Article Text

Abstract
Objective Iron-containing micronutrient powders (MNPs) reduce anaemia in African infants, but the current high iron dose (12.5 mg/day) may decrease gut Bifidobacteriaceae and Lactobacillaceae, and increase enteropathogens, diarrhoea and respiratory tract infections (RTIs). We evaluated the efficacy and safety of a new MNP formula with prebiotic galacto-oligosaccharides (GOS) combined with a low dose (5 mg/day) of highly bioavailable iron.
Design In a 4-month, controlled, double-blind trial, we randomised Kenyan infants aged 6.5–9.5 months (n=155) to receive daily (1) a MNP without iron (control); (2) the identical MNP but with 5 mg iron (2.5 mg as sodium iron ethylenediaminetetraacetate and 2.5 mg as ferrous fumarate) (Fe group); or (3) the identical MNP as the Fe group but with 7.5 g GOS (FeGOS group).
Results Anaemia decreased by ≈50% in the Fe and FeGOS groups (p<0.001). Compared with the control or FeGOS group, in the Fe group there were (1) lower abundances of Bifidobacterium and Lactobacillus and higher abundances of Clostridiales (p<0.01); (2) higher abundances of virulence and toxin genes (VTGs) of pathogens (p<0.01); (3) higher plasma intestinal fatty acid-binding protein (a biomarker of enterocyte damage) (p<0.05); and (4) a higher incidence of treated RTIs (p<0.05). In contrast, there were no significant differences in these variables comparing the control and FeGOS groups, with the exception that the abundance of VTGs of all pathogens was significantly lower in the FeGOS group compared with the control and Fe groups (p<0.01).
Conclusion A MNP containing a low dose of highly bioavailable iron reduces anaemia, and the addition of GOS mitigates most of the adverse effects of iron on the gut microbiome and morbidity in African infants.
Trial registration number NCT02118402.
- diarrhea
- gut
- microbiome
- prebiotic
- enteric pathogens
- micronutrient powder
- anemia
- iron deficiency
- infancy
- kenya
Statistics from Altmetric.com
- diarrhea
- gut
- microbiome
- prebiotic
- enteric pathogens
- micronutrient powder
- anemia
- iron deficiency
- infancy
- kenya
Significance of this study
What is already known on this subject?
Iron-containing micronutrient powders (MNPs) are widely used in Sub-Saharan Africa to reduce anaemia.
Current MNPs contain a high daily iron dose (12.5 mg/day) that may decrease gut Bifidobacteriaceae and Lactobacillaceae, and increase enteropathogens, diarrhoea and respiratory tract infections (RTIs); thus, safer MNP formulations are needed.
Prebiotic galacto-oligosaccharides (GOS), commonly added to infant formula, can selectively enhance growth of Bifidobacteriaceae and Lactobacillaceae.
What are the new findings?
A new MNP formulation containing a low dose (5 mg/day) of highly bioavailable iron reduces anaemia in Kenyan infants.
The addition of 7.5 g GOS to this low-iron MNP mitigates most of the adverse effects of iron on the gut microbiome, resulting in greater abundances of Bifidobacterium and Lactobacillus, lower abundances of virulence and toxin genes of pathogens, less enterocyte damage, and a lower incidence of treated RTIs.
How might it impact on clinical practice in the foreseeable future?
Our findings demonstrate that the iron dose in MNPs can be reduced to 5 mg while maintaining efficacy, and that prebiotics given with low-iron MNPs in the African setting may be beneficial to reduce the adverse effects of iron on the infant gut.
Introduction
The WHO estimates that 62.3% of preschool children in Africa are anaemic,1 of which ≈50% is attributable to iron deficiency anaemia (IDA).2 Iron-containing micronutrient powders (MNPs) added to complementary foods effectively reduce IDA in infants, a strategy described as ‘in-home fortification’.3 However, current MNPs contain a high iron dose (12.5 mg) that is equivalent to the dose provided by oral iron supplements for the treatment of IDA in infancy.3 The safety of these MNPs is uncertain,4 5 as recent MNP trials (12.5 mg iron/day) in infants reported increases in diarrhoea and respiratory tract infections (RTIs).6 7 Iron absorption from MNPs by African infants is only 4%–9%8; thus, most of the iron passes unabsorbed into the infant colon. Iron is a growth-limiting nutrient for many gut bacteria,9 and for most enteric Gram-negative bacteria (eg, Salmonella, pathogenic Escherichia coli) iron acquisition is essential for virulence and colonisation.10 In contrast, lactobacilli and bifidobacteria can improve gut integrity and reduce colonisation by enteric pathogens,11 12 but lactobacilli require no iron13 and some Bifidobacterium spp can efficiently sequester iron, limiting its availability to pathogens.14 Large increases in colonic iron could favour pathogen growth, especially in African infants in areas of poor hygiene who have high carriage rates of pathogens.15 In a trial in Kenyan infants, MNPs increased the Enterobacteriaceae:Bifidobacteriaceae ratio, numbers of enteropathogenic E. coli, gut inflammation and diarrhoea.15
Thus, safer MNP formulations are needed. One strategy could be to reduce the iron dose but at the same time maximise absorption by providing iron as sodium iron ethylenediaminetetraacetate (NaFeEDTA), a chelated highly bioavailable iron compound, along with a phytase to reduce phytic acid in complementary foods, an inhibitor of iron absorption.16 A second strategy could be to include a component that potentially mitigates the adverse effects of fortification iron on the gut microbiome, such as a prebiotic, that could maintain and/or promote growth of the beneficial bacteria in the infant gut.12 17 Galacto-oligosaccharides (GOS) selectively enhance growth of Bifidobacteriaceae and Lactobacillaceae, and increase colonic production of short-chain fatty acids (SCFAs) that decrease luminal pH, which may reduce growth of enteropathogens.12 17 GOS has Generally Recognized as Safe (GRAS) status18 and is often added to commercial infant formula.12
We hypothesised that (1) a MNP formula containing 5 mg of highly bioavailable iron would reduce anaemia; and (2) the addition of GOS to a MNP containing 5 mg of iron would mitigate the adverse effects of iron on the infant gut microbiome and inflammation. Coprimary outcomes were (1) the gut microbiome assessed by 16S rDNA sequencing (the taxa of primary interest: Bifidobacterium, Lactobacillus, Enterobacteriaceae, Clostridiales and Bacteroidetes); (2) enteropathogen abundance assessed by quantitative PCR (qPCR) of virulence and toxin genes (VTGs); and (3) gut inflammation/integrity assessed by faecal calprotectin and plasma intestinal fatty acid-binding protein (I-FABP). Secondary outcomes were haemoglobin (Hb), iron status and morbidity.
