Article Text

Abstract
Background The immune contexture predicts prognosis in human colorectal cancer (CRC). Whereas tumour-infiltrating CD8+ T cells and myeloid CD16+ myeloperoxidase (MPO)+ cells are associated with favourable clinical outcome, interleukin (IL)-17-producing cells have been reported to correlate with severe prognosis. However, their phenotypes and functions continue to be debated.
Objective To investigate clinical relevance, phenotypes and functional features of CRC-infiltrating, IL-17-producing cells.
Methods IL-17 staining was performed by immunohistochemistry on a tissue microarray including 1148 CRCs. Phenotypes of IL-17-producing cells were evaluated by flow cytometry on cell suspensions obtained by enzymatic digestion of clinical specimens. Functions of CRC-isolated, IL-17-producing cells were assessed by in vitro and in vivo experiments.
Results IL-17+ infiltrates were not themselves predictive of an unfavourable clinical outcome, but correlated with infiltration by CD8+ T cells and CD16+ MPO+ neutrophils. Ex vivo analysis showed that tumour-infiltrating IL-17+ cells mostly consist of CD4+ T helper 17 (Th17) cells with multifaceted properties. Indeed, owing to IL-17 secretion, CRC-derived Th17 triggered the release of protumorigenic factors by tumour and tumour-associated stroma. However, on the other hand, they favoured recruitment of beneficial neutrophils through IL-8 secretion and, most importantly, they drove highly cytotoxic CCR5+CCR6+CD8+ T cells into tumour tissue, through CCL5 and CCL20 release. Consistent with these findings, the presence of intraepithelial, but not of stromal Th17 cells, positively correlated with improved survival.
Conclusions Our study shows the dual role played by tumour-infiltrating Th17 in CRC, thus advising caution when developing new IL-17/Th17 targeted treatments.
- COLORECTAL CANCER
- T LYMPHOCYTES
- CANCER IMMUNOBIOLOGY
- IMMUNE RESPONSE
- INFLAMMATORY MEDIATORS
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Infiltration of colorectal cancers (CRCs) by defined populations of immune cells predicts clinical outcome irrespective of tumour stage.
CRC-infiltrating CD8+ T cells and CD16+ myeloperoxidase (MPO)+ neutrophils have been found to be associated with prolonged survival, whereas infiltration by interleukin (IL)-17-producing cells, as evaluated in a limited number of cases, has been suggested to correlate with more severe prognosis.
IL-17 is a proinflammatory cytokine mediating protumorigenic and proangiogenic effects.
Monoclonal antibodies targeting IL-17/IL-17-receptor or impairing expansion of IL-17-producing cells may represent a new therapeutic option in CRC.
What are the new findings?
Analysis of a large cohort of CRCs shows that tumour-infiltrating IL-17-producing cells are not themselves predictive of poor clinical outcome.
Intraepithelial localisation of CRC-infiltrating IL-17+ cells is associated with improved survival.
CRC infiltration by IL-17+ cells correlates with the presence of beneficial CD8+ T cells and CD16+ MPO+ neutrophils.
CRC-infiltrating IL-17+ cells, mostly consisting of polyfunctional T helper 17 cells (Th17), can recruit highly cytotoxic CD8+ T cells into tumour nests through CCL5 and CCL20 release.
How might it impact on clinical practice in the foreseeable future?
By disclosing the dual role played by CRC-Th17, our findings question therapeutic approaches aimed at inhibiting Th17 development or expansion, possibly resulting in impaired tumour infiltration by beneficial effector cells. The positive contribution of Th17 to anti-tumour immune responses should not be disregarded when developing new IL-17/Th17 targeted treatments in CRC.
