Article Text

Abstract
The GI tract is the most exposed organ to proteases, both in physiological and pathophysiological conditions. For digestive purposes, the lumen of the upper GI tract contains large amounts of pancreatic proteases, but studies have also demonstrated increased proteolytic activity into mucosal tissues (both in the upper and lower GI tract), associated with pathological conditions. This review aims at outlining the evidences for dysregulated proteolytic homeostasis in GI diseases and the pathogenic mechanisms of increased proteolytic activity. The therapeutic potential of protease inhibition in GI diseases is discussed, with a particular focus on IBDs, functional GI disorders and colorectal cancer.
- INFLAMMATORY BOWEL DISEASE
- INFLAMMATORY BOWEL SYNDROME
- INFLAMMATORY MECHANISMS
- ENZYMOLOGY
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Key messages
Protease inhibition as therapeutic approach in intestinal pathologies: what should we consider?
Profiles of active proteases have to be performed in pathological tissues in order to define the best molecular targets for therapeutic intervention
Large spectrum inhibitors might have severe side effects
Promote the expression or delivery of natural endogenous inhibitors could be a safe therapeutic option
Local versus systemic delivery would have to be considered
The use of food-grade bacteria as carrier for the delivery of therapeutic proteins has been proposed
Protease targets for IBD
MMPs inhibitors have been abandoned
Serine proteases are considered
Ubiquitin–proteasome system inhibitors are considered
Protease targets for IBS
Trypsin inhibition
Tryptase inhibitors
Protease-Activated Receptor (PAR1)/PAR2 antagonists
Introduction
Proteases represent up to 2% of the human genome, with 500–600 different proteases that have been identified. Through the evolution, proteases have adapted to the different conditions that characterise complex organisms: pH variation, oxydo-reduction environment, temperature, etc. Proteases specifically cleave proteins at their extremities (N-terminal or C-terminal regions) and are then called exopeptidases, or in the middle of the proteins, being qualified then as endopeptidases. Depending on their proteolytic mechanism, human proteases are classified as serine, threonine, cysteine, aspartic or metalloproteases (figure 1 and table 1). Some of them are secreted and released in the extracellular milieu, while others have intracellular functions and exclusively remain inside the cells (figure 1).
Proteases identified in GI tissues and cells, and disease-associated upregulation

Representation of human cell proteases according to their catalytic mechanism and their intracellular or extracellular representation. MMPs, matrix metalloproteinases.
Proteases and protease inhibitors of the GI tract
Proteases
In the GI tract, proteases are heavily present, both in the lumen and deeply into the tissues.1 Pancreatic proteases (trypsins, chymotrypsin, elastase, etc) are released into the lumen of the upper GI tract, where they exert digestive functions. The microbiota constitutes also an important source of proteases (figure 2). Bacteria, yeasts and helminths potentially present in the intestinal lumen produce and release proteases.2 For some pathogens such as pathogenic forms of Escherichia coli or the enterotoxigenic Bacteroides fragilis, their ability to release proteases is crucial for their pathogenicity. Serine, cysteine, aspartic and metalloproteases are expressed and released by the microbiota (table 2).2 However, it is interesting to note that when the nature of the proteases present in human faeces was investigated, only host proteases were identified. These findings could question the contribution of microbial proteases, to overall luminal proteolytic activity in the gut. Another study has established significant associations between specific bacterial subgroups and faecal protease activity, suggesting that microbiota composition could affect intestinal proteolytic homeostasis. More recently, forms of secreted proteases have been identified in intestinal epithelium: mesotrypsin mRNA is found in human intestinal epithelial cells3 and trypsin activity is released by cultures of those cells (Vergnolle, personal communication). Other resident cells of the intestinal mucosa produce and release proteases (figure 2). For instance, the major protein content of mucosal mast cells is proteases. Mast cells release different forms of proteases: tryptase and chymase for the vast majority, and also cathepsin G and granzyme B. Resident macrophages also produce and/or release different forms of proteases: matrix metalloproteinases (MMPs) (MMP-12 among other MMPs), caspase, cathepsins L and D.1 In the inflamed gut, inflammatory cells are another major source of proteases, which they use to degrade extracellular tissues and intracellular particles, thereby increasing their phagocytic properties.4 Upon inflammatory cell infiltration and activation, tissue proteolytic activity is considerably increased. Neutrophils in particular release massive amounts of elastase, proteinase-3 and cathepsin G5 (figure 2). Finally, all resident cells of the GI tract express intracellular proteases: caspases, which have fundamental roles in cell apoptosis, and autophagins, which are the proteolytic enzymes responsible of autophagy processes6 (table 1). A special case can be made for deubiquitylases, which are crucial regulators of intracellular protein turnover through the proteasome system. These enzymes present in all cell types, are either cysteines or metalloproteinases and target ubiquitinylated proteins, thereby changing their degradation fate inside the cell.
