocyte counts in circulating blood occur in infectious and inflammatory conditions, and on the other side, the magnitude of cell change is enormous if we consider the total mucosal surface of the GI tract.
Conditions that may alter, commonly increase, the number of mast cell counts in the GI tract
Allergic diseases: chronic urticaria, food allergy, atopy, hereditary angioedema.11 ,98
Mastocytosis and mast cell activation syndrome.52
Coeliac disease: increased in initial stages and decreased in later stages.135
Neuroendocrine cancer, lymphoma, epithelial cancers, carcinoid syndrome
H. pylori gastritis, infectious and parasitic enteritis,
IBS,136 lymphocytic colitis.48
Intestinal pseudo-obstruction, diverticulitis.137
Vasculitis, amiloidosis, drugs.
When evaluating MCs in IBS subtypes, some studies show that MC hyperplasia is more common in IBS-D69 ,70 and in non-PI IBS71 than in other subtypes, though in many other studies this is not the case.72–74 In contrast, MCs are increased similarly in gastric biopsies in PI-FD and non-specific FD.75 Moreover, others found MC numbers decreased in the descending colon of diarrhoea and alternating predominant IBS, but not constipation predominant IBS compared with health.51 There is also some indication that MC numbers remain increased compared with both non-PI IBS and controls, 3 years after Shigella infection.73 Although not the scope of this review, an increased number of MCs have been reported in the colorectal mucosa, in the lamina propria and in the submucosa from patients with Crohn's disease and UC.10
The role of gender differences in MC number is unclear. Several lines of evidence indicate that gonadal steroids are involved in gender-related differences in tissue MC infiltration in the colon. This difference in the number of MCs has been described in a variety of tissues from rodents, such as skin, myocardium and rat colon. When specifically analysed, some authors found increased MC counts in the terminal ileum, ascending and descending colon, and rectum of female versus male controls,57 ,60 ,74 with females showing 43% increase in the area occupied by MCs,7 similar to observations in patients with chronic undefined diarrhoea,53 while others do not.51 ,60 ,63 These data raise the hypothesis that gender-dependent differences in immune responses are involved in the observed higher prevalence of IBS in females, in the described gender-related differences in IBS pathophysiology and in the known effects of the menstrual cycle in the modulation of rectal sensitivity.76
Differences in MC numbers in the jejunum, caecum, colon or rectum of IBS are not attributable to age, stress and cortisol levels, anxiety or depression, or duration of the disease.51 ,60 ,62 ,69 Although disputed, it seems that changes in MC counts cannot be easily explained by differences in bowel preparation.7 ,48 The role of diet on MC counts remains to be established. Thus, the diagnostic utility of routine MC stains in GI biopsies remains unclear and requires further investigation.
MC activation in the GI tract in FGIDs
MC activation in the GI tract may be evaluated by (1) morphological analysis, most commonly by checking ultrastructural characteristics of piecemeal or anaphylactic degranulation on transmission electron microscopy (TEM); (2) measuring the spontaneous or stimulated release of mediators in tissue, intestinal fluid and blood, most commonly tryptase and histamine, and less often hexosaminidase, carboxypeptidase A, heparin, chromogranin A, leukotriene E4, prostaglandin D2 and prostaglandin 9α,11βPGF2 and methylhistamine in urine; and (3) the expression of related genes and proteins in the mucosa (figure 2).

Schematic representation of the experimental procedure to assess mucosal mast cell activation. Mast cell activation can be measured in intestinal samples. Luminal content can be obtained by aspiration, before biopsies are collected, and tryptase content can be quantified. Different mucosal biopsies can be processed for histological examination, including mast cell counting after immunohistochemistry (tryptase and/or c-kit staining) and laser microdissection for ulterior gene expression analysis; ultrastructure analysis, to assess the type and degree of degranulation and to identify granule pattern; gene expression analysis of specific mediators synthesised and released by mast cells (tryptase, carboxypeptidase, chymase); quantification of mediators that are spontaneously released from biopsies and/or performing functional studies in vitro (muscle/nervous cells) or in vivo (mice/rats); and electrophysiology experiments in Ussing chambers for identification of mast cell-dependent changes in barrier function. Finally, analysis of the possible association between clinical manifestations and mast cell activation can be performed.
