Article Text

Abstract
Objectives Habitual diet plays a major role in shaping the composition of the gut microbiota, and also determines the repertoire of microbial metabolites that can influence the host. The typical Western diet corresponds to that of an omnivore; however, the Mediterranean diet (MD), common in the Western Mediterranean culture, is to date a nutritionally recommended dietary pattern that includes high-level consumption of cereals, fruit, vegetables and legumes. To investigate the potential benefits of the MD in this cross-sectional survey, we assessed the gut microbiota and metabolome in a cohort of Italian individuals in relation to their habitual diets.
Design and results We retrieved daily dietary information and assessed gut microbiota and metabolome in 153 individuals habitually following omnivore, vegetarian or vegan diets. The majority of vegan and vegetarian subjects and 30% of omnivore subjects had a high adherence to the MD. We were able to stratify individuals according to both diet type and adherence to the MD on the basis of their dietary patterns and associated microbiota. We detected significant associations between consumption of vegetable-based diets and increased levels of faecal short-chain fatty acids, Prevotella and some fibre-degrading Firmicutes, whose role in human gut warrants further research. Conversely, we detected higher urinary trimethylamine oxide levels in individuals with lower adherence to the MD.
Conclusions High-level consumption of plant foodstuffs consistent with an MD is associated with beneficial microbiome-related metabolomic profiles in subjects ostensibly consuming a Western diet.
Trial registration number This study was registered at clinical trials.gov as NCT02118857.
- SHORT CHAIN FATTY ACIDS
- INTESTINAL BACTERIA
- DIET
- DIETARY FIBRE
- ENTERIC BACTERIAL MICROFLORA
Statistics from Altmetric.com
Video abstract
Significance of this study
What is already known on this subject?
Diet can significantly impact the gut microbiota and metabolome.
Negligible differences in gut microbiota and faecal short-chain fatty acids were previously reported between habitual omnivores and vegans.
Mediterranean diet is a recognised healthy dietary pattern that has not previously been related to the composition of the gut microbiota and related metabolome.
What are the new findings?
Habitual vegetarian and vegan diets promote enrichment of fibre-degrading bacteria in the gut.
Subjects who consume a Mediterranean diet rich in fruit, legumes and vegetables have higher levels of faecal short-chain fatty acids, regardless of the diet type.
Low adherence to the Mediterranean diet corresponds to an increase in urinary trimethylamine oxide levels, a potential risk factor for cardiovascular disease.
How might it impact on clinical practice in the foreseeable future?
Microbiota modulation through consumption of diets rich in diverse vegetable foods offers the prospect of increasing health and mitigating disease risk.
Introduction
The role of diet in shaping the gut microbiota is widely recognised, and several recent reviews provide a comprehensive treatment of the subject.1–6 However, until recently, not many studies have broadly and systematically considered the association between habitual diet and gut microbiota.2 ,7
Some populations eat differently because they have different access to foods, and this can determine significant differences in the taxonomic composition of their gut microbiota, distinguishing agrarian and Western diets.8–10 Specific compositional patterns of the gut microbiota have also been associated with habitual diet, clearly linking different compositions of the microbiota with animal fat and protein-based diets versus vegetable-based diets.11 In addition, several studies have shown an association between consumption of fibre, fruit and vegetables and increased microbial richness, at either taxonomic or gene level.12–14 It is well established that a sudden diet modification can promptly cause a change in the gut microbiome,11 ,15 ,16 although such changes are apparently not sufficiently large or enduring to cause individuals to switch between enterotypes.11 ,17
Although there is a general agreement on the association of some enterogradients18 with vegetable-rich (Prevotella) or protein/fat-rich diets (Bacteroides/Clostridia),11 ,15 ,19–22 little is known about the structure of the gut microbiota in individuals with defined dietary habits such as strict vegans or vegetarians compared with omnivores. Negligible differences were recently found between the gut microbiota of a small-size cohort of omnivore and vegan individuals in the USA.7 In addition, a few self-reported vegetarians had a non-significantly higher abundance of Prevotella compared with Bacteroides.11
Vegetarian and vegan diets confer well-described health benefits.23 ,24 Conversely, residence in industrialised countries and consumption of Western diets, which are mainly omnivore-type diets, apparently lead to a composition of the microbiota that is more associated with different types of disease.3 Unesco has recognised the Mediterranean diet (MD) as an intangible cultural heritage (http://www.unesco.org/culture/ich/RL/00884). It is a nutritionally recommended dietary pattern characterised by high-level intake of fruit, vegetables, legumes, nuts and minimally processed cereals, moderately high consumption of fish, low intake of saturated fat, meat and dairy products and regular, but moderate, consumption of alcohol.25 The MD is a pronounced, but not exclusively vegetarian-oriented dietary style, which has been demonstrated to be beneficial for the treatment of obesity, type 2 diabetes, inflammatory diseases and cardiovascular diseases.26–30
The intestinal microbiome can be considered a useful biomarker of long-term consumption of healthy or unhealthy diets.3 Thus, it is important to determine if and to what extent long-term dietary choices can impact on the composition of the microbiota and how this can influence the production of beneficial microbial metabolites.