Introduction
The tumour immune contexture—that is, type, location, density and functional orientation of tumour-infiltrating immune cells,1 predicts clinical outcome in human colorectal cancer (CRC). In particular, CD45RO+ memory T lymphocytes, cytotoxic CD8+ T cells (CTLs) and interferon (IFN)-γ-producing T helper 1 cells (Th1) have been found to be associated with prolonged survival, irrespective of tumour stage (5–7). Unexpectedly, Foxp3+ regulatory T cells (Tregs),2 ,3 CD16+ and myeloperoxidase (MPO)+ myeloid cells,4–6 also correlate with favourable clinical outcome. In contrast, tumour infiltration by interleukin (IL)-17A-producing cells, evaluated so far in a limited number of cases (50–200), appears to be associated with unfavourable prognosis.7 ,8
IL-17A (hereafter referred to as IL-17) is an inflammatory cytokine, secreted by different cell types, including CD4+ T helper cells (Th17),9 ,10 CTLs, γδT cells, Tregs,11–13 natural killer (NK) cells, NKT cells, lymphoid tissue inducer (LTi)-like cells and neutrophils.14 ,15 IL-17 plays a prominent role in protective immune responses against bacterial and fungal infections and in the pathogenesis of inflammatory disorders.9 ,10 ,16
Experimental models indicate that IL-17 promotes intestinal tumorigenesis,17–22 either by favouring proliferation of aberrant epithelial cells21 or by inducing IL-6 release by tumour-associated stroma.18 Furthermore, IL-17 promotes angiogenesis through vascular endothelial growth factor (VEGF) production,18 ,20 thus mediating tumour resistance to antiangiogenic treatments.18
Monoclonal antibodies targeting IL-17/IL-17-receptor, or cytokines, such as IL-23, supporting Th17 development, have been recently developed and their clinical application in several inflammatory and autoimmune diseases is being evaluated.23–25 These reagents may provide a new therapeutic option in CRC.26 However, before testing IL-17/Th17-targeted treatments, a more comprehensive analysis of CRC-infiltrating, IL-17-producing cells is required.27
We evaluated the prognostic significance of IL-17 in a tissue micro-array (TMA) including 1148 CRC cases and we investigated phenotypes and functions of CRC-derived IL-17+ cells. Here we show that CRC-infiltrating IL-17-producing cells, mainly consisting of polyfunctional Th17, do not themselves predict clinical outcome, but rather play a dual role. On the one hand, owing to IL-17 secretion, they favour release of protumorigenic factors by tumour and tumour-associated stroma. However, on the other hand, they promote recruitment of beneficial neutrophils and CTLs by secreting specific chemokine and cytokine patterns. Interestingly, the presence of intraepithelial Th17 was significantly associated with patient survival, consistent with the ability of Th17 to drive beneficial immune cells into the tumour. The potential contribution of tumour-infiltrating Th17 to anti-tumour immune responses should not be disregarded when considering new IL-17/Th17 targeted treatments.
Materials and methods
Immunohistochemistry
A previously described TMA, including 1420 non-consecutive primary CRCs and 71 normal colonic mucosa samples,2 ,4–6 ,28 was stained with a goat polyclonal anti-human IL-17 antibody (R&D Systems; staining I). IL-17 expression was evaluable in 1151 CRCs and 39 healthy mucosa samples. A randomised subgroup, including 746 CRCs and 27 healthy mucosa cases, was stained with a rabbit polyclonal anti-human IL-17 antibody (H-132, Santa Cruz Biotechnology; staining II). Secondary stainings and negative controls were performed as described.29
Protein markers were scored by three observers (AL, FA and FT). Cases were classified in four groups, according to numbers of positive cells/punch (0, 1–10, 11–50, >50). Staining protocols for CD8, CD16 and MPO have been previously reported.4 ,6 ,28 Clinical information (see online supplementary table S1) was retrieved from patient records. The use of this information was approved by local ethical authorities.