Based on TEM studies, it has been shown that MCs display higher activation rates in the caecum and rectum in IBS-D, and that activation rates increase even more when nerve–MC distance is <2 μm.59 Moreover, MCs located within 5 μm of nerve fibres were 3.1 times more frequent in the descending colon of IBS than in controls, and there was a 150% increase in the number of degranulating MCs.7 Furthermore, the ileal and colonic density of neuronal specific enolase, SP and 5-hydroxytryptamine positively stained nerve fibres increased and appeared in clusters, surrounding an increased number of MCs with no differences between PI and non-post-infection patients with IBS.73 ,77
Supernatants of mucosal biopsies of patients with IBS contain increased concentrations of histamine, serotonin, trypsin, tryptase, prostaglandin E2, other proteases and cytokines.7 ,78–80 Moreover, jejunal luminal tryptase release was five times higher62 and the expression of both tryptase mRNA and protein enhanced in jejunal tissue8 in IBS-D, while serum tryptase remained unaltered. Tryptase protein expression was also higher in both PI FD and non-specific FD gastric biopsies.64
It is interesting that λIgLC+ MCs but not IgE or IgG+ MCs are reduced in the colon of IBS.51 This finding, together with the description of elevated serum concentrations of λ and κ IgFLC in IBS,81 suggests that Ig light chain-mediated MC activation may be associated with IBS.
Taken together, evidence indicates that the activity of MCs rather than an increased number is essential in the pathophysiology of FGIDs, a point that has been recently raised by several experts in the field.
Linking MC infiltration and activation in the GI tract with clinical manifestations in FGIDs
Role of MCs in visceral hypersensitivity and motility changes: motor and neuronal activation and sensitisation
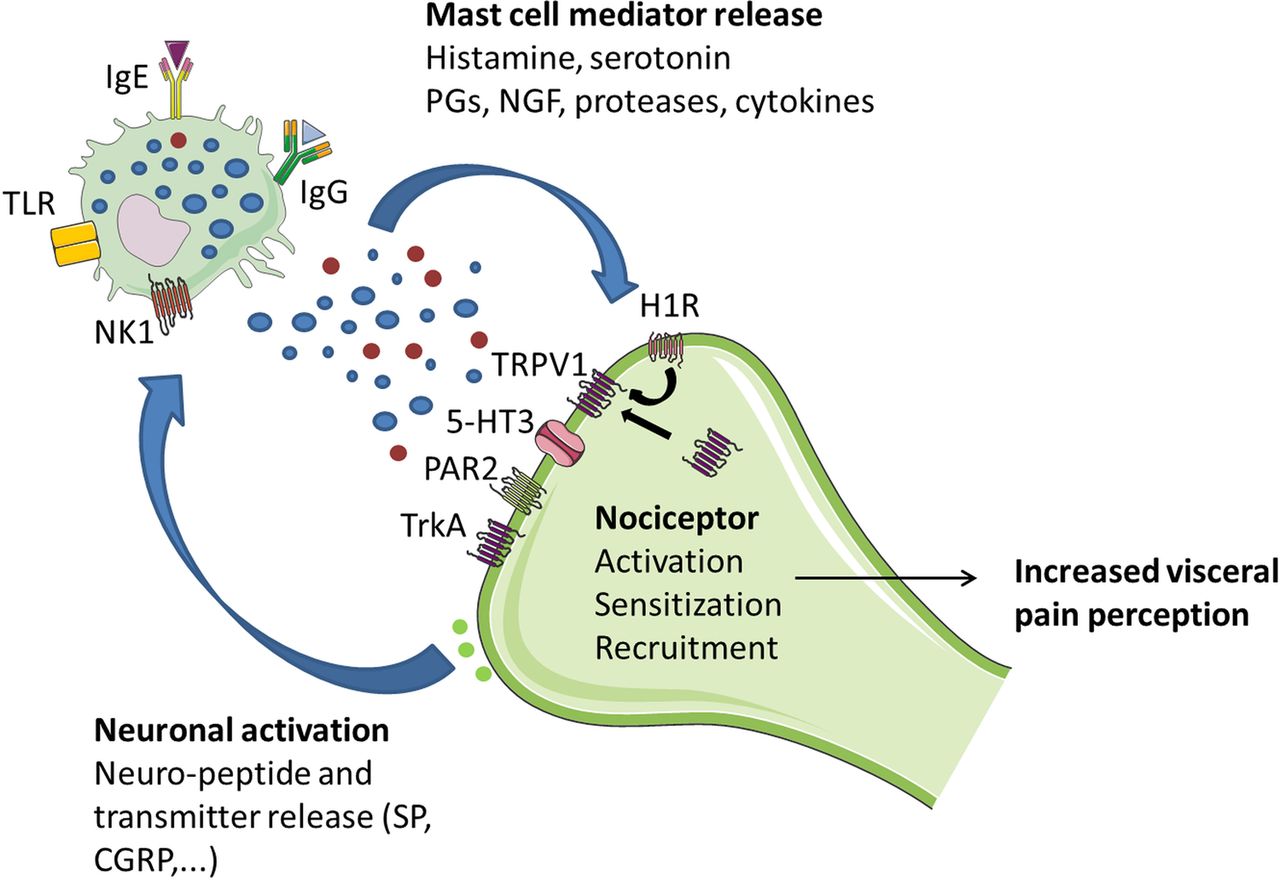
In the human gut, MCs lie in close proximity to GI mucosal sensory nerve fibres containing neuropeptides, including visceral afferents expressing TRPV1 receptors.82 This close spatial association, when coupled with MC activation, has been suggested to be of functional relevance for neuromuscular function and altered pain perception in response to insults such as infections, stress and emotions in FGIDs.47 ,83 Indeed, afferent innervation of enteric MCs can trigger the release of histamine and MC protease II, mediators that act in a paracrine manner to elevate the