Major identified pathogen-associated microbial proteases

Source of proteases in the GI tract.
Although specific proteases can be detected in tissues, the cellular origin of most proteases is quite difficult to define, and no study so far has determined the origin of proteases detected in intestinal tissues. The site of action of a given protease is also debated. As of today, one can only specify the possible site of action of a given protease.
Another level of difficulty in studying proteases is that for activity tests, substrates are never fully specific of one protease, neither are their inhibitors. Therefore, the proteolytic activity that is measured is possibly due to a mix of proteases and cannot be attributed to one specific protease.
Protease inhibitors
Protease inhibitors have coevolved with proteases, in order to control their destructive nature. Natural endogenous protease inhibitors are particularly present in the GI tract.1 They are either circulating inhibitors produced at distance from the GI tract (mostly in the liver), or are produced on site, by intestinal epithelial cells or infiltrated inflammatory cells (table 3). Serpins A1, A3, A4, E1 and C1 are circulating protease inhibitors inhibiting serine proteases such as trypsins, chymase, tryptase, elastases, kallikreins and cathepsin G (table 3). Secretory leucocyte protease inhibitor (SLPI) and elafin are produced in situ by intestinal epithelial cells or leucocytes. Both inhibit elastase and proteinase-3, while SLPI also inhibits trypsin, chymotrypsin, cathepsin G, tryptase and chymase7 (table 3). Tissue inhibitors of metalloproteinases (TIMPs) are ubiquitously produced, TIMP-1, TIMP-2, TIMP-3 and TIMP-4 are present in the GI tract, where they inhibit a number of different MMPs8 (table 3). The caspase-9 inhibitor, which is a cellular inhibitor of apoptosis protein-2 (c-IAP2) is also ubiquitously produced by cells present in intestinal tissues.
Endogenous protease inhibitors detected in the GI tract
Proteases and intestinal physiology
The roles and functions of proteases and their inhibitors under physiological conditions have been poorly investigated. While digestive proteases are released into the lumen of the upper GI tract for digestive purposes, intestinal microbes largely inhibit them as they progress down to the tract.9 In addition to their physiological role in digestive process, constitutive expression of some proteases seems also to be necessary to intestinal homeostasis. Matriptase for example, is a trypsin-like protease that colocalises with E-cadherin in intestinal epithelial cells. Mice deficient for matriptase expression specifically in intestinal epithelial cells develop from birth diarrhoea, and then later in life develop megacolon and colitis.10
Proteases from the A Disintegrin And Metalloprotease (ADAM) family also seem to play roles in maintaining intestinal barrier function. ADAM-19 colocalises with the tight junction-associated protein zonula occludens-1,11 ADAM-17 deficiency in human induces bowel dysfunctions.12 Cathepsin K-deficient mice showed a disrupted expression of Occludin, a deposit of type IV collagen at the basement membrane and an increased expression of E-cadherin at the apical junction, all together suggesting barrier dysfunctions.13
Mucus formation and properties also seem to be tightly regulated by endogenous proteases. Recently, a study has demonstrated that in contrast to physiological states, mice deficient for the metalloproteinase meprin β has an attached mucus layer in the small intestine, which can be released by the addition of meprin β.14 In the small intestine, mucus is secreted attached to the goblet cells, and requires a protease meprin β, to be detached from the epithelium. This example illustrates the importance of some proteases for mucus properties, and mucosal homeostasis.
Dysregulated proteolytic homeostasis in GI diseases
Because of the large distribution of proteases in the GI tract, and their tight control by endogenous protease inhibitors, association of dysregulated proteolytic homeostasis with GI pathologies has often been investigated (table 1).