Materials and methods
Study design
We conducted the study from October 2014 to December 2015 in southern coastal Kenya. Based on previous studies in African infants and children,15 19 we estimated a sample size of ≈35 infants in each group would be sufficient to detect a difference of approximately 0.85 log number of copies/g faeces in Lactobacillaceae and Bifidobacteriaceae, considering an SD of 1.25 in log abundance, with α=0.05 and β=0.8. We anticipated an attrition rate of ≈30% and planned to enrol 50 infants per group. The study was approved by the ethics and research committees of the Kenyatta National Hospital/University of Nairobi, Kenya (P521/10/2013) and the Zurich Cantonal Ethical Commission (2014–0232). Caregivers gave informed consent by either a written signature or a fingerprint. An independent Data Safety Monitoring Board (DSMB) monitored the study.
We prescreened 334 infants (study design, figure 1) for the following inclusion criteria: (1) age 6.5–9.5 months; (2) no chronic or acute illnesses; (3) no vitamin and mineral supplements in the previous 8 weeks; (4) no antibiotic treatment in the previous 10 weeks; and (5) anticipated residence in the area for the study duration. For 199 infants, we received informed consent and we enrolled them into a 2-week run-in period without MNPs to familiarise the infants and caregivers with preparation and consumption of the maize porridge; 171 infants completed the 2-week run-in period and were invited for the complete baseline screening. Inclusion criteria during the baseline screening were (1) Hb >70 g/L; and (2) Z scores for weight-for-age and weight-for-height both > −3. Finally, 155 infants met the inclusion criteria and were randomly assigned in consecutive order to the three intervention groups using a computer-generated list and three colour codes. The first (control group) received daily a MNP sachet containing 30 mg of ascorbic acid and other vitamins and minerals, 190 phytase unit (FTU) of a food-grade microbial phytase (Tolerase 20000G, DSM Nutritional Products, Kaiseraugst, Switzerland),16 and as filler, 10.5 g of maltodextrin. The second (Fe group) received the identical MNP but with 2.5 mg iron as NaFeEDTA and 2.5 mg iron as ferrous fumarate. The third (FeGOS group) received the identical MNP as the second group, except the maltodextrin was replaced with 10.5 g of 75% GOS (Vivinal GOS 75 Powder, FrieslandCampina, Wageningen, The Netherlands). GOS consists of chains of galactose units of varying length: the degree of polymerisation (DP) of the GOS used in this study (on weight percentage of oligosaccharide) was 31% DP2 (other than lactose), 38% DP3, 18% DP4, 8% DP5 and 5% DP6 and higher (FrieslandCampina). The composition of the MNPs is shown in table 1.
Composition of the three MNP formulations used in the study

Study design and participants flow chart. FeFum, ferrous fumarate; GOS, galacto-oligosaccharides; MNP, micronutrient powder; NaFeEDTA, sodium iron ethylenediaminetetraacetate.
The MNPs were packed in colour-coded sachets containing a daily dose, and the codes were kept by the DSMB; study participants and investigators were masked to group assignment. A baseline faecal sample and a venous blood sample were collected from the infants before the MNPs were distributed. Caregivers were carefully instructed on faecal sample collection and were given plastic diapers, spatulas and screw-cap plastic containers containing a carbon dioxide generator system to create an anaerobic atmosphere (Microbiology Anaerocult A mini, Merck, Darmstadt, Germany). The faecal samples were collected at home into the containers, and the study team aliquoted and stored them at −20°C the same day until further analysis. We measured infant weight and height, and recorded demographic characteristics, brief medical history and feeding habits of the infants using a questionnaire and local health records.
Weekly for 4 months, we dispensed seven MNP sachets and 2 kg of unfortified, refined maize flour to the families. During weekly visits, we assessed compliance by questioning the caregiver and collecting the previous week’s used and unused MNP sachets. At the weekly visits, we assessed infant morbidity over the previous week as described in the online supplementary material. After 3 weeks and 4 months of the intervention, we collected a faecal sample, and at 4 months we collected a venous blood sample and measured infant weight and length as described above. Any infants remaining anaemic at 4 months were examined and treated by the study clinicians according to local guidelines.
Laboratory methods
Blood samples
At baseline and 4 months, we collected a 3 mL venous blood sample. We measured Hb on the day of collection by using a HemoCue 300 analyser (HemoCue, Angelholm, Sweden) and separated plasma by centrifugation and froze it on the day of collection until analysis. Plasma ferritin (PF), soluble transferrin receptor (sTfR), C reactive protein and alpha-glycoprotein were analysed using a multiplex immunoassay,20 and I-FABP by using a commercial available ELISA (Hycult Biotech, Uden, The Netherlands). Reference values are given in the online supplementary material.