sensitivity of spinal afferent terminals.84 The use of supernatants obtained from biopsies allows us to study the effect of these mediators on neuronal activation and sensitisation. Injection of IBS-derived supernatants into rat mesenteric arteries evoked a marked increase in afferent nerve discharge, whereas injection of control supernatants had no effect.79 In addition, IBS-dependent excitation of dorsal root ganglia (DRG) was inhibited by histamine H1 receptor blockade and serine protease inactivation,78 underscoring the role of MC mediators in neuronal activation. These findings were confirmed by Buhner et al, who reported that IBS biopsy supernatants, but not those of healthy controls, significantly increased the spike discharge of human submucosal neurons. This effect was inhibited by histamine receptor (H1-H3) antagonists, 5-HT3 receptor antagonist and protease inhibition.80 Moreover, supernatants from hypersensitive patients with IBS caused stronger activation of guinea pig enteric and mouse DRG neurons compared with supernatants of normosensitive patients,85 indicating that neuronal activation responses in vitro correlate with the individual pain threshold pressure values. Others showed that intracolonic infusion of IBS supernatants, but not controls, caused increased nociception in response to colorectal distention in mice, an effect that could be prevented by a serine protease inhibitor and was absent in neurons lacking functional protease-activated receptor-2.79 More recently, Cenac et al86 showed that colonic biopsies from patients with IBS contain increased levels of polyunsaturated fatty acid (PUFA) metabolites, these are endogenous TRPV4 agonists, compared with healthy subjects, and these increases correlated with pain and bloating scores. PUFA metabolites extracted from IBS biopsies or colons of mice with visceral hypersensitivity activated mouse sensory neurons in vitro, by activating TRPV4, an effect that could be prevented by siRNA knockdown of TRPV4.86 Finally, application of supernatants on muscle strips evoked excitatory cholinergic longitudinal muscle contractions of the guinea pig ileum, an effect that was not dependent on serotonin, proteases or histamine but was (partially) mediated by TRPV1, purinergic P2X receptors and prostanoid receptors.87 Ballestra et al speculate that afferent nerve activation may induce myenteric cholinergic depolarisations, leading to altered motor function (figure 3).