To investigate the potential benefits of agrarian and MDs, we retrieved daily dietary information from a cohort of 153 individuals following self-declared omnivore (O), vegetarian (VG) or vegan (V) diets, and found that 88% of all subjects showed medium to high adherence with the MD. Subjects consuming more plant-based foods showed increased faecal short-chain fatty acid (SFCA) levels and a higher proportion of fibre-degrading bacteria in the faecal microbiota. In contrast, the lower adherence to the MD in some omnivore individuals corresponded with higher urinary trimethylamine oxide (TMAO) levels.
Methods
Recruitment and dietary information
A cohort of 153 apparently healthy volunteers was assembled comprising 51 vegetarians, 51 vegans and 51 omnivores distributed across four geographically distant cities in Italy (Bari, Bologna, Parma and Torino). The characteristics of the subjects as well as the recruitment and exclusion criteria are reported in the online supplementary material. Total food and beverage consumption was assessed by means of a 7-day weighed food diary, completed every day for a total of 7 days as previously described.31 Daily intake of macronutrient and micronutrient was calculated using Microsoft Access application linked to the food database of the European Institute of Oncology, covering the nutrient composition of more than 900 Italian foods.32 The nutrient composition of vegetarian products was evaluated on the basis of information given by suppliers or written on the package. Food items consumed were also retrieved and collapsed into food categories.
The level of adherence to the Mediterranean dietary pattern was assessed using the 11-unit dietary score based on tertiles proposed by Agnoli and collaborators.33 The Healthy Food Diversity index (HFD) was calculated as reported previously.34 Median food and nutrient intakes in different diet types and MD adherence levels are summarised in online supplementary table S1.
Sample collection and microbiota analysis
Faecal and urinary samples were collected on the same day of three consecutive weeks, and the three samples were pooled for both the microbiota and metabolome analyses. Library preparation and sequencing were carried out as previously described.35 Details are provided as online supplementary material. The 16S rRNA gene sequences are available at the Sequence Read Archive of NCBI (National Center for Biotechnology Information) (accession number SRP042234).
Bioinformatic and statistical analysis
Raw reads were first filtered according to the 454 processing pipeline. The sequences were then denoised and further filtered using QIIME V.1.8.0 software.36 Operational Taxonomic Units (OTUs) were de novo picked at 97% sequence similarity following the uclust pipeline, and representative sequences of each cluster were used to assign taxonomy through matching against the Greengenes database (V.05/2013). Alpha diversity indices were calculated by using the diversity function in R vegan package (http://www.r-project.org). Weighted and unweighted UniFrac distance matrices were obtained through QIIME, and imported in R to obtain principal coordinates analysis (PCoA) plots.
Correlation analyses, sample clustering and statistical analyses were carried out in R environment. Pairwise Spearman correlations were calculated between microbial genera, dietary and metabolome datasets. The correlation plots were visualised and clustered in R using the Made4 package, and the function heatplot. The statistical package DESeq237 as well as the Kruskal–Wallis and Mann–Whitney tests were used to find significant differences in microbial taxa abundance, metabolome profiles and alpha diversity according to the HFD, the three types of diet and adherence to the MD. Unless stated otherwise, box plots represented the IQRs between the first and the third quartile, with the error bars showing the lowest and the highest value within 1.5×IQR. Correction of p values for multiple testing38 was performed when necessary. Linear regression was used to compare SCFA concentration or intake of the different nutrients/foods and diet (O, V, VG) or MD score by using the glm function in R, which performs a linear regression for a response variable (x) and one or more predictor variables (y), allowing to test for the effect of y on x by correcting for all other relevant variables.
In order to explore the relationship between the genus-level microbiota and the dietary datasets, we carried out principal components analysis (PCA) on the individual datasets and then integrated them using coinertia analysis (CIA),39 which allows identification of the shared biological trends within the two datasets.
Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt),40 a programme that predicts abundances of gene families based on 16S-based structure of the microbiota was used to investigate the functional profiles in microbial communities. For the analysis with PICRUSt, the pick OTUs module was performed at 97% identity in a closed reference way using the Greengenes database (V.05/2013) in QIIME 1.8. Data were normalised for 16S rRNA gene copy numbers, and the metagenomes were predicted. From the inferred metagenomes, KEGG Orthologs (KO) were identified. The KO abundance table was used as input in gage Bioconductor package41 in order to carry out pathway enrichment analysis.