Faecal samples
We measured faecal calprotectin using the Calprest ELISA assay for stools, following the manufacturer’s procedures (Eurospital, Trieste, Italy) and faecal pH. Details of the method and reference values are given in the online supplementary material. We extracted and quantified faecal DNA as described in the online supplementary material. Using qPCR, we targeted VTGs of selected enteropathogenic bacteria: Bacillus cereus, Clostridium difficile, Clostridium perfringens, enterohemorrhagic E.coli (EHEC), enteropathogenic E.coli (EPEC) and enterotoxigenic E.coli (ETEC), Salmonella spp, and Staphylococcus aureus; details and primers are given in online supplementary table 1. These pathogens were chosen based on their common occurrence in Kenyan infants in our previous study.15 Barcoded amplicons from the V3–V4 region of 16S rRNA genes were generated using a two-step PCR, and universal primers appended with Illumina adapters were used for initial amplification of the V3–V4 part of the 16S rRNA gene. PCR products were purified and quantified, followed by multiplexing, clustering and sequencing on an Illumina MiSeq with the paired-end (2×) 300 bp protocol and indexing; details are given in the online supplementary material.
Statistical methods
We analysed biochemical and anthropometric data using the R statistical programming environment (R V.3.2.3, 2015). Normally distributed data were expressed as means and SD; non-normally distributed data as median and ranges or IQRs. When data were non-normally distributed, appropriate transformation of values was performed before statistical analysis. Details of the methods of comparison for the haematological, biochemical and microbiological data are given in the online supplementary material. Because we felt near significant p values should not be routinely ignored in this study, as some of these may provide interesting hints on the relationships of iron and GOS with the complex and still poorly understood infant gut microbiome, we mention p values <0.1 as statistical trends. We considered p values <0.05 as statistically significant.
Results
During the intervention, 10 infants dropped out (figure 1), and the final analysis was completed on 145 infants: control group (n=48), Fe group (n=49) and FeGOS group (n=48). At baseline, all infants were still partially breast fed except one infant in the Fe group, and all were already receiving complementary foods, mainly maize porridge; 97% of infants were still partially breast fed at endpoint. The mean±SD native iron concentration in the maize flour used for porridge preparation was 1.02±0.13 mg/100g dry matter. Compliance with the MNP sachets in the control, Fe and FeGOS groups was 94%, 95% and 94%, respectively.
Haematological and inflammation status
Table 2 shows, at baseline and 4 months, age, gender, iron and inflammation status. At baseline, there were no significant group differences. At 4 months, there were no significant group differences in any of the inflammation variables. At 4 months, compared with the control group, Hb and PF were higher in the Fe and FeGOS groups (p<0.001 for both); sTfR was lower in the Fe and FeGOS groups (p<0.001 for both); and the prevalence of anaemia and IDA was lower in the Fe and FeGOS group (p<0.001 for both). There were no significant differences between Fe and FeGOS at 4 months in any of these haematological variables. Online supplementary table 2 shows there were no significant differences in anthropometric variables between groups during the study.