IBDs including Crohn's disease and UC were the first diseases to be investigated, initially because of the additional source of proteases represented by infiltrated and activated inflammatory cells. The expression of a very large number of proteases is upregulated in IBD.1 Protein or mRNA expressions of proteases from infiltrated immune cells (neutrophil elastase, proteinase-3, cathepsin G, tryptase, chymase or granzymes) are obviously increased in inflamed tissues from patients with IBD (table 1). Being involved in tissue remodelling, a process of major importance in IBD, MMPs expression is also significantly increased both in Crohn's disease and UC, while ADAMTS proteases expression is unchanged.1 ,15 ,16 Inappropriate induction of cell death through apoptosis or autophagy is also involved in IBD, and proteases involved in such processes (caspases, autophagins) are upregulated in IBD, particularly in UC.17 Genetic evidence supporting the association of proteases and protease inhibitors genes with IBDs was raised in a systematic review. In that study, 75 genes coding for proteases and 7 genes coding for protease inhibitors were retained for Crohn's disease, while for UC, 14 proteases and 4 protease inhibitors genes were retained.18 Among the identified genes, proteins of the ubiquitin–proteasome system were top ranked, and further studies have identified single nucleotide polymorphism in five of those proteins (CYLD, USP40, USP3, DAG1 and APEH) associated with IBD.19 The expression of protease inhibitors in IBD is rather conflicting, reporting increased, decreased or stable levels of expression for serpins,1 elafin or SLPI.20–24 TIMP-1 and TIMP-2 seem to be consistently increased in UC and Crohn's disease,16 while TIMP-3 is decreased in Crohn's disease25 ,26 and c-IAP2 is decreased in UC.17
One major problem with most of the studies that have investigated protease expression in colonic tissues of patients is that this does not reflect the function of proteases associated with the disease. Indeed, mRNA or protein expression may be increased, but depending on the presence of endogenous inhibitors in tissues, the biological activity of proteases might remain the same. Similarly, investigating mutations on protease genes does not provide answers on the function of the protein. Definitive answers on the role of proteases associated with disease states have to come from studies investigating the in situ net activity of proteases. Elastolytic activity has been investigated in tissues from patients with IBD,22 demonstrating that elastase activity was upregulated, mostly in the mucosa. Surprisingly, elastolytic activity was upregulated both in inflamed tissues from patients with Crohn's disease or UC and in non-inflamed parts of the colon of those patients, where no inflammatory cell infiltration was detected. Interestingly, when in situ zymography for elastolytic activity is performed in human colonic tissues of healthy and Crohn's disease patients, the strongest elastolytic activity is detected on intestinal epithelial cells (figure 3A). These two observations made on tissue proteolytic activity suggest that elastase might not originate exclusively from infiltrated inflammatory cells, and provide unexpected directions to investigate the role of elastase in the context of IBD. Only few studies have investigated protease activities in IBD. A recent study has shown that increased MMP activity in tissues from patients with IBD was restored to control levels after infliximab treatment.27 Trypsin activity was also increased in tissues from patients with Crohn's disease and UC.28 Other studies have investigated the proteolytic activity in stools of patients with IBD, reporting an increased activity, associated with dysbiosis.2 However, depending on the faeces collection and conservation methods, variable results could be observed in faecal proteolytic activity.

In situ proteolytic activity (elastolytic in A, trypsin-like in B) performed as previously described in ref. 20, in human colons of healthy individuals, patients with IBS and patients with Crohn's disease.
To a lesser extent, protease expression has been investigated in tissues from patients with IBS. The expression of specific serine proteases such as tryptase,29 elastase,30 trypsin28 ,31 or cathepsin G32 were significantly increased in tissues or in the faeces of patients with IBS, compared with healthy controls. Two types of cysteine proteases (calpain-8 and proteases from the proteasome) were also upregulated in tissues from patients with IBS, compared with controls.33 ,34 But here again, very few studies have investigated the resultant proteolytic activities in tissues or faeces of patients with IBS. Trypsin-like activity seems to be upregulated in tissues from patients with IBS, compared with healthy controls, with a predominant activity in intestinal epithelial cells, as observed by in situ zymography (figure 3B). Faecal protease activity was found upregulated in faeces from patients with IBS30 and association between proteolytic activity and specific intestinal bacterial groups has further been established.35
In colorectal cancer as well, the expression of a number of proteases was upregulated (table 1), but the proteolytic activity associated with colorectal cancer tissues is for the most part unknown. Among the upregulated proteases in colorectal cancer, serine proteases are well represented, but caspases, cathepsins, calpains, deubiquitinases and MMPs are also prevalent (table 1).