Metabolomic analysis
Gas-chromatography mass spectrometry-solid-phase microextraction (GC-MS/SPME) was carried out according to the protocol previously described42 by using a Chromopack CP Wax 52 CB fused silica capillary column (Chromopack, Middelburg, The Netherlands) and an Agilent gas chromatograph (Agilent Technologies, Palo Alto, California, USA) coupled to an Agilent mass selective detector operating in electron impact mode (ionisation voltage, 70 eV). The identification of the molecules was based on comparison of mass spectra contained in available databases (NIST V.2005 and Wiley V.1996). ChemStation (Agilent Technologies) was used to convert the GC-MS/SPME raw files into netCDF format for further processing with the XCMS toolbox (http://metlin.scripps.edu/).
Urine samples were prepared for nuclear magnetic resonance (NMR) analysis by centrifugation for 15 min at 15 000 rpm at 4°C. Seven hundred microlitre of supernatant were added to 160 μL of a D2O solution of 3-(trimethylsilyl)-propionic-2,2,3,3-d4 acid sodium salt 6.25 mM. After pH adjustment to 7.0 by the addition of HCl or NaOH (0.5 M), the samples were stored at −20°C. Immediately before analysis, the samples were thawed and centrifuged as described above. Faecal samples were prepared for NMR analysis by vortex mixing 40 mg of stool for 5 min with 1 mL of deionised water. The obtained mix was then processed as explained for urine samples. 1H-NMR spectra were recorded at 298 K with an AVANCE III spectrometer (Bruker, Milan, Italy) operating at 600.13 MHz. The HOD (deuterated water) residual signal was suppressed as explained elsewhere. Each spectrum was acquired as described elsewhere.43 The signals were assigned by comparing their chemical shift and multiplicity with the Human Metabolome Database44 and Chenomx software data bank (Chenomx, Canada, V.8). To prevent biases based on hydration of the individuals, data were normalised by means of probabilistic quotient normalisation.45
Results
High-level adherence to the MD, correlating with HFD
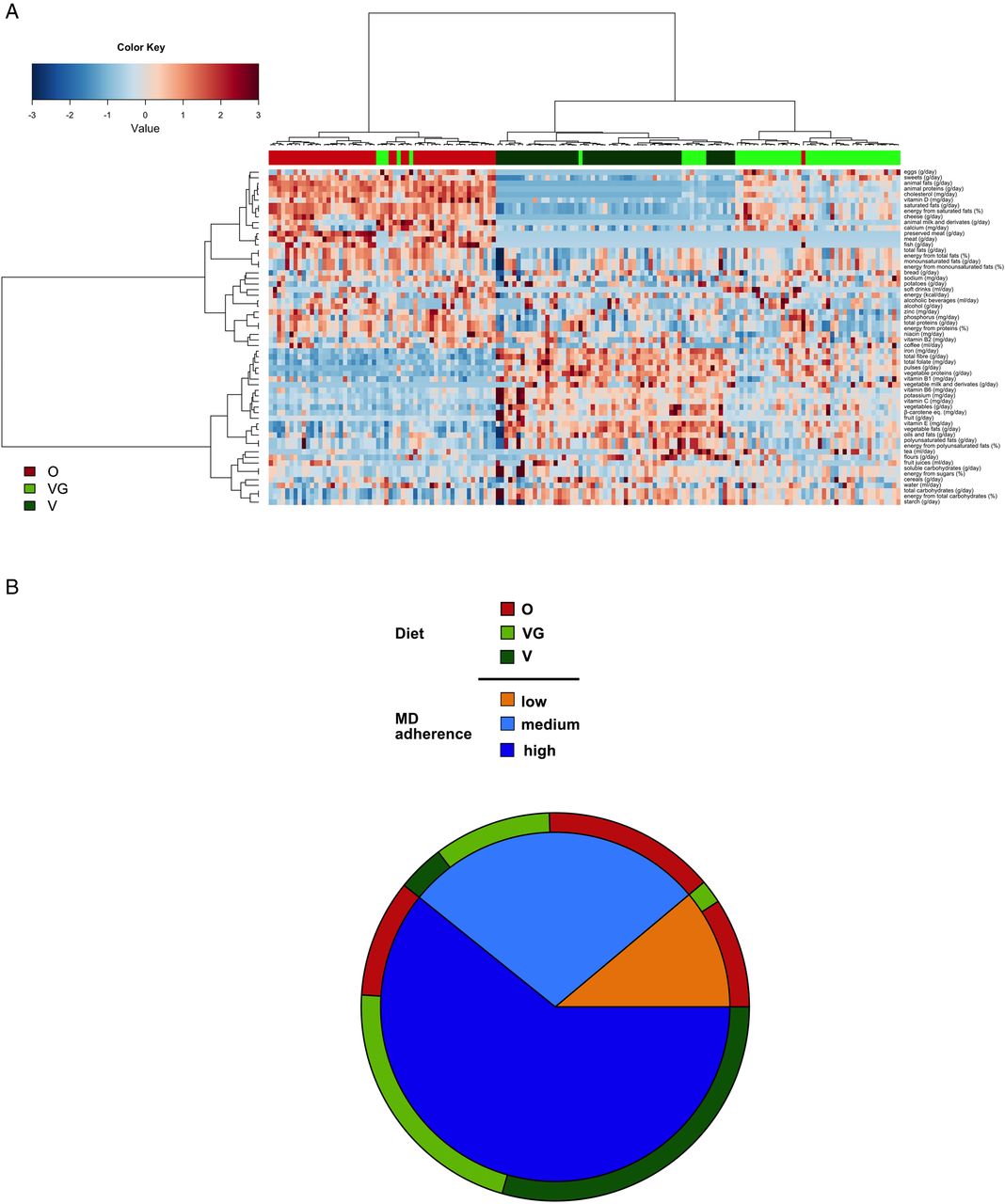
Subjects in this cohort in Italy were able to self-declare their habitual diet category, but were not wittingly aware of their level of adherence to the MD or their HFD. Hierarchical clustering based on habitual diet clearly separated omnivore, vegetarian and vegan individuals with few exceptions (figure 1A). Analysis of the adherence level to the MD33 divided the subjects in three groups with low, medium and high adherence to MD (figure 1B). Only 11% of the subjects (14 omnivores and 3 vegetarians) had low adherence to the MD, while 88% of the vegans, 65% of the vegetarians and also 30% of the omnivores showed high MD adherence rates. Moreover, the HFD index34 values correlated with the adherence to the MD (Spearman's r=0.61, p<0.00001), and were significantly higher in vegetarian and vegan subjects, as well as in subjects with medium and high adherence to the MD (p<0.0001) (see online supplementary figure S1).