Schematic illustration of mast cell–nerve interactions in human gut. MCs and nerves communicate bidirectionally, thereby modulating peristalsis and pain signalling. The release of bioactive, pro-inflammatory, mediators by mast cells results in a variety of neuronal effects including activation, sensitisation and recruitment of nociceptors to the cell membrane, neurogenic inflammation and neural sprouting, ultimately leading to visceral hypersensitivity. On the other hand, neuronal activation triggers the release of neuropeptides and neurotransmitters, thereby further activating mast cells. 5-HT3, 5-hydroxytryptamine receptor 3; CRGP, calcitonin-related gene peptide; H1R, histamine receptor 1; Ig, immunoglobulins; NK1, neurokinin 1 receptor; NGF, neuronal growth factor; PGs, prostaglandins; PAR2, proteinase-activated receptor-2; SP, substance P; TRPV1, transient receptor potential vanilloid 1; TLR, toll-like receptor; TrkA, receptor for nerve growth factor.
Besides increased neuronal activation, supernatant of biopsies from patients with IBS also has the capacity to potentiate sensory nerves. In a recent, elegant study, murine DRG neurons were incubated overnight with supernatants of submucosal colonic biopsies of IBS. Patch clamp recordings the next day revealed that the intrinsic excitability of the colonic nociceptive DRG neurons was increased by IBS-D supernatants. This increased excitability was not observed in DRG neurons lacking PAR-2.88 Finally, incubation of a neuronal cell line or rat primary myenteric neuron cultures with mucosal biopsy supernatants from IBS also induced long-lasting neuroplastic changes as reflected by increased NGF-dependent neuronal sprouting.77
Together, these preclinical data consistently indicate that the mucosa and submucosa of patients with IBS contains increased levels of various MC mediators that have the potential to activate and potentiate intrinsic and afferent neurons, thereby leading to increased visceral pain perception and altered motor function that may cause diarrhoea or constipation as a result of excessive segmental contractile colonic motor activity (table 3). Of note, the use of human supernatants on animal models or isolated neurons may not completely reflect human physiology as MCs and enteric neurons exhibit species specificity in mediator release mechanisms and receptor profile.16 To further assess the functional relevance and specificity of supernatant-mediated activation of nerve endings in the gut, it may be of great interest to perform live imaging of MC-nerve signalling in human preparations or to perform confocal endomicroscopy. The latter has recently been used to identify suspected food intolerance in patients with IBS.23
Effect of mast cell mediators on GI function
Role of MCs in the regulation of intestinal barrier function: secretion and permeability
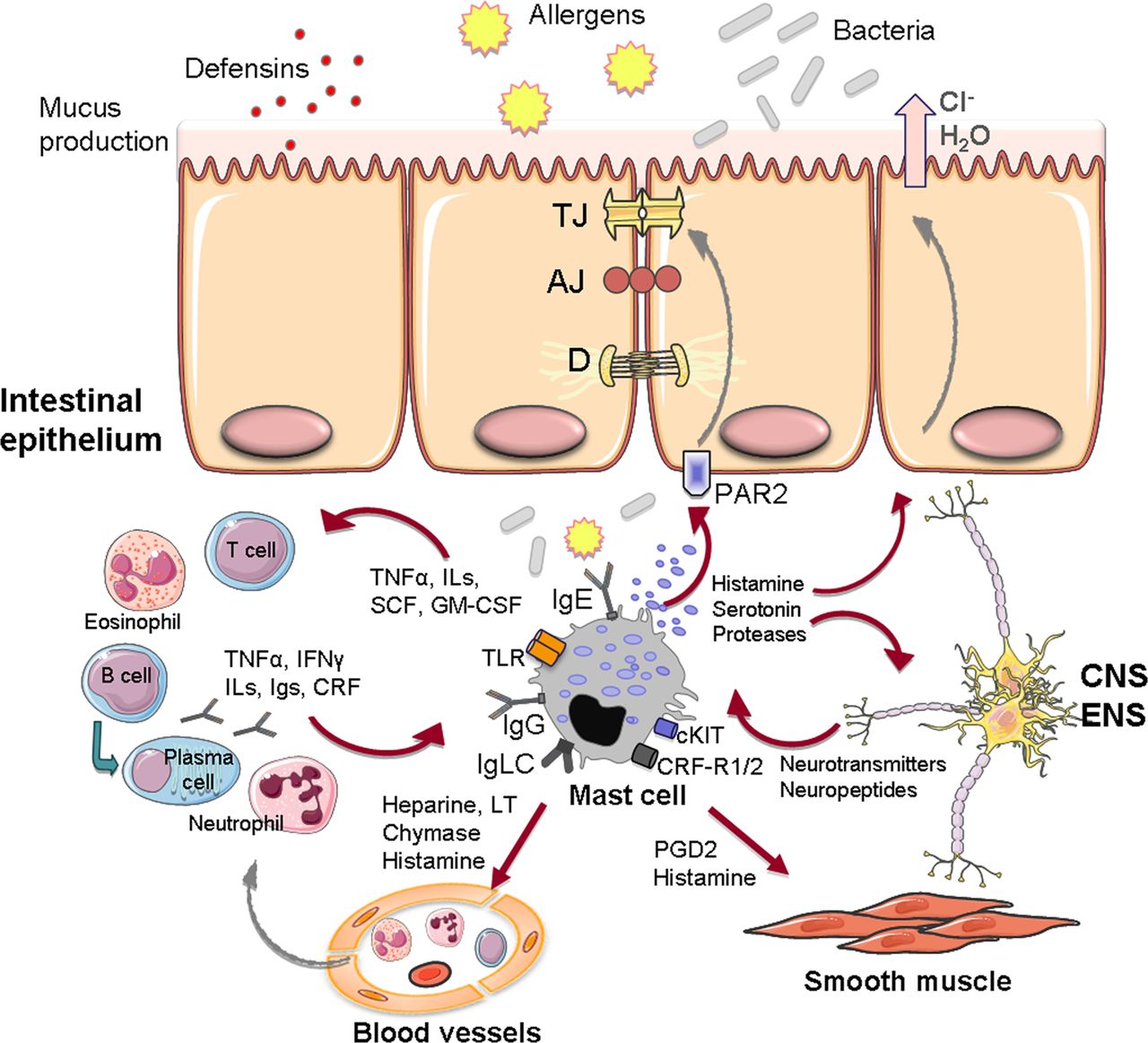