Mechanisms of action of proteases in GI diseases
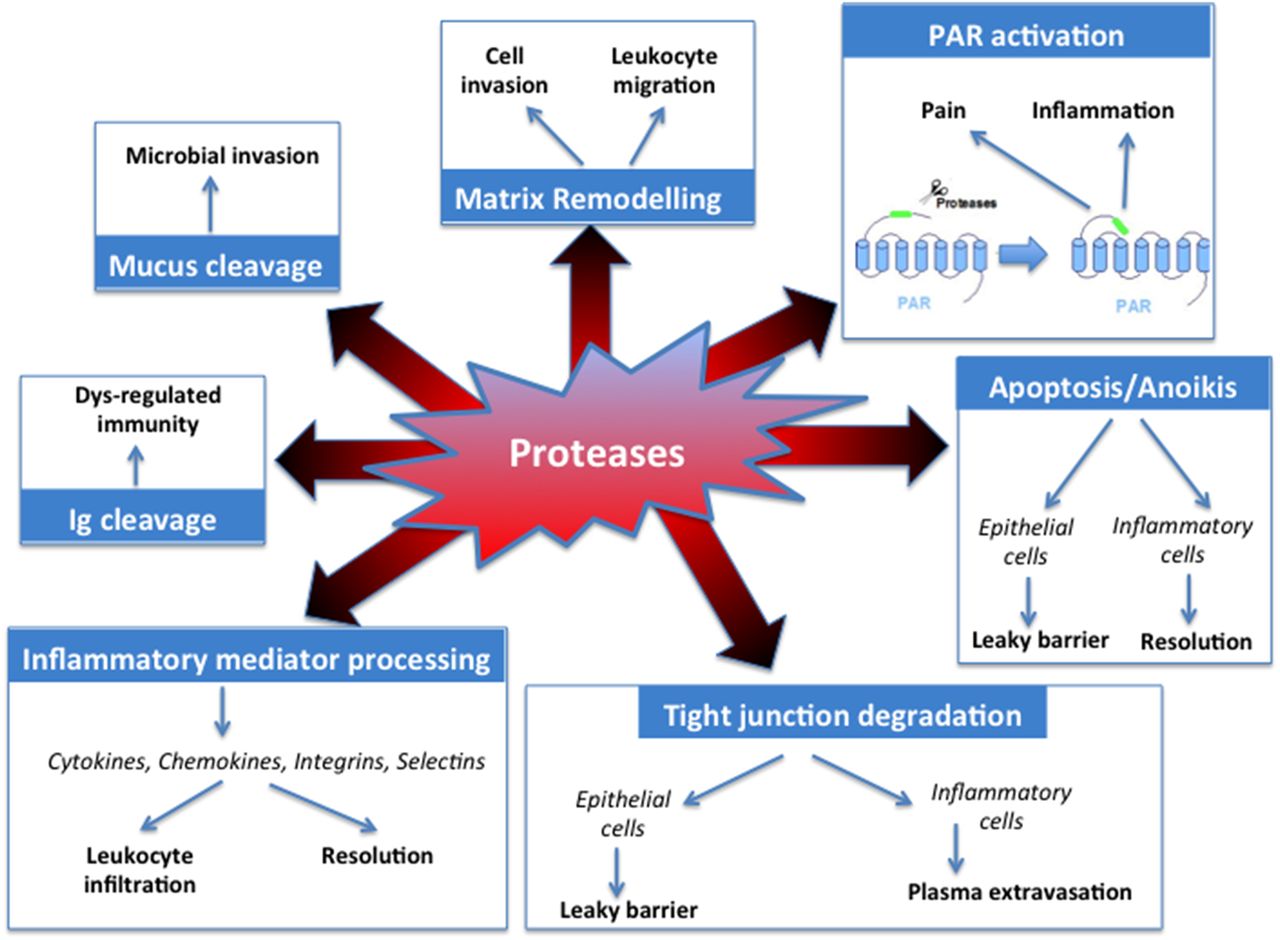
Proteases present in diseased intestinal tissues dispose of several mechanisms of action to participate in pathogenesis or symptoms generation. They act by proteolytic processing of other molecules (mediators, receptors), thereby inducing a number of intracellular signals (figure 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mechanism of action of proteases in GI diseases. PAR, protease-activated receptor.
Receptor activation
In the GI tract, the receptors that have been mostly studied belong to the family of protease-activated receptors (PARs).36 ,37 These receptors are ubiquitously expressed in the GI tract (present in intestinal epithelial cells, in neurons, in infiltrated inflammatory cells, in mast cells, in fibroblasts, etc).37 They are activated by the proteolytic cleavage of their extracellular N-terminal domain, which releases a new N-terminal domain that acts as a tethered ligand to induce intracellular signals.38 ,39 Members of the PAR family (PAR1, PAR2, PAR3 and PAR4) can be activated by serine, and cysteine and metalloproteinases.40 Activation of PARs in the GI tract induces a wide array of pro-inflammatory, pronociceptive and proliferative effects (figure 4). In the gut, PAR activation is able to modify a number of physiological functions: ion exchange,41 motility,42 nociception,43 permeability,44 ,45 secretion, etc. The involvement of PAR activation in GI diseases has been proposed for IBD, IBS and colorectal cancer.46 ,47 Because PARs are expressed both on the apical and basolateral sides of intestinal epithelial cells, these receptors might be activated both by lumenal proteases (including microbial proteases) and by tissue proteases.37
Elastase seems to have receptor-dependent effects involving another type of receptor: the Gram-negative bacteria receptor toll-like receptor-4. Proteolytic activity is necessary to this effect, but the exact mechanism is still unknown.48
Cathepsin G interacts with the G protein-coupled formyl peptide receptor, leading to the activation of mitogen-activated protein kinase (MAPK) pathways.49 However, the pathophysiological consequences of this activation are still unclear.