Clustering of subjects according to diet (A) and proportion of subjects for each diet type (O, VG, V) assigned to three levels of adherence to the MD (low, medium, high) (B). In the heatplot, hierarchical ward-linkage clustering is based on the Spearman's correlation coefficient of the dietary information abundance. Column bar is colour-coded by diet type. The colour scale represents the scaled abundance of each variable, denoted as Z-score, with red indicating high abundance and blue indicating low abundance.MD, Mediterranean diet; O, omnivore; V, vegan; VG, vegetarian.
Separation by habitual diet and microbiota concords with diet type and MD adherence
The microbiota alpha diversity values were not significantly different between subjects of the three diet types or between subjects showing different levels of adherence to the MD (data not shown). Similarly, there was no clear separation by microbiota composition of omnivore, vegetarian and vegan individuals in PCoA plots based on UniFrac distances (see online supplementary figure S2). However, CIA (figure 2) based on PCA of microbiota composition and diet information revealed a significant relationship between genus-level microbiota composition and habitual diet (RV coefficient=0.18; Monte Carlo p=0.003) involving a significant subset of the microbiota. The first component of the CIA (horizontal) accounted for 50.7% of the variance, and the second component (vertical) accounted for another 11.7%. Although not providing clearly separated clusters, the CIA showed clear gradients of subjects separated according to diet type (figure 2A) and adherence to MD (figure 2B). Although the level of diversity of the subjects based on the structure of their microbiota was clearly not comparable with their strong differences in habitual diet (figure 1), it was noteworthy that their stratification in gradients was consistent when both diet and microbiota were considered (figure 2).

Coinertia analysis combining PCA of microbiota and dietary information. Subjects’ clustering and colouring were done according to the diet type (A) and adherence to the MD (B). Arrow ends of the line indicate sample position in the microbiota dataset, while black dot end indicates sample position in the diet dataset. MD, Mediterranean diet; O, omnivore; PCA, principal components analysis; V, vegan; VG, vegetarian.
Pie charts of family level microbiota composition showed an increase in Prevotellaceae abundance with plant-based diets, although not statistically significant after correction for multiple comparisons (see online supplementary figure S3A). Overall, the Bacteroidetes phylum was more abundant in vegans and vegetarians compared with omnivores (p<0.05); accordingly, higher Firmicutes:Bacteroidetes ratios were found in most of the omnivores (see online supplementary figure S3B).