Clinical specimen collection and processing
Clinical specimens were collected from consenting patients undergoing surgical treatment at Basel University Hospital, St Claraspital Basel, Kantonsspital Olten, Kantonsspital St Gallen and Ospedale Civico Lugano. Tumour and corresponding tumour-free mucosa fragments were snap-frozen for RNA extraction or enzymatically digested (2 mg/mL collagenase IV, Worthington Biochemical Corporation and 0.2 mg/mL DNAse I, Sigma-Aldrich, for 1 h at 37°C) to obtain single cell suspensions.
Flow cytometry and cell sorting
Cell suspensions from CRCs and tumour-free mucosa, and peripheral blood mononuclear cells (PBMC) of healthy donors (HDs) or patients with CRC, were incubated with 50 ng/mL phorbol 12-myristate 13-acetate, 1 μg/mL ionomycin and 5 μg/mL brefeldin A (Sigma-Aldrich) for 5 h. Cells were fixed and surface stained with fluorochrome-conjugated antibodies specific for human CD3, CD4, CD8, CD45RO, CD56, CD127, CCR6, HLA-DR, γδTCR (all from BD Biosciences), V α 24 J α 18 TCR (eBioscience) and CD66b (BioLegend). Intracellular staining was then performed with antibodies specific for human IL-17 (eBioscience), IFN-γ, tumour necrosis factor (TNF)-α, IL-22, IL-21, IL-8 (all from BD Biosciences) or granulocyte-macrophage colony stimulating factor (GM-CSF) and Foxp3 (BioLegend).
Chemokine receptor expression on CD8+ T cells was evaluated in CRC samples and autologous PBMC by surface staining with anti-human CCR5, CCR6 and CXCR3 antibodies (BD Biosciences). Stained cells were analysed by FACSCalibur flow cytometer (BD Biosciences) and FlowJo software (Tree Star).
Tumour, endothelial and mesenchymal cells were sorted from CRC cell suspensions using a BD Influx (BD Biosciences), upon staining with EpCAM-, CD31- and CD90-specific antibodies (BD Biosciences), respectively.
T cell expansion and cloning
CRC-isolated T lymphocytes were stimulated with 1 μg/mL of phytohaemagglutinin (Sigma-Aldrich) and expanded in medium supplemented with 100 IU/mL IL-2 (Roche Applied Science) and 5% of pooled human AB serum (provided by Blutspendenzntrum Beider Basel, Basel University Hospital) for 20 days. Th17 cells, identified based on CD4+ CXCR3-CCR4+CCR6+ phenotype,30 were sorted by flow cytometry and further expanded as bulk populations. Th17 clones (hereafter referred to as CRC-Th17) were subsequently generated from bulk populations by limiting dilution. Supernatants from Th17 bulk populations and clones were obtained by T cell activation with plate bound anti-CD3 (10 μg/mL, clone UCHT1, eBioscience) and soluble anti-CD28 antibodies (1 μg/mL, clone CD28.2, BD Biosciences). After overnight culture, supernatants were collected and used for determination of cytokine contents by ELISA and for migration assays.
Real-time reverse-transcription PCR
Total RNA was extracted from tissues or sorted cells using the RNeasy Mini Kit protocol (Qiagen), treated with DNAse I (Invitrogen) and reverse transcribed using the Moloney murine leukemia virus reverse transcriptase (M-MLV RT, Invitrogen). Quantitative real-time PCR was performed in the ABI prism 7700 sequence detection system, using TaqMan Universal Master Mix and No AmpErase UNG (both from Applied Biosystems). Commercially available primers and probes specific for human IL-17A (Hs99999082_m1), CCL5 (Hs00982282_m1), CXCL9 (Hs00171065_m1) and CXCL10 (Hs99999049_m1, all from Applied Biosystems) were used.
Cell lines
Human CRC cell lines LS180, COLO205 and HT29, purchased from the European Collection of Cell Cultures, were maintained in RPMI 1640 (GIBCO) or, for HT29, McCoy's 5A medium (Sigma-Aldrich), supplemented with 10% fetal bovine serum, GlutaMAX-I and kanamycin (GIBCO). HMEC cells (provided by Professor T Resink, University of Basel) were cultured in EBM-2 medium (Lonza). Tumour-associated stromal cells (TASC) were expanded from CRC samples in α-MEM (GIBCO) supplemented with 10% fetal bovine serum and 5 ng/mL FGF-2 (R&D Systems).