Inflammatory mediators processing
Proteases may also modulate the bioactivity of inflammatory mediators. This is the case for cytokines, chemokines and their cognate receptors. Proteolytic cleavage increases the bioactivity of chemokines and cytokines by promoting the processing of an inactive precursor, thereby increasing their pro-inflammatory or chemotactic properties. For example, this has been shown for CXCL-8 and CXCL-5, which respectively can be cleaved by proteinase-3 and cathepsin G, the truncated forms of these chemokines having higher chemotactic activity towards neutrophils.50 ,51 Proteinase-3 is also known to activate interleukin (IL)-1, IL-18 and tumour necrosis factor (TNF)-α.52–54 However, proteases can also have opposite effect, degrading cytokines: IL-6 is inactivated by cathepsin G and proteinase-3,55 while elastase and cathepsin G both degrade mature TNF.56 The net effect of proteolytic modifications of chemokines and cytokines, particularly in the context of IBD still has to be clarified. The initial steps of leucocyte recruitments (ie, rolling and adhesion events) might also be tightly controlled by proteases. Selectins, which are expressed at the cell surface, where they initiate the rolling signals, are shed by metalloproteinases (ADAM-17), by stromelysin, collagenase and chymotrypsin,57 ,58 but not by other serine proteases.59 Proteases also regulate the next step of leucocyte recruitment, which involves integrins. Cathepsins are able to cleave members of the integrin family, inhibiting the attachment of migrating cells to extracellular matrix components.60 Here again, the net effect of proteolytic modifications on diapedesis and migration of leucocytes still has to be clarified, but this mechanism of action could play a central role in inflammatory and cancer pathologies.
Apoptosis and anoikis
Caspases and autophagins play essential roles in programmed cell death, which is an important process in chronic inflammatory diseases and cancer. Proteases such as thrombin and granzymes are also able to induce apoptosis or anoikis.44 ,61 When apoptosis is induced in inflammatory cells, this process favours the resolution of inflammation. Protease-induced neutrophil apoptosis would therefore be protective in the context of chronic intestinal inflammation. In contrast, epithelial cell apoptosis leads to a decreased barrier function.44 In that case, protease-induced apoptosis would further feed inflammatory response in the gut, by favouring a leaky barrier, and further penetration of luminal content.
Tight junction degradation
Proteases have been shown to disrupt cell–cell interactions. Therefore, depending on their cellular target, proteases can potentially influence transmigration and microvascular leakage by acting on endothelial cells, or proteases can influence intestinal barrier functions by acting on intestinal epithelial cells. Some proteases such as chymase are able to alter tight junction proteins (ZO-1, occludins)62 or in the case of elastase-2A, a form of chymase, to directly cleave proteins important in barrier functions.63 Adherent junctions seem also to be the targets of some proteases. This is for example the case for neutrophil elastase, which upon the transepithelial passage of neutrophils in inflammatory conditions,64 cleaves the E-cadherin protein. However, neutrophil elastase is unable to cleave tight junction proteins.65 ,66 Other proteases overexpressed in inflammatory conditions might be able to degrade tight junction proteins, although the question of the accessibility of those proteases to tight junction proteins has not really been addressed in vivo. One can question whether proteases could have a direct access to tight junction domain proteins, or whether the effects of proteases on barrier functions are rather mediated by the activation of receptors. Indeed, in the case of thrombin and trypsin, their effects on increased intestinal permeability are mediated by PAR1 and PAR2 activation.44 ,45
Matrix remodelling
The extracellular matrix is a highly dynamic structure, which interacts with cells to regulate proliferation, migration and differentiation. Cleavage of extracellular matrix components constitutes the main regulatory process of these functions. MMPs, ADAMs and ADAMTS are the main enzymes involved in extracellular matrix remodelling. Their activities are controlled by TIMPs (table 3). Excessive extracellular matrix degradation, as observed in chronic inflammatory disorders such as IBD or in colorectal cancer, causes tissue destruction, inflammatory cell infiltration, fibrosis and metastasis.8
Mucus cleavage
Mucus is a major component of mucosal barrier. It efficiently protects host tissues from their luminal content. Mucins are large highly glycosylated proteins that constitute the major component of mucus. Defective mucus layer leads to pathophysiological mechanisms including chronic inflammation and infection. Digestive enzymes are usually unable to digest the glycans composing the mucus, thereby leaving mucins intact.67 Probiotic bacteria such as Lactobacillus and Bifidobacterium do not release proteases that can cleave the MUC2 mucin, the mucus core protein,68 while others, such as Akkermansia muciniphila, are able to degrade mucins.69 Proteases from bacterial pathogens such as Porphyromonas gingivalis,70 from parasites such as Entamoeba histolytica71 or nematodes such as Trichuris muris72 degrade mucus barrier. Under pathophysiological circumstances such as IBD, where proteolytic activity is largely increased in the mucosa, it is reasonable to think that proteases (microbial or host proteases) modify mucus properties.