Microbial genera linked to specific intake of nutrients and dietary patterns
We performed a number of analyses investigating the microbiota in dietary types and differential dietary quality as assessed by HFD. A number of overlapping microbiota signatures were identified. Lachnospira and Prevotella were significantly linked to highest and medium quartiles of HFD values, respectively, and they were both associated to plant-based diets. On the contrary, L-Ruminococcus (Ruminococcus genus assigned to Lachnospiraceae family) was positively associated to omnivore diets (see online supplementary table S2). Plotting the correlation between genera and dietary information, an influence of the diet on the structure of the microbiota could be detected, although not strong in terms of absolute values (figure 3). Three major clusters of genera and three clusters of nutritional information were detected. The first group included foods and nutrients typical of the omnivore diets while the third group included those mainly associated with vegetarian or vegan diets. A small cluster of genera was associated with the vegetable-based diet, with the strongest correlations found for Roseburia, Lachnospira and Prevotella, the last two also displaying a significant negative association with the omnivore diet (p<0.01, see online supplementary table S2). A larger cluster correlated positively with the nutrients of animal origin and negatively with the vegetable-based diet patterns (figure 3). The strongest and most significant associations were shown by L-Ruminococcus and Streptococcus (p<0.001).
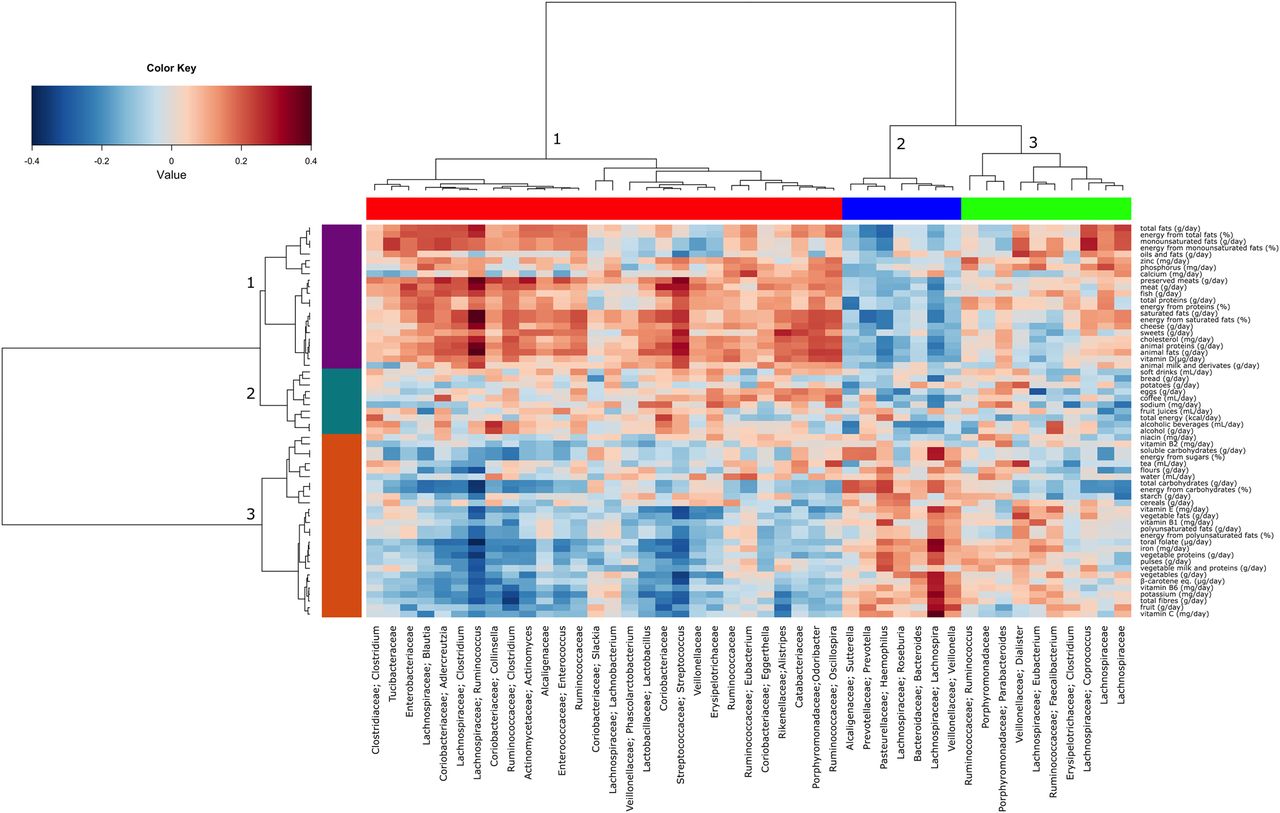
Correlations between microbiota and dietary information. Heatplot showing Spearman's correlations between microbial genera (filtered by a subject prevalence of 20%) and dietary information. Rows and columns are clustered by Euclidean distance and Ward linkage hierarchical clustering. The intensity of the colours represents the degree of association between the taxa and foods/nutrients as measured by the Spearman's correlations.