In vitro migration assay
Migration of neutrophils, isolated from PBMC of HDs by magnetic beads (EasyStep enrichment kit, StemCell Technologies), was assessed in transwell plates (5 µm pore size, Corning Costar), towards Th17 supernatants or recombinant cytokines (IL-17, R&D Systems, 50 ng/mL; IL-8, R&D Systems, 100 ng/mL; GM-CSF, R&D Systems, 100 ng/mL), for 90 min at 37°C. In specific experiments, anti-IL-8 or anti-GM-CSF antibodies (10 μg/mL, R&D Systems) were added to Th17 supernatants.
Migration of CD8+ T cells, sorted from PBMC of HDs by magnetic microbeads (Miltenyi Biotec) and pre-activated overnight with anti-CD3/CD28 antibodies, was evaluated in transwell plates (5 µm pore size) towards Th17 supernatants, supernatants of HMEC cells, untreated, or exposed overnight to Th17 supernatants or recombinant proteins (IL-17, 50 ng/mL; TNF-α, 1 ng/mL, R&D Systems; IFN-γ, 1 ng/mL, Biolegend; or their combination), or towards CCL5 (60 and 200 ng/mL, R&D Systems) and/or CCL20 (300 and 1000 ng/mL, R&D Systems). Depletion of CCL5 and/or CCL20 from Th17-derived supernatants was obtained by specific capture antibodies (R&D Systems). Cell migration was quantified by flow cytometry.
ELISA
Cytokine/chemokine contents in supernatants were assessed by ELISA using CCL5, CCL20, CXCL10 and VEGF DuoSet ELISA (all from R&D Systems) and IL-6-specific reagents (BD Biosciences).
T cell migration into engineered tumours
Tridimensional tumour tissues were engineered in a previously described bioreactor system.31 HT29 cells (1×106) were injected and perfused through a collagen scaffold (Ultrafoam Avitene Collagen Hemostat, Davol Inc). At day 7, 5×106 CRC-Th17 cells were injected and, after overnight incubation, were activated by CytoStim (Miltenyi Biotec) for 3 h. After extensive washing, perfusion was stopped and CD8+CD45RO+, sorted from PBMC of HDs, were then injected and allowed to spontaneously migrate overnight. Tissues were enzymatically digested and analysed by flow cytometry. Additionally, paraffin-embedded or cryosections were collected for H&E staining and immunofluorescence analysis.
Histological and immunofluorescence analysis
Paraffin sections (5 μm) were stained with H&E and analysed by light microscopy. Cryosections (10 μm) were incubated with rabbit polyclonal anti-human CD8 (Abcam) and mouse monoclonal anti-human CD4 (DakoCytomation) antibodies, or with rabbit polyclonal anti-human IL-17 (H-132, Santa Cruz Biotechnology) and mouse monoclonal EpCAM (Cell Signalling), followed by secondary species-specific Alexa Fluor 488- or Alexa Fluor 547-conjugated antibodies (Invitrogen). Nuclei were counterstained with 4,6-diamidino-2-phenylindole. Sections were examined under an Olympus BX61 fluorescence microscope (Olympus) and images captured with 10× and 20× magnifications using a F-VIEW II camera (Olympus) and AnalySIS software (Soft Imaging System GmbH).
In vivo migration assay
LS180 cells were inoculated subcutaneously in 8-week-old NSG mice (Charles River, Germany). CRC-Th17 and CD8+ T cells, isolated from PBMC of HDs, were activated overnight with anti-CD3/CD28 antibodies. Carboxyfluorescein succinimidyl ester-labelled CD8+ T cells were adoptively transferred by intravenous injection in tumour-bearing mice (maximum tumour volume 1 cm3) alone or together with Th17 (5×106 cells/subset/mouse). After 48 h, tumours were harvested and frequencies of CD8+CSFE+ T cells in tumour cell suspensions were evaluated by flow cytometry.