MCs’ contribution to barrier function was first described in animal studies in which increased ion secretion and transepithelial transport of macromolecules was reversed with an MC stabiliser.89 In humans, stress induces the release of MC mediators (tryptase, histamine) to the intestinal lumen39 and increases intestinal permeability, which can be reversed by oral DSCG.42 Stress can severely impact on barrier function and favour intestinal disease, as might be the case for FGIDs. IBS and FD patients experience high levels of anxiety, depression and stress3 and intestinal permeability, as measured by probe excretion assays, has been found altered, primarily in PI-IBS and IBS-D.90 The mechanisms underlying epithelial barrier alterations are not fully understood, but disruption of the proteins that seal the paracellular space seems to play a role. Actually, in IBS, the expression of several tight junctions (TJ) proteins is reduced compared to controls and, in IBS-D, this reduction correlates with MC activation and with common clinical symptoms.91 In FD, the altered expression of cell-to-cell adhesion proteins also correlates with impaired duodenal integrity and with mucosal inflammation.9 MCs’ proximity to the epithelium facilitates tryptase activation of PAR-2 receptors on the basolateral side of enterocytes, leading to redistribution of TJ and increased paracellular permeability to macromolecules.92 Other mediators released by MCs upon activation, such as histamine, chymase and prostaglandin D2, regulate epithelial chloride and water secretion and permeability.93 ,94 MC-mediated intestinal barrier alterations have been also related to neuropeptides, neurotransmitters, hormones (vasoactive intestinal peptide, SP, NGF, oestrogen, oestradiol), and inflammatory mediators (tumour necrosis factor-α, interferon-γ and cytokines) released by other immunocytes93 (figure 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Intestinal barrier function elements and mast cell interactions in the intestinal mucosa. Illustration of the potential mast cell interactions in the regulation of barrier function, including epithelial permeability (through TJ modulation and secretory response), recruitment and activation of other immunocytes, endothelial functions (vascular permeability and blood flow), peristalsis and pain signalling through bidirectional communication with the nervous system. 5HT3R, 5-hydroxytryptamine receptor; AJ, adherens junction; CRFR1/2, CRF receptors 1 and 2; CNS, central nervous system; CRF, corticotropin-releasing factor; D, desmosome; ENS, enteric nervous system; GM-CSF, granulocyte and monocyte colony stimulating factor; IFN-γ, interferon gamma; Igs, immunoglobulins; IgE, Immunoglobulin E; IgG, Immunoglobulin G; IgLC, immunoglobulin free-light chains; ILs, interleukins; LT, leukotrienes; PAR2, proteinase-activated receptor-2; PGD2, prostaglandin D-2; SCF, stem cell factor; TNF-α, tumour necrosis factor alpha; TLR, toll-like receptor; TJ, tight junction.