Correlations between diet, microbiota and metabolome
There was a very clear relationship between faecal SCFA profile and nutrient intake, with the latter variable also splitting into two distinct groups (figure 4A). The faecal levels of acetic, propanoic and butanoic acids (and corresponding esters) strongly correlated with the consumption of fruit, vegetables, legumes (p<0.05) and fibre (p<0.001). By contrast, valerate and caproate concentrations were linked to consumption of protein-rich animal foods and fat (p<0.05). The levels of the three most abundant soluble SCFA showed the same significant increase with increasing adherence to the MD (figure 4B) and in plant-based diets (see online supplementary figure S4). Even within the category of omnivore subjects when analysed separately, clear trends for increase in the levels of the principal SCFA with increasing MD scores were observed (figure 4C). Propanoate and acetate in omnivores were significantly higher in the highest MD adherence category compared with the lowest (p<0.05). Consistent significant associations were found between the volatile fraction of SCFA and adherence to the MD or diet type (see online supplementary figure S5). A list of additional molecules with significantly different (p<0.05) levels according to diet type or MD adherence is provided in online supplementary table S3.

Correlations between short-chain fatty acid (SCFA) levels and diet. Heatplot showing Spearman's correlations between SCFA and dietary information (A). Rows and columns are clustered by Euclidean distance and Ward linkage hierarchical clustering. The intensity of the colours represents the degree of association between the SCFA and foods/nutrients as measured by the Spearman's correlations. Box plots showing abundance of soluble butanoate, propanoate and acetate in faecal samples of all (B) or only omnivore (C) subjects grouped according to adherence to the Mediterranean diet. (*p<0.05 and **p<0.01).
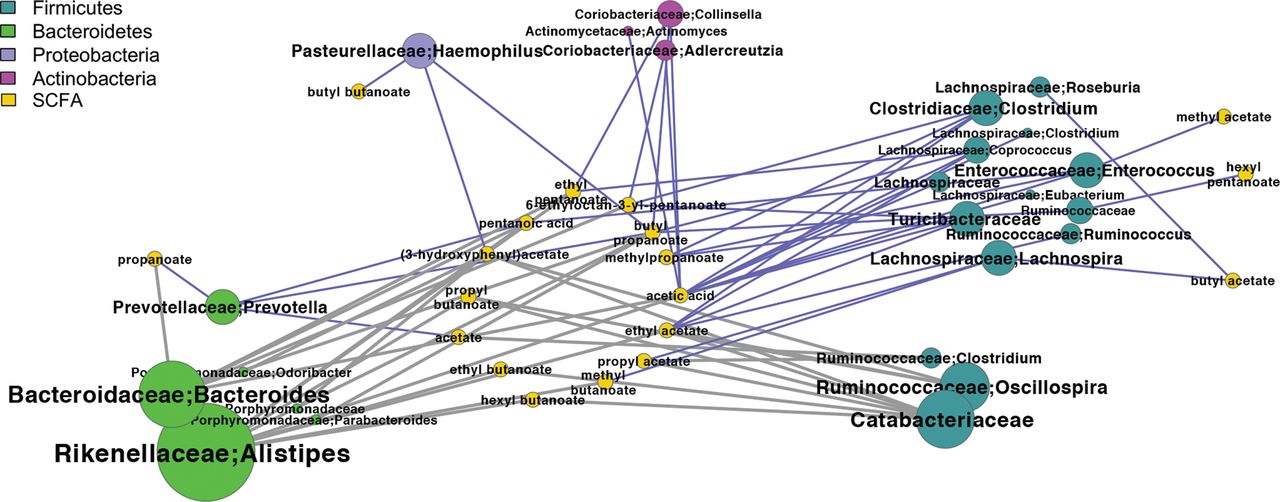
We identified a number of positive correlations between bacterial genera and SCFA levels. In a correlation network, Firmicutes and Bacteroidetes showed the highest number of significant positive and negative correlations with SCFA, respectively (figure 5). The associations between Lachnospira and esters of butanoic and acetic acids were significant (p<0.05). Moreover, Prevotella was the only Bacteroidetes having positive correlations with SCFA (p<0.05).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Correlation between short-chain fatty acid (SCFA) levels and microbiota. Network showing Spearman's correlations between bacterial genera and SCFA. Nodes are coloured according to phylum. Node and node label size is proportional to the number of correlations. Blue edges—positive correlations, grey edges—negative correlations. Only correlations with corrected p value <0.1 are displayed.
Urinary TMAO levels were significantly lower in vegetarian and vegan (p<0.0001) compared with O subjects (see online supplementary figure S6A). In addition, the higher the adherence to the MD, the lower the measured TMAO level (see online supplementary figure S6B). Associations were found between TMAO levels and a number of microbial genera that were more linked to the intake of animal proteins and fat. L-Ruminococcus correlation with TMAO was found to be significant after correction for multiple comparisons (p=0.022).