Age, gender, iron and inflammation variables, faecal calprotectin and faecal pH, by group. Differences in these variables among Kenyan infants (n=155) at baseline and after 4 months of consuming daily a micronutrient powder containing either no iron (control), 5 mg of iron (Fe), or 5 mg of iron and 7.5 g GOS (FeGOS)
Gut microbiome by 16S rDNA sequencing
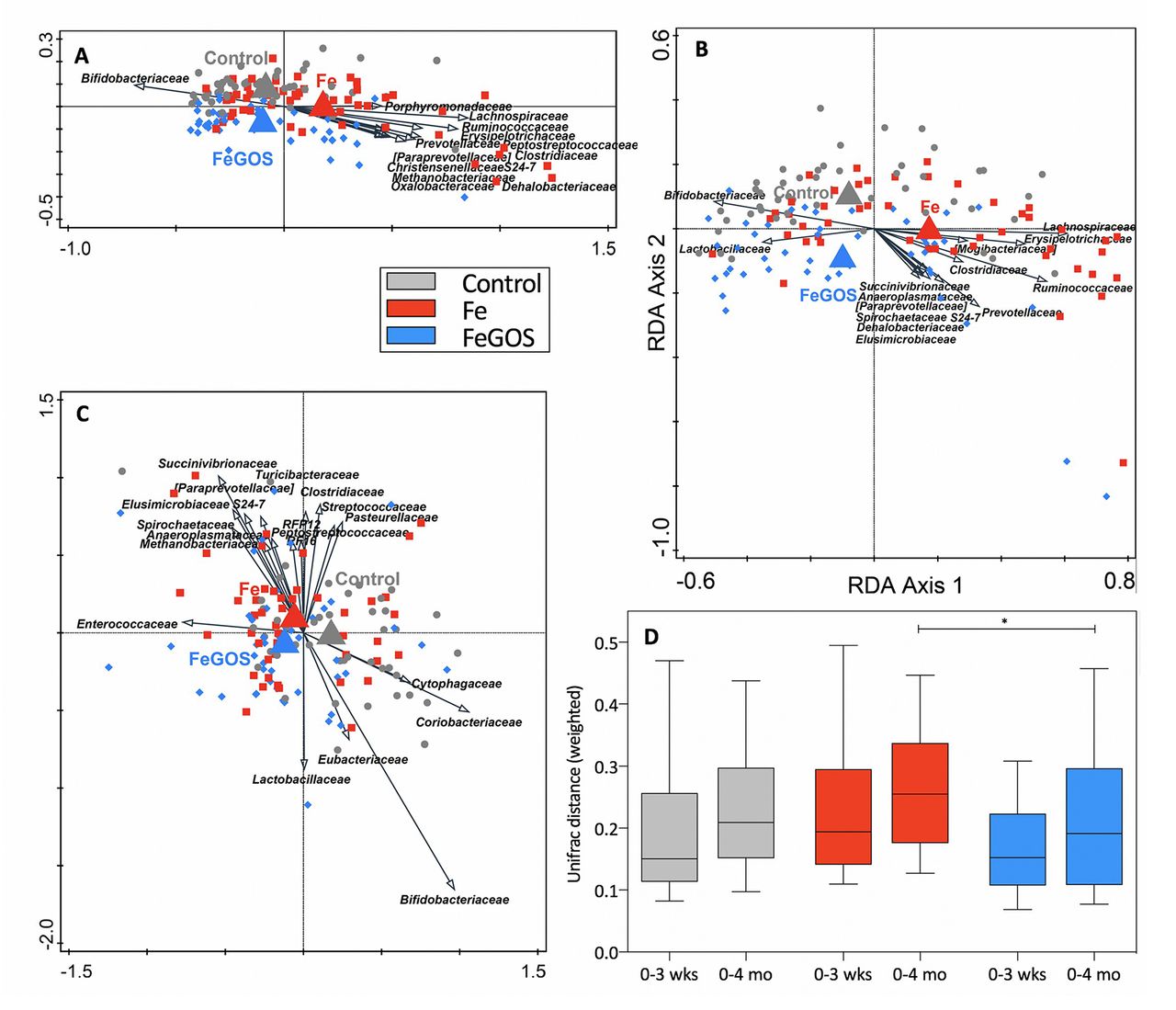
On average 23 902 bacterial 16S rDNA sequences per sample were analysed by 16S profiling. At baseline, the microbiome consisted of the phyla Actinobacteria (66.4% of the 16S rDNA reads), which was mostly represented by the family Bifidobacteriaceae, contributing 60.5% to the total 16S rDNA, Firmicutes (28.6%; including 4.6% Lactobacillus), Bacteroidetes (2.1%; including 0.6% Bacteroides and 1.3% Prevotella) and Proteobacteria (2.6%), with the predominant family Enterobacteriaceae (2.4%). In the redundancy analyses (RDAs) (figure 2A–C), the variation in the microbiota that could be explained by group at baseline was 0.5% (p=0.048). There was a treatment signal at 3 weeks and 4 months: variation explained by the group at 3 weeks increased to 0.7% (p=0.014) (figure 2A) and was 0.8% at 4 months (p=0.034) (figure 2B). In the ratio RDA of baseline to 4 months, the three groups were separated (trend, p=0.084; variation explained by treatment 0.7%) (figure 2C).

Redundancy analysis (RDA) and phylogenetic distances, by group. Kenyan infants (n=155) receiving daily a micronutrient powder containing either no iron (control); 5 mg of iron (Fe); or 5 mg of iron and 7.5 g of galacto-oligosaccharides (FeGOS): in the RDAs, the variation explained by group was (A) 0.7% at 3 weeks (p=0.014) and (B) 0.8% at 4 months (p=0.034); (C) the ratio RDA of baseline to 4 months showed separation of the three groups (variation explained by treatment 0.7%; trend, p=0.084). (D) The phylogenetic distance (UniFrac weighted index) from baseline to 3 weeks (0–3 weeks) and from baseline to 4 months (0–4 months) within each infant. The phylogenetic distance from 0 to 3 weeks and from 0 to 4 months among the intervention groups differed (trend, p=0.079 and p=0.055, respectively; Kruskal-Wallis test), and for 0–4 months was larger in Fe compared with FeGOS (trend, *p=0.063; Dunn’s post-hoc test).
Maturation of the infant gut microbiome during the study was reflected in the phylogenetic distance from baseline to 3 weeks and from baseline to 4 months within each individual, as shown in the weighted UniFrac distances (figure 2D). The phylogenetic distance from 0 to 3 weeks and from 0 to 4 months among the intervention groups differed (trend, p=0.079 and p=0.055, respectively), and for 0–4 months the phylogenetic distance in the Fe group was larger than in the FeGOS group (trend, p=0.063), while there was no significant difference in phylogenetic distance comparing the FeGOS and control groups. In the Fe group at 4 months, there was a shift towards a more mature, adult-like composition21 22 enriched in taxa such as Roseburia, Clostridium, Anaerostipes and Ruminococcus, while less mature, infant-associated taxa including Collinsella, Lactobacillus, Bifidobacterium and Megasphaera were associated with the control and FeGOS (see online supplementary figure 1).