Role of MCs in IBS cardinal manifestations
MCs in close proximity to nerves in the descending colon were significantly correlated with severity and frequency of abdominal pain/discomfort.7 In another study, mucosal MC infiltration was significantly associated with abdominal bloating frequency and with symptoms of dysmotility-like dyspepsia.74 In contrast, in IBS, there was no correlation between severity or frequency of abdominal pain/discomfort and lamina propria area occupied by MCs, release of tryptase and histamine, and number of degranulated MCs per field.7 ,57 IBS-D patients with rectal hypersensitivity, according to the maximally tolerable pressure to barostat distention, showed significantly lower counts of MCs in the terminal ileum, ascending colon and rectum in one study.57 Park et al57 speculated that this counterintuitive finding was related to tissue desensitisation by MC mediators. More recently, Braak et al51 found no correlation between the sensory thresholds to barostat distention, abdominal pain, bloating, urgency, incomplete evacuation, hard stools, loose stools, frequent and decreased bowel moments and flatulence and MC counts in the colon. An association between duodenal and antral MCs with pain, and postprandial distress syndrome, respectively, has been shown in children with FD.67
Impaired intestinal permeability, and the expression of TJ proteins, has been shown to correlate with pain/discomfort and/or bowel habit.90 ,91 ,95 ,96 Interestingly, tryptase mRNA and protein expression in the jejunum of patients with IBS-D correlated with stool frequency and consistency but not with abdominal pain, whereas the correlation with MC number was poor.91
Caecum MC counts correlated significantly with the fatigue and depression scores in IBS,60 and in some studies, there is a tendency or an association between depression and state of anxiety scores and the number of MCs in patients with IBS-D.57 A significant correlation with antral MC densities with anxiety, depression and somatisation has been reported in children with FD.67 Moreover, the degranulation of MCs in the duodenum appears to be highly sensitive and specific for the identification of adult patients with FD as shown by 100% sensitivity and specificity indicated by an area under the receiver operating characteristic curve of 1.0 for the optimal degranulation rate cut-off values of 30.2% at the duodenal bulb and 36.8% at the descending part of the duodenum.66
Taken together, these findings suggest that interactions between the MCs and the enteric and brain–gut neural networks could be of importance in symptom perception in at least a subgroup of patients with FGIDs.