Immunoglobulin cleavage
Immunoglobulins are sensitive to proteases. A number of studies have demonstrated that bacterial proteases are able to degrade both IgG and IgA, the immunoglobulins the most present at the intestinal mucosa surface.73–75 Indeed, a specific subclass of microbial proteases called ‘IgA proteases’ constitutes a group of extracellular endopeptidases. In pathologies-associated dysbiosis, microbial proteases might then be able to modify the composition and function of resident immunoglobulins and therefore, to modify intestinal immune response. In vivo degradation of immunoglobulins in the intestinal mucosa has never been demonstrated, and one can only speculate on whether bacterial proteases might act on immunoglobulins from the luminal side or whether they could penetrate the tissues. It is not known yet whether host intestinal proteases are also capable of immunoglobulin degradation in an immune-related pathological context.
Protease inhibition as possible treatments for IBD
Overall, considering all their mechanisms of action, proteases associated with IBD exert rather pro-inflammatory properties: they potentiate cytokines and chemokines pro-inflammatory properties, they remodel extracellular matrix to allow leucocyte infiltration, they degrade tight junction proteins inducing plasma extravasation and increased intestinal permeability, they induce apoptosis in intestinal epithelial cells and it is known that activation of PAR1, PAR2 and PAR4 in the colon leads to pro-inflammatory effects.37 ,40 Taken together, these facts suggest that protease inhibition could have strong therapeutic benefits to treat IBD. However, considering the large number of proteases that have been found upregulated in IBD (table 1), and their diverse functions, it is quite difficult to identify single molecular targets among all those proteases. As previously discussed, one major step would be to define which proteases are overactivated in pathological situation, and to establish the profile of IBD-associated overactivated proteases.
One option could be to consider large spectrum protease inhibitors as new therapeutic approach for IBD. However, large spectrum inhibitors might also bear a number of side effects. From all the families of proteases that are upregulated in IBD, MMPs have raised some interests, mainly due to the fact that synthetic inhibitors have been developed for cancer research. MMP inhibitors demonstrated good anti-inflammatory properties in animal models of colitis, but in human, they appeared to be more efficient at helping mucosal healing and extracellular matrix restructuration. MMPs are important factors of extracellular matrix remodelling. Inhibition of proteases implicated in matrix turnover could therefore induce tissue fibrosis. More surprisingly, the use of MMP inhibitors has revealed antitumorigenic and anti-inflammatory effects for some MMPs.76 These data identify MMPs as antitargets for inflammation and cancer rather than targets.
Upon active protease identification, studies have identified some interesting targets in IBD. Elastase is one of them, as its activity is dramatically increased in IBD and elastase has demonstrated a large number of pro-inflammatory effects. Trypsin activity might be another interesting proteolytic target as more aggressive disease and rapid progression to surgery was observed in patients with UC bearing a serpin A1 (or α-1-antitrypsin) deficiency.77 For both targets, instead of raising synthetic inhibitors, which might bear off-target effects, a better option might be to favour the expression of natural endogenous inhibitors of these targeted proteases. Re-equilibrating the protease–antiprotease balance in the inflamed gut by delivering natural endogenous protease inhibitors, which are down-regulated in disease, could constitute a safe and efficient therapeutic option. One challenge though would be to deliver protease inhibitors locally, where they are naturally produced, and where they exert their homeostatic role. Local delivery would also decrease possible side effects of therapeutic intervention. To that aim, the use of genetically modified bacteria could constitute a major advance. Commensal or probiotic bacteria that colonise the gut can be genetically transformed to express human epithelium-derived protease inhibitors such as elafin or SLPI. Strong anti-inflammatory properties have been described in different animal models for such recombinant bacteria.22 ,78 Elafin delivered by recombinant lactic acid bacteria after oral administration in mice was detected in the colon lumen, as well as in the mucosal tissues. How this recombinant protein was able to cross the intestinal barrier: through passive diffusion in damaged epithelia or through active transport, is not clear yet. However, its presence was detected both in damaged areas and in areas where the epithelium was intact.22 Therefore, one can consider that protease inhibitor delivery through this approach might act both from the lumen and superficial mucosal tissues. Anti-inflammatory properties have also been demonstrated in cultured biopsy supernatants from patients with IBD.22 Treatments with bacteria recombinant for the expression of protease inhibitors were drastically more effective than treatments with bacteria recombinant for anti-inflammatory cytokines such as IL-10 or transforming growth factor-β. This is strongly in favour of targeting proteolytic activity for therapeutic options in IBD. However, the use of the recombinant bacteria strategy will have to consider the development of non-disseminating bacteria because of their genetically modified nature. Such development has already been described for other recombinant bacteria.79
Other interesting proteolytic targets for IBD treatment are the proteases from the ubiquitin–proteasome system.18 ,19 Polymorphisms on several genes of this system have been identified in patients with IBD, and pathogenic bacteria modify this system turnover.19 Proteasome inhibitors therapy targeting the ubiquitin–proteasome system, such as the use of bortezomib, which was successfully developed for cancer treatment, could constitute a new option to treat efficiently patients with IBD.