Diet-driven differences in predicted metagenomes
The pathway enrichment analysis of the predicted metagenomes showed an enrichment in vegans compared with omnivores of folate biosynthesis (ko00790), one-carbon pool by folate (ko00760), fructose and mannose metabolism (ko00051), amino sugar and nucleotide sugar metabolism (ko00520) and other glycan degradation (ko00511) (p<0.05). Interestingly, subjects with high compliance to MD showed enrichment in the same pathways, as well as those for pentose and glucuronate interconversions (ko00040), and propanoate (ko00640) and butanoate metabolism (ko00650) (p<0.05). Moreover, omnivores showed higher abundance of genes in the galactose metabolism pathway (ko00052) compared with both vegans and vegetarians (p<0.05).
Discussion
The remarkable adherence to the MD in the subjects studied here, together with the remarkable link we identified with HFD, make the cohort a good model for a population following a healthy Western diet. Higher adherence levels to the MD were found here than in a larger Italian cohort study33 and also in the multicentre prospective cohort study on the dietary habits of 10 European countries.46 Almost 90% of the subjects recruited in this study showed at least medium adherence while 60% showed high-level adherence to the MD. Remarkably, this latter category included about one-third of the omnivores. A healthy dietary pattern of the subjects (omnivores included) was also supported by the higher intake of total carbohydrates, mainly starch and fibre, compared with the intakes recorded in the European Prospective Investigation into Cancer and Nutrition study.47 The high correlation between HFD34 and MD compliance suggests that the results of this study could be easily extrapolated to other populations (different from Mediterranean), if eating a healthy diverse diet rich in fruit and vegetables.
We provide the first concrete evidence for the interconnection between Mediterranean dietary patterns, gut microbiota and microbial metabolites. Consumption of fruit, vegetables and legumes by subjects with satisfactory adherence to the MD associates here with an increase in faecal SCFA levels, and this is likely boosted by certain bacteria belonging to both Firmicutes and Bacteroidetes that are able to degrade carbohydrates not digestible by the host. By contrast, a lower adherence to the MD is linked to higher urinary TMAO levels.
Wu et al7 recently reported negligible differences in the faecal microbiota composition and plasma SCFA profiles in 15 vegan and 16 omnivore subjects, and found no evidence for diet-driven separations in either measure. With a larger cohort of 153 individuals that also included vegetarians, and with a remarkable adherence to the MD, we were able to detect significant associations between intake of fibre and vegetable foods and faecal SCFA concentrations. Interestingly, we recorded higher median intake of fibre and starch in both omnivore and vegan subjects compared with the US cohort.7 In addition, although we noted that the overall structure of the microbiota was similar in subjects across the different diet types, gradients were detected with subjects stratifying on the basis of diet and microbiota.
SCFA are produced by the intestinal microbiota during fermentation of undigested polysaccharides.48 The protective role of acetate, propionate and butyrate against different types of disease is well documented.49–52 The abundance of SCFA in this study was clearly correlated with the diet. Based on greater intake levels of fibre through fruit and vegetables, vegan and vegetarian diets were associated with higher production levels of SCFA, whereas the animal-based diet supported much lower levels. Accordingly, differences between herbivore and carnivore diets were suggested to be the primary factor driving butyrate production in different animal species,53 and higher SCFA levels were found in native Africans on agrarian diet compared with African–Americans.22 ,54 However, differences in SCFA levels according to MD adherence were not so dramatic compared with differences reported between Western and agrarian cohorts8 ,22 and on extreme dietary changes achieved in interventional studies.15 ,55 Very interestingly, even in the omnivores alone, the higher the adherence to the MD, the higher the faecal SCFA levels. These trends indicate that the MD can be pivotal in providing the appropriate substrates and establishing the catabolic microbes to support the production of SCFA even in an overall omnivore-type diet.
A remarkable degree of interindividual variability in SCFA levels was found in this study compared with the current literature. This may be a feature of the great heterogeneity of the groups considered here, as individuals within these groups have a wide range and variety of fruit and vegetable intake. Moreover, besides the diet, multiple factors can affect human metabolome, such as gender or age.56–58 In spite of such variability, significant differences were found between diet groups, although this within group variance prevents extrapolating the results on individual basis.
Some additional molecules were differentially abundant depending on the diet (see online supplementary table S3). At the state of the art, it is difficult to accurately link most of them to a specific dietary pattern. Esters of SCFA were consistently associated with the plant-based diets (see online supplementary table S3). Conversely, succinate levels were significantly higher in omnivore compared with vegan subjects, and accordingly, genes related to succinate production pathway were enriched in subjects receiving an animal-based diet.15 Higher content of phenolic and indole derivatives, produced by colonic deamination of aromatic amino acids and methylamines, was found in omnivore subjects and in those with low adherence to the MD. Accordingly, a reduced excretion of faecal phenols was observed following a 3-week high-resistant starch diet,59 while a significantly lower production of p-cresol sulfate and indoxyl sulfate were found in vegetarians.60
The same microbiota that was more associated with vegetable-based diets tended to correlate with SCFA levels. Prevotella among Bacteroidetes and Lachnospira among the Firmicutes appeared as the most probable candidates for fermentation of carbohydrates, leading to a higher SCFA production. Collectively, the abundance of the phylum Firmicutes was associated with a high intake of animal-associated foods, but individual members of this phylum showed a higher number of positive correlations to individual SCFA than phylum Bacteroidetes, corroborating the hypothesis that their role in polysaccharide breakdown has been underestimated.61 As shown here, their gene pool may be as useful as that of Bacteroidetes (ie, Prevotella) in Western subjects eating Mediterranean-type food. Consistent with this, the predicted metagenomes showed a significant increase in abundance of genes associated with polysaccharide degradation and SCFA metabolism with increasing adherence to the MD patterns and plant foods consumption.