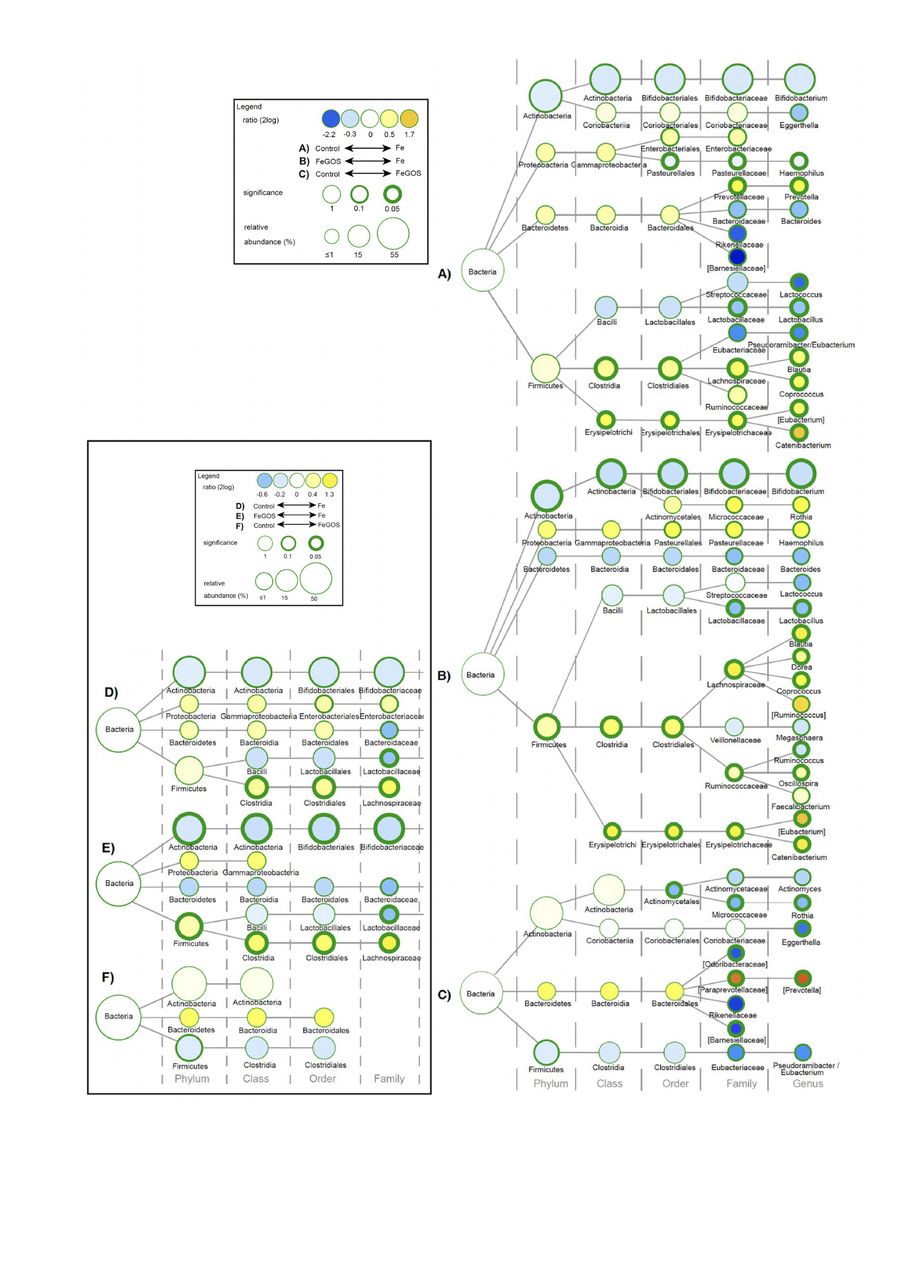
Between-group differences in all taxa at 3 weeks and 4 months are shown in figures 3A–C and 4A–C, respectively, and differences in taxa of our primary interest (Bifidobacterium, Lactobacillus, Enterobacteriaceae, Clostridiales and Bacteroidetes) at 3 weeks and 4 months are shown in the inset simplified figures 3D–F and 4D–F, respectively. Fe treatment had a major impact on the microbiome composition compared with FeGOS and control at 3 weeks and 4 months. At 4 months, compared with the control group, in the Fe group (figure 4D), there were lower abundances of the genera Lactobacillus (p=0.048), lower abundances of the genera Bifidobacterium (trend, p=0.058), higher abundances of the order Clostridiales (p=0.015) and higher abundances of the family Enterobacteriaceae (trend, p=0.086), but no significant differences in phylum Bacteroidetes. At 4 months, compared with the FeGOS group, in the Fe group (figure 4E), there were lower abundances of the genera Bifidobacterium (p=0.007) and Lactobacillus (p=0.006) and higher abundances of the order Clostridiales (p=0.001), but no significant difference in the abundance of Enterobacteriaceae and Bacteroidetes. In addition, at 4 months, Ruminococcaceae and Lachnospiraceae (mainly Dorea, Blautia and Coprococcus) and Erysipelotrichaceae were significantly more abundant in the Fe group compared with FeGOS and control (all, p<0.05). Remarkably, comparing the control and FeGOS groups, only a few low abundance taxa differed, and there were no significant differences in the abundances of Bifidobacterium, Lactobacillus, Enterobacteriaceae, Clostridiales and Bacteroidetes (figure 4F).

Group differences in gut microbial composition at 3 weeks. Differences in gut microbial composition in Kenyan infants (n=155) after 3 weeks of receiving daily a micronutrient powder containing either no iron (control); 5 mg of iron (Fe); or 5 mg of iron and 7.5 g of galacto-oligosaccharides (FeGOS): (A–C) All taxa. (D–F) Inset box shows only taxa that were prespecified targets of interest. Nodes represent taxa; edges link the different taxonomic levels. The node-size corresponds to the relative abundance (in %). The fold difference is calculated as the 2log of the ratio of the relative abundance between groups. Significance is expressed as the p value of a Mann–Whitney U test.

Group differences in gut microbial composition at 4 months. Differences in gut microbial composition in Kenyan infants (n=155) after 4 months of receiving daily a micronutrient powder containing either no iron (control); 5 mg of iron (Fe); or 5 mg of iron and 7.5 g of galacto-oligosaccharides (FeGOS). (A–C) All taxa. (D–F) Inset box shows only taxa that were prespecified targets of interest. Nodes represent taxa; edges link the different taxonomic levels. The node-size corresponds to the relative abundance (in %). The fold difference is calculated as the 2log of the ratio of the relative abundance between groups. Significance is expressed as the p value of a Mann–Whitney U test; exact p values for significance are in the text.