Targeting MCs: implications for treatment of FGIDs
The MC stabiliser DSCG abolished the effect of acute psychological stress on small bowel permeability in human subjects.42 Aside from experimental studies demonstrating the efficacy of several MC inhibitors to decrease colonic hypersensitivity, in humans, a number of uncontrolled observations,56 and open clinical studies with DSCG, in doses between 600 and 1800 g/day, suggest its clinical benefit for chronic persistent diarrhoea,97 allergic enteritis,98 FD99 and IBS.100–102 However, these studies had several limitations, including poor design, small sample size and selection bias. Likewise, ketotifen has been recently proven to increase the sensory threshold, leading to improved visceral perception, especially in the hypersensitivity IBS group.103 Although preliminary, there is some indication of the clinical benefit of ketotifen and the tryptase inhibitor APC 2059 in UC.104 ,105 Our group has recently finished an open trial (awaiting publication) and a consecutive double-blind, placebo-controlled, clinical assay, with prolonged (6 months) oral administration of DSCG, with promising results in the control of main clinical manifestations in patients with IBS-D (Gastroenterology 2015;148(Suppl 1):S-494).106 In addition, small studies have shown improvement in GI symptoms with DSCG therapy in systemic mastocytosis.107 However, the mechanisms by which MC stabilisation could interfere with IBS clinical response have not been clearly delineated.
Other interventions that block the effects of MC mediators and improve GI symptoms should be considered. In this sense, anti-inflammatory treatment with mesalazine appeared to show improvement in symptom perception in unselected patients with IBS in a small proof-of-concept randomised, double-blind, placebo-controlled trial, in which, in addition, a 36% decrease in MC numbers and a reduction of the number of total immune cells and T cells was observed in the colonic mucosa.108 However, two subsequent large clinical trials differ in the clinical benefit of mesalazine in IBS,109 ,110 and the effect of mesalazine on MC counts and degranulation was not confirmed.110 Furthermore, there was no effect of mesalazine on 5-HT containing enterochromaffin and CD68 cell numbers, although there was a significant increase in CD3 count in the mesalazine group.110 In further reinforcing the role of MC activation in the origin of FGID manifestations, it is important to note that heartburn, cramping, nausea, abdominal pain and diarrhoea are the second most common complaint of patients with mastocytosis, and that H2-histamine receptor antagonists have been quite effective in controlling these symptoms.56 ,98 ,111 A recent proof-of-principle clinical trial confirmed the clinical relevance of these findings showing improvement of abdominal pain and global relief by the H1R antagonist ebastin in patients with IBS (Gastroenterology 2013;144(Suppl 1):S-160).112 Palmitoylethanolamide and other inhibitors of cannabinoid receptors seem efficacious in controlling pain, motor disturbances and inflammation in animal models through modulation of neuronal and non-neuronal cells, including MCs113 ,114 Slow release of vitamin C may be also helpful as it increases degradation of histamine and inhibits MC degranulation in doses not superior to 750 mg/day.111 Natural flavonoids (fisetin, kaempferol, quercetin, rutin, luteolin) and the active alkaloid berberine inhibit the mediator release of MCs in vitro115 and protect intestinal epithelial barrier.114 While some of these products have been shown to be useful in cardiovascular health117 and cancer,118 their clinical efficacy in FGIDs has not been established. There is some evidence of symptomatic response to specific diets in FGIDs, such as low FODMAP and gluten-depleted food.20 However, there is no support for the role of MCs in this symptomatic response with the exception of the benefit after individualised exclusion of foods in FGIDs suffering food allergy.
Finally, the development of more specific and safe blockers or modulators of IgE, IgG or other activation pathways of MC activation, including pathways involved in the selective release of mediators, may offer therapeutic advantages, although their benefit remains to be established.
Conclusion
Current evidence implicating MCs in the pathogenesis and pathophysiology of FGIDs, particularly in IBS, and the contribution of their activation and released mediators to the development of cardinal manifestations, such as epigastric and abdominal pain, and altered defecation is robust, and supports the targeting of MCs in the management in FGIDs.
References
Footnotes
Funding Supported in part by Fondo de Investigación Sanitaria and CIBERehd, Instituto de Salud Carlos III, Subdirección General de Investigación Sanitaria, Ministerio de Economía y Competitividad: CP10/00502 & PI13/00935 (MV); PI11/00716 & PI14/00944 (JS); Centro de Investigación Biomédica en Red de Enfermedades Hepáticas y Digestivas: CB06/04/0021 (MV and JS), and MMW is supported by a FWO postdoctoral fellowship (1248513N).
Competing interests None declared.
Provenance and peer review Commissioned; externally peer reviewed.