Protease inhibition as possible treatment for functional GI disorders
Proteases, through the activation of PARs, modify a number of physiological functions that are dysregulated in IBS. PAR2 activation causes visceral hypersensitivity, modifies intestinal motility and both PAR1 and PAR2 activation increase intestinal epithelial permeability.37 All these functions take an important part in IBS symptoms generation. In addition, increased trypsin-like activity (measured using a preferred trypsin substrate) has been demonstrated in tissues from patients with IBS.28 The increased activity was observed in all patient subgroups: diarrhoea-predominant, constipation-predominant or alternate-predominant, suggesting that protease activity might be a unifying feature of IBS. Further, several studies have reported that proteolytic activity released from tissues of patients with IBS provoke an increased permeability, and signal to extrinsic sensory neurons and intrinsic enteric neurons.80–83 This confirms the prominent effect of IBS-associated proteases on neuron signalling. Taken together, these studies highlight trypsin proteases as important molecular targets for IBS treatment.
Tryptase is another protease that is significantly increased in the mucosa of patients with IBS. Studies have demonstrated that enhanced tryptase activity is responsible for the increased permeability of rectal mucosa in diarrhoea-predominant patients.84 Tryptase inhibitors have been raised for mast cell-associated pathologies and may be tested in IBS, particularly on visceral hypersensitivity symptoms and increased permeability.
Both trypsin and tryptase have been shown to activate PAR2.85 In all animal studies investigating by which mechanisms tryptase, trypsin or IBS patient biopsy supernatants were causing increased permeability, neuron hyperexcitability or visceral hypersensitivity, proteases and/or PAR2 activation were identified as the principal mechanism of action.86 This suggests that PAR2 antagonism could constitute a valid therapeutic option for the treatment of IBS. However, a study investigating the effects of IBS patient biopsy supernatants on human submucosal or myenteric neurons preparations has determined that PAR1 rather than PAR2 was activated in human tissues.87 ,88 This suggests that in human, PAR1 antagonists should be considered for the treatment of IBS symptoms. However, the most recent advances in the pharmacology of PARs has taught us that PARs have several ways to signal other than calcium mobilisation usually measured.89 Adenylyl cyclase, MAPK and ERK signalling and β-arrestin recruitment would also have to be investigated in PAR2 response of human neurons, before ruling out a possible involvement of PAR2. In addition, the most striking effect of PAR2 activation was observed on visceral hypersensitivity symptoms and in sensory primary afferents, which might respond differently from submucosal or myenteric neurons. Therefore, for the time being, both PAR1 and PAR2 antagonism should still be considered as potential therapeutic options for IBS treatment.
Downstream from PAR activation (at least PAR2 and PAR4), mobilisation and potentiation of TRPV4 channel seem to be involved in the context of somatic mechanical hyperalgesia,90 and in the context of IBS.91–93 Most recently, a study has demonstrated that proteases, through the activation of PAR2, were able to induce the release of TRPV4 endogenous agonists, which were found upregulated in tissues from patients with IBS.94 Taken together, these data established the ion channel (TRPV4)-dependent mechanisms by which proteases influence neuronal signalling and visceral hypersensitivity in IBS.
A study that has investigated faecal proteases suggests that in diarrhoea-predominant patients with IBS, most of faecal protease activity is coming from the pancreas and is due to accelerated transit.30 Lowering transit time could therefore constitute a way in those patients to decrease luminal proteolytic activity and thereby the potential effects of this activity on microbiota composition or intestinal permeability.
Protease inhibition as possible treatment for colorectal cancer
As discussed above, numerous proteases are upregulated and potentially play a role in colorectal cancer. The identification of proteases that favour normal physiological functions instead of helping oncogenesis or tumour growth had most important clinical implications. The fact that proteases might have opposite effects in cancer might explain the failure of clinical trials that have used large spectrum protease inhibitors for treating patients with cancer.95 Furthermore, a significant number of proteases, and in particular intracellular proteases, have been defined as tumour suppression natural agents. Therefore, extreme caution is now associated with any antiprotease therapeutic strategy for cancer, and the inhibitory profile of antiprotease therapy is carefully evaluated according to the characteristics of the enzyme to be targeted, and its cellular source.