Occurrence and abundance of gut pathogens by qPCR
From all analysed stool samples at baseline (n=155), we detected C. perfringens in 65% (median in detected samples: 4.6 log gene copies/g faeces (IQR: 4.1–5.5)), C. difficile in 35% (5.6 (4.6–6.0)), S. aureus in 13% (4.7 (4.5–5.2)), B. cereus in 5% (4.7 (4.4–5.5)) and Salmonella spp in 4% (4.5 (4.3–5.2)). Further, we detected EPEC in 63% (5.5 (4.8–6.5)), the heat-labile enterotoxin (ETEC LT) and heat-stable enterotoxin (ETEC ST) of enterotoxigenic E. coli in 26% (6.6 (6.1–8.0)) and 21% (4.4 (3.8–5.7)), respectively, and Shiga toxin 2 (EHEC stx2) and Shiga toxin 1 (EHEC stx1) of enterohemorrhagic E. coli in 18% (4.3 (4.0–4.8)) and 8% (4.3 (3.9–4.9)), respectively. There were no significant differences among groups at baseline in any of these VTGs. At 4 months, in samples positive for EHEC stx2, there were lower abundances in the FeGOS group (4.0 (3.6–4.5)) compared with the Fe (4.8 (4.3–5.3)) and control groups (4.9 (4.7–5.3)) (p=0.036 and p=0.012, respectively). For the sum of the 10 VTGs of all pathogens, at 3 weeks there were lower abundances in the FeGOS group (median (IQR), 5.7 (4.9–6.5)) compared with both the control group (6.3 (5.6–7.1)) and the Fe group (6.4 (5.7–6.9)) (p=0.005 and p=0.007, respectively) (figure 5A). For the sum of VTGs of the pathogenic E. coli, at 3 weeks, there were lower abundances in the FeGOS group (5.8 (4.9–6.4)) compared with the control group (6.3 (5.5–7.1)) (p=0.035). At 4 months there were no significant differences in the VTGs of all pathogens or in the VTGs of pathogenic E. coli among groups. Within groups, from baseline to 4 months, there was a decrease in the VTGs of all pathogens in the FeGOS group (5.9 (5.3–6.7) to 5.4 (4.6–6.3)) (p=0.024) and in the control group (6.0 (5.2–6.7) to 5.4 (4.1–6.4)) (p=0.037), but no significant change in the Fe group. Correlations between enteropathogens and markers of gut and systemic inflammation are given in the online supplementary material.

Abundances of the sum of virulence and toxin genes of the 10 targeted pathogens and intestinal fatty acid-binding protein (I-FABP), by group. Kenyan infants (n=155) receiving daily a micronutrient powder containing either no iron (control); containing 5 mg of iron (Fe); or containing 5 mg of iron and 7.5 g of galacto-oligosaccharides (FeGOS). (A) The abundances of the sum of virulence and toxin genes of the 10 targeted pathogens (online supplementary table 1) at baseline, 3 weeks and 4 months, by group, *p<0.05, **p<0.01. Significance expressed as the p value of a Wilcoxon rank-sum test. (B) I-FABP at baseline and 4 months, by group, *p=0.0498. Significance expressed as the p value of analysis of covariance with baseline values as covariate. Boxes show the median and 25th and 75th percentiles; whiskers show the range; points show individual values.
Faecal calprotectin and pH, and plasma I-FABP
At 3 weeks and 4 months, there were no significant group differences in faecal calprotectin, although there was a non-significant ≈30% decrease from baseline to 3 weeks in the FeGOS group (table 2). At 3 weeks, faecal pH was higher in the Fe group than in the control and FeGOS groups (p=0.005 and p=0.001, respectively) (table 2). At 4 months, compared with the control group, I-FABP was higher in the Fe group (p=0.0498) (figure 5B).
Infant morbidity
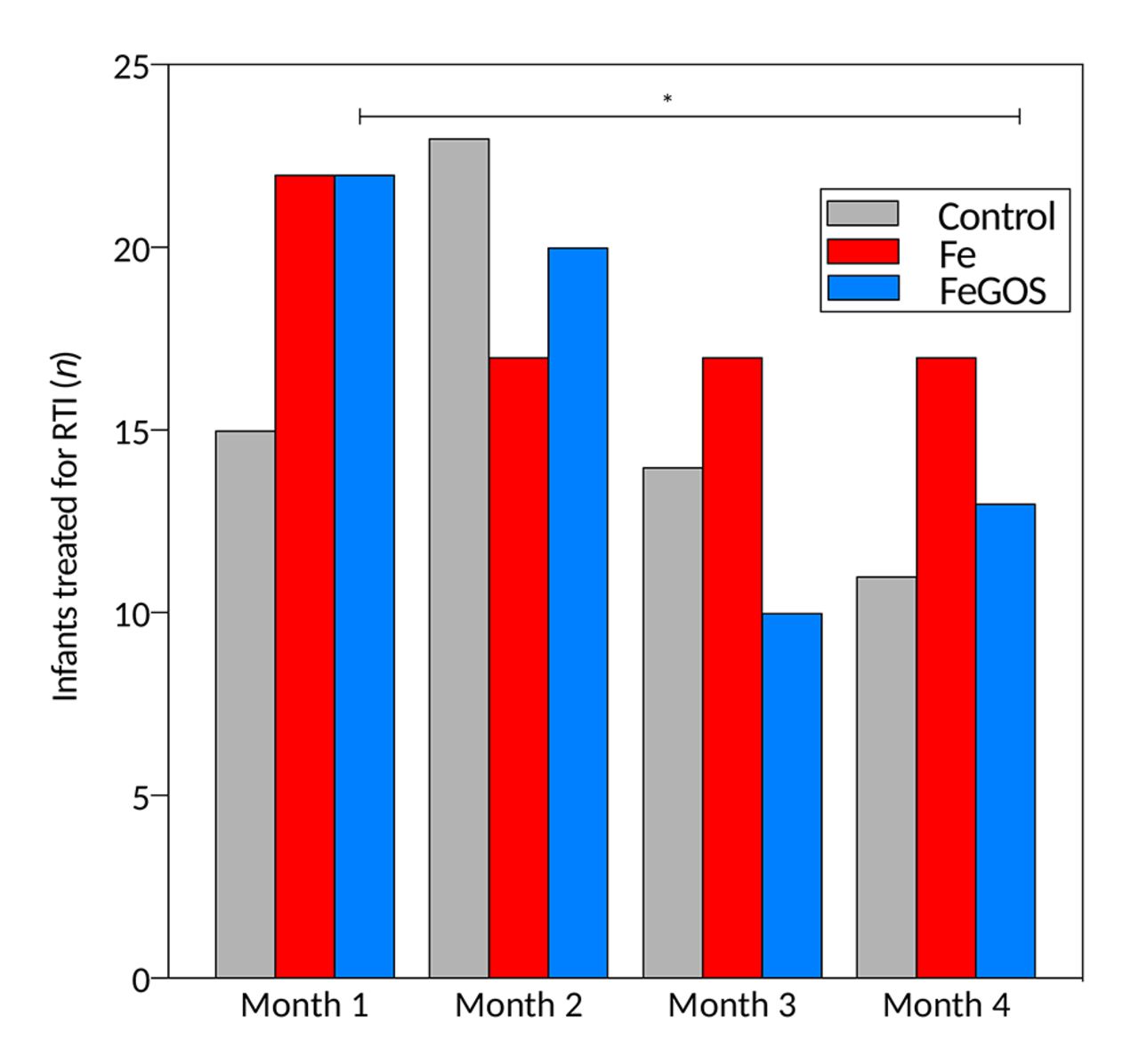
For morbidity assessed at the weekly visits, there were no significant time by treatment effects on diarrhoea or fever, but there was a significant time by treatment effect on the incidence of RTIs, with a decrease in RTIs over time in the FeGOS group (p=0.039). During the 4-month intervention, we recorded 465 treated cases in the health clinics: 237 for RTI, 74 for diarrhoea, 56 for malaria, 39 for skin infection and 59 for other health problems. There were no significant group differences in the number of infants treated for diarrhoea or other health problems during the intervention. However, more infants in the Fe group were treated for a RTI during the intervention (n=45; 87% of all infants) compared with the control group (n=38; 75%) (p=0.024) and compared with the FeGOS group (n=39; 75%) (trend, p=0.098). Over the 4 months of the study, there was a decrease in treated cases of RTIs in the FeGOS group (p=0.013) (figure 6) but no significant change in the other two groups.