Lachnospira is one of the core genera of the gut microbiota4 and recognised as the only Firmicutes able to degrade pectins beyond Faecalibacterium and Eubacterium.56 Decrease in Lachnospira abundance was recently highlighted in the gut microbiota of subjects with cholesterol gallstones62 as well as in the rectal mucosa of patients with HIV,63 tentatively suggesting that the decrease in these SCFA-producing Firmicutes might be linked to predisposition to disease.
Most of the studies and reviews of gut microbiota in relation to diet report an increased level of Prevotella in response to consumption of an agrarian diet or on fibre administration.2–4 ,8 ,10 ,11 ,64 Levels of Prevotella were not significantly different between omnivores and vegans.7 However, correlations between Prevotella abundance and fruit and vegetable intake were found in the present study. Remarkably, the Prevotella:Bacteroides ratio in these subjects was higher than reported for elderly Irish subjects,12 American11 and Danish adults,65 and it is very likely linked to the higher fibre and starch intake in the Italian subjects.
A certain number of bacteria were associated with a diet based on animal foods and with saturated and monounsaturated fat intake. Some of the Bacteroidetes, such as Alistipes, have been already reported as increasing as a function of elevated consumption of animal foods,12 ,15 and showed a clear negative correlation with vegetable nutrients and faecal SCFA. Moreover, L-Ruminococcus, associated with fat intake and omnivore diet, was also correlated with urinary TMAO levels. Consistently, Ruminococcus torques (Lachnospiraceae) was recently found at increased levels in mice consuming a high fat and sugar diet and with susceptibility to Crohn's disease.66 TMAO is linked to risk of atherosclerosis and cardiovascular disorders.67 ,68 Eggs, beef, pork and fish are the major sources of carnitine and choline that are converted by the gut microbiota to trimethylamine, then oxidised in the liver and released into circulation as TMAO.69 ,70 The diet–TMAO association found in this study provides valuable clues on the possible modulation of TMAO by the diet, and although no data were collected here on plasma levels, the adherence to the MD offers encouraging paths towards the reduction of its levels.
Properly controlled interventions could confirm the gut-related health benefits of MD and allow drawing conclusions on individual basis. Notwithstanding this, we provide here tangible evidence for the impact of a healthy diet and a Mediterranean dietary pattern on gut microbiota and on the beneficial regulation of microbial metabolism towards health maintenance in the host. Western omnivore diets are not necessarily detrimental when a certain consumption level of vegetable foods is included. Healthy diets can, therefore, contribute to beneficially modulate host–microbe interactions and to establish effective pathways to prevent disease.
References
Supplementary materials
Press release
Files in this Data Supplement:
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online methods
- Data supplement 2 - Online figures
- Data supplement 3 - Online tables
Footnotes
Contributors DE was responsible for the development of the study and preparation of the manuscript. DE, EN, LC, PB and MG contributed to the study design and coordinated the recruitment sites. RDC, IF, CL and ST recruited subjects and collected biospecimens. FDF and ALS performed the 16S rRNA gene sequencing. NP collected and analysed the dietary information. LV, LL and DIS performed the metabolome analysis. FDF and IBJ carried out the bioinformatics and statistical analyses and generated the manuscript figures. PWOT supervised the data analysis and contributed to manuscript preparation. MG is the project coordinator. All authors revised and approved the final version of the manuscript.
Funding This study was supported by the Italian Ministry of University and Research (MIUR) programme PRIN 2010–2011 (grant number 2010WZ2NJN). Work in PWOT's laboratory was supported by Science Foundation Ireland through a Centre award to the Alimentary Pharmabiotic Centre, and by an FP7 award to the NuAge project.
Competing interests None declared.
Patient consent Obtained.
Ethics approval The study was approved by the Ethics Committee of (a) Azienda Sanitaria Locale (Bari) (protocol N.1050), (b) Azienda Ospedaliera Universitaria of Bologna (protocol N.0018396), (c) Province of Parma (protocol N.22884) and (d) University of Torino (protocol N.1/2013/C).
Provenance and peer review Not commissioned; externally peer reviewed.