The ubiquitin–proteasome system is however the most protease-targeted system for cancer treatment. A number of bioactive compounds targeting E1, E2 enzymes and E3 ligase are now available for therapeutic tests96 and are currently under investigation.
The special case of coeliac disease
Coeliac disease is an autoimmune disorder of the small intestine, which involves an immune reaction to gluten non-degraded peptides such as gliadin. Strict and life-long gluten-free diet constitutes an effective treatment. However, therapies based on protease or antiprotease therapies have recently been suggested. First, the idea that assisted digestion to detoxify gluten by using microbial endopeptidases has been proposed.97 The use of microbial peptidase is necessary because no human enzyme exists to cleave at proline and glutamine sites, which are the most prominent sites in toxic gliadin peptides. This approach has numerous drawbacks, and in particular the fact that most of the enzymes used are inactivated in the stomach by pepsin and acidic pH. Rather than enabling patients to have a full gluten diet, protease therapy can protect patient with severe disease from unwanted or hidden exposure to gluten. In that case, proteases, but not protease inhibition, are considered as a therapeutic approach.
In contrast, a recent study proposes to use a protease inhibitor for coeliac disease treatment. In that study, the authors described that coeliac disease patients express lower amounts of the natural endogenous elastase inhibitor elafin.98 They further demonstrated that elafin inhibited the transformation of gliadin peptide into its immunogenic form. Finally, they demonstrated in a mouse model of coeliac disease that elafin delivery decreased inflammatory symptoms and enhanced barrier function. This study thus highlights the possible use of the protease inhibitor elafin as a therapeutic option for coeliac disease.
Beneficial effects of proteases in gut pathologies
Intestinal tissues demonstrate basal proteolytic activity in physiological conditions. Although very low compared with the activity detected in pathological tissues, the presence of low proteolytic activity in healthy tissues suggests that proteases can exert physiological functions in intestinal tissues and may even be protective. As discussed above, this has been clearly established for some MMPs that demonstrated antitumorigenic and anti-inflammatory properties.76 Surprisingly, some proteases such as chymotrypsin and neutrophil elastase seem to foster intestinal barrier function at least in vitro, increasing transepithelial resistance of intestinal epithelial cell monolayers.99 The authors demonstrated that this effect was independent of PAR activation. These findings could indeed suggest a protective role for some proteases in intestinal pathologies associated with a loss of intestinal barrier integrity. Additional anti-inflammatory effects for host or microbial proteases have been described along with their ability to degrade pro-inflammatory cytokines and chemokines.100–103 MMPs, microbial serine protease such as lactocepin or even cysteine proteases such as cathepsin B are among the proteases exerting such effect, which therefore protects from chronic inflammatory insults.100 ,101 ,103 Clearly, the physiological functions of proteases and their potential protective effects in gut pathologies have to be considered and taken into account, especially for therapeutic initiatives that would propose the use of protease inhibitors. However, more studies are necessary to define the spectrum of protective proteases and above all, the concentrations and conditions at which they might exert their protective effect.
Conclusion
Protease inhibition has definitively been raised in the recent years to the rank of ‘hot-topic’ for therapeutic strategies to treat GI diseases. Initially considered for cancer treatment, protease inhibition strategy has considerably evolved from strategies targeting large spectrum proteases, to strategies now targeting specific proteases. The evolution has also considered other indications than cancer. A very large amount of work has been performed in the domain of IBD and IBS, identifying new proteolytic targets (mostly extracellular proteases). New approaches, based on natural protease inhibitor delivery, and re-equilibration of specific proteolytic homeostasis have also been proposed and are considered as the most promising strategies in the near future. In the long term, there is a need to characterise the proteolytic profiles associated with each intestinal disease, or even within a same pathology, the proteolytic profile of patient's subgroups. Such definition will have to take into account only active proteases. To a given proteolytic profile, an adapted therapeutic strategy could then be proposed, targeting one or several proteases.
Acknowledgments
Thanks to Dr Celine Deraison and to Claire Rolland-Fourcade for realising and providing the in situ zymography pictures in figure 3.
References
Footnotes
Collaborators Celine Deraison, Claire Rolland-Fourcade.
Funding This work was supported by the Agence Nationale de la Recherche (R12177BB), the Region Midi-Pyrénées, by the European Research Council (ERC-2012-StG-20111109), the AFA (Association Francois Aupetit) and the AFER.
Competing interests None declared.
Provenance and peer review Commissioned; externally peer reviewed.