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Treated respiratory tract infections (RTIs), by group. Kenyan infants (n=155) receiving daily a micronutrient powder containing either no iron (control); containing 5 mg of iron (Fe); or containing 5 mg of iron and 7.5 g of galacto-oligosaccharides (FeGOS).*p=0.013. p value expresses the significant time effect of a generalised linear model with infants treated for RTI as response variable, time as fixed factor and subject as a random effect.
Discussion
Due to the potential adverse effects of unabsorbed iron on the infant gut,15 the iron dose in MNPs should be as low as possible while retaining efficacy against anaemia. In this study, daily use for 4 months of a MNP containing 5 mg iron as ferrous fumarate and NaFeEDTA was clearly efficacious in reducing anaemia and all measures of iron deficiency (table 2). The haematological response comparing Fe and FeGOS was comparable, suggesting the addition of the GOS did not affect iron bioavailability. This 5 mg dose is 60% lower than the 12.5 mg in currently used MNPs, yet the improvements in Hb (≈+5–9 g/L) and reductions in anaemia (≈−30%–50%) compare favourably with those reported for 12.5 mg iron MNPs.3
Previous intervention studies of the effect of iron on the infant and child microbiome are limited.23 In a controlled trial in 6-month-old Kenyan infants, iron-containing MNPs increased the Enterobacteriaceae:Bifidobacteriaceae ratio and numbers of enteropathogenic E. coli.15 In contrast, in 6-month-old Malawian infants randomised to no intervention or dietary supplements providing 5.5–6.0 mg iron/day, there were no significant differences in the gut microbiota after 12 months.24 In a controlled trial in Ivorian school-aged children receiving iron-fortified biscuits (≈9 mg iron/day), iron increased Enterobacteriaceae and decreased Lactobacillaceae.19 In contrast, in South African school-aged children from an area with better hygiene, iron supplements (50 mg/day, 4 days/week) caused no discernible effects on the gut microbiota.25 The varied findings from these studies are likely due to age-related differences in the gut microbiota between infants and older children, differences in methods used to characterise the gut microbiome, differences in geographical setting and hygiene and/or the iron compound and dose given.
In our study, addition of GOS to an iron-containing MNP counterbalanced the effects of iron: after 4 months of the intervention in the Fe group, there were lower relative abundances of Bifidobacteriaceae and Lactobacillaceae, and higher relative abundances of Clostridiales, compared with the control and FeGOS groups; in contrast, there were no significant differences in these taxa comparing the FeGOS group with the control group (Figure 4F). We chose a GOS dose of 7.5 g/day based on studies that reported a bifidogenic effect in infants and adults at this dose and because this is the typical daily dose delivered by GOS-containing commercial infant formulas.12 17 The enhancement of commensal bacteria strengthens the barrier effect against enteropathogen growth: commensal bacteria occupy more microbiological niches, compete for nutrients, increase secretory IgA and reduce pathogen adhesion site.12 17 The suppressive effect on enteropathogens was visible in our data: during the intervention, the FeGOS group had lower abundances of the VTGs of all pathogens (figure 5A) and the VTGs of pathogenic E. coli compared with the Fe and control groups. Prebiotics also increase commensal production of SCFAs that decrease luminal pH, which may reduce growth of pathogens,26 27 and in our study, at 3 weeks, faecal pH was lower in the FeGOS group than in the Fe group (table 2).
Previous studies in African infants15 and children19 have shown that iron fortification increases faecal calprotectin. Faecal calprotectin is a non-specific marker of gut inflammation that reflects neutrophil infiltration of the mucosa28 29; levels are normally high in early infancy and decrease over the first 2 years as the gut matures.15 In contrast to our previous MNP trial in Kenyan infants where a 12.5 mg iron dose increased faecal calprotectin, the 5 mg dose used here (and a 2.5 mg dose used previously15) did not increase faecal calprotectin (table 2), suggesting that daily doses of ≤5 mg iron may not trigger gut inflammation in this setting. Damaged enterocytes release I-FABP into the bloodstream30; it is a sensitive marker for enterocyte injury and can be combined with faecal calprotectin to assess disease severity.31 I-FABP in our infants at baseline was higher than in European children with sepsis,32 and high plasma I-FABP in African infants may reflect enterocyte injury due to common enteric infections.33 In our study, at 4 months, I-FABP was higher in the Fe group than in the control group, while there was no significant difference comparing the FeGOS group with control (figure 5B), suggesting the addition of GOS may have reduced Fe-mediated enterocyte damage; this effect may be mediated through changes in gut microbiota34 and/or iron-induced oxidative stress to enterocytes.35
Compared with the Fe group, there was a lower incidence of RTIs in the FeGOS group and a decrease in RTIs in the FeGOS group over the 4-month intervention (figure 6), but no group differences in diarrhoea. In our previous MNP study in Kenyan infants, there was a non-significant increase in incidence of diarrhoea with the 12.5 mg Fe dose, but no increase in RTIs.15 This difference might be explained by the lower dose of iron in the present study, and/or because the infants in the present study were ≈2 months older at entry. In a large MNP study (12.5 mg iron/day) that systematically collected morbidity data in Pakistani infants aged 6–18 months, there was an increased incidence of RTIs and diarrhoea with iron.7 Controlled trials in industrialised countries have reported that GOS alone or in combination with other prebiotics and probiotics reduced RTIs in infants.36 37 Possible mechanisms of how prebiotics and probiotics reduce RTIs include reduced pathogen colonisation in the respiratory epithelium,38 regulation of mucosal immunity through activation of inflammasomes and modulation of the systemic immune response.39
Strengths of our study include (1) its double-blind, randomised controlled design; (2) a study population with high carriage rates of enteropathogens; (3) low attrition rate (≈6%) during the study; and (4) extensive characterisation of the gut microbiome using the complementary methods of sequencing and qPCR for selected target species. We used maltodextrin in the control and Fe groups as a neutral comparator to the GOS because glucose from maltodextrin is rapidly absorbed in the proximal small intestine,40 so its presence is unlikely to have influenced our comparisons. A limitation of our study is that despite randomisation, there were small baseline differences in gut microbiota composition among the groups, which may have affected our comparisons. Also, because of the small sample size and high between-subject variability for the gut microbiota, a beta error may have contributed to non-significant comparisons. In addition, we characterised the gut microbiome in faecal samples, which may not necessarily represent bacterial abundances in the distal small intestine, caecum and proximal colon.40
In conclusion, a MNP containing a 5 mg daily dose of highly bioavailable iron is efficacious in reducing IDA in African infants, and this low dose is likely safer in that it induces less adverse changes in the gut microbiome and no increase in faecal calprotectin compared with a 12.5 mg iron dose.15 Our findings suggest that prebiotics given with iron-containing MNPs in the African setting may be beneficial to reduce the adverse effects of iron on the infant gut microbiome. Expert groups have judged prebiotics to be safe and well tolerated in infants18; however, nearly all previous studies were done in infants from high-income countries with good hygiene. Further studies confirming the safety and tolerability of GOS in the African setting are needed, as well as the smallest effective dose of GOS needed to offset the adverse effects of iron on the infant gut. In addition, studies combining GOS with iron fortificants or supplements in other geographical settings (eg, South Asia) and in older children would be valuable.
Acknowledgments
We thank the families who participated in this study, the field workers and nurses from Msambweni County Referral Hospital (Msambweni, Kenya) and Kikoneni Health Centre (Kikoneni, Kenya), as well as C Zeder, A Krzystek, V Galetti, A Die, A Minder and S Kobel (ETH Zurich, Switzerland) and J Erhardt (Willstaett, Germany) for support of the laboratory and data analysis. This study is registered at ClinicalTrials.gov as NCT02118402.
References
Footnotes
Contributors DP, CIC, DM, TB-J, CL and MBZ designed the study; DP, MAU and CIC conducted the study; DP, GAMK, JB, HMT and MBZ analysed the data and performed the statistical analyses; DP, MAU, CIC, GAMK, DM, TB-J, CS, JB, HMT, CL, SK and MBZ participated in the data interpretation; DP, GAMK, CS and MBZ wrote the manuscript. All authors edited the manuscript. All authors read and approved the final version of the manuscript.
Funding ETH Global and the Sawiris Foundation for Social Development, ETH Zurich, Switzerland; DSM Nutritional Products, Kaiseraugst, Switzerland. Sight and Life (Kaiseraugst, Switzerland) donated the MNPs used in this study. FrieslandCampina (Wageningen, The Netherlands) donated the GOS used in this study.
Ethics approval Kenyatta National Hospital/University of Nairobi, Kenya (P521/10/2013) and the Zurich Cantonal Ethical Commission (2014-0232).
Provenance and peer review Not commissioned; externally peer reviewed.