Statistical analysis
CRC cases were classified into four categories according to numbers of IL-17+ cells/punch (0; 1–10; 11–50; >50). Specific cut-off values for CD8, CD16 and MPO (10, 50 and 60, respectively) were obtained by receiver operating curve (ROC) analysis.4 ,6 ,28
A χ2 test was used to determine the significance of differences between dichotomous variables. Survival analysis was depicted by Kaplan–Meier method and compared with log-rank test. Statistical analyses were performed using R (V.2.15.2, http://www.r-project.org).
Differences in frequencies of IL-17+ cells within PBMC of HDs or patients with CRC and between tumour and control tissues were evaluated by t test. Differences in migration rates and cytokine release were tested by one way analysis of variance using GraphPad Prism5 (GraphPad Software).
Results
CRC-infiltrating IL-17+ cells do not predict clinical outcome
IL-17+ cells were evaluated by immunohistochemistry upon staining of a characterised TMA (38–39 and see online supplementary table S1) with two different polyclonal anti-IL-17 antibodies (staining I, figure 1A and staining II, see online supplementary figure S1A). Results obtained from the two stainings showed good and significant correlation (r=0.436 p<0.00001, weighted Cohen κ=0.284). IL-17+ cells were detectable within epithelial and stromal compartments. Numbers of intraepithelial IL-17+ cells were significantly higher in tumour samples than in normal colonic mucosa (figure 1Β). Accordingly, higher IL-17 mRNA levels in CRC than in corresponding control tissues were detected (see online supplementary figure S1B).

Tumour infiltration by interleukin-17 (IL-17)-producing cells does not predict survival in colorectal cancer (CRC). IL-17 expression was evaluated by immunohistochemistry on a tissue microarray (TMA) including 1151 cases of primary CRC. (A) Representative pictures of IL-17 staining I (see ‘Materials and methods’). Numbers of IL-17+ cells per punch are indicated. (B) Distribution of IL-17+ cells within the epithelial or stromal fraction of healthy colonic tissues (Ctr) or tumour samples (T). Statistical significance was assessed by χ2 test. (C) Kaplan–Meier curves illustrating overall survival (OS, left panel) and relapse-free survival (RFS, right panel) probability according to IL-17+ cell density. Numbers of deaths/total cases within each category are indicated. Statistical significance was assessed by log-rank test. (D) Numbers of IL-17+ cells within CRC cases characterised by low or high infiltration of CD16+, MPO+ and CD8+ cells, according to cut-off scores identified by receiver operating characteristic curve analysis, as described in ‘Materials and methods’. Statistical significance was assessed by χ2 test. MPO, myeloperoxidase.
No association between IL-17+ infiltrates and tumour location, mismatch repair status or tumour border configuration was seen (table 1). In a limited group of cases for which additional clinical information was available, no correlation between numbers of IL-17+ cells and local recurrences (n=446), or distant metastasis (n=452) was seen (table 1). In contrast, IL-17 + cells strongly correlated with peritumoral lymphocytic infiltration (p<0.001). A slight increase in IL-17+ cell numbers was seen in tumours characterised by early T and N stage, low grade and absence of vascular invasion. Unexpectedly, no significant impact of IL-17+ cells on overall survival (figure 1C left panel, n=1151 and see online supplementary figure S1C, n=649) or relapse-free survival (figure 1C right panel, n=443) was seen.
Association of IL-17 lymphocyte count and clinicopathological features in CRC (n=1148)
Tumour infiltration by IL-17+ cells is associated with the presence of CTLs and neutrophils
Interestingly, IL-17 was found to correlate significantly with CD16, MPO and CD8 markers (figure 1D), predictive of improved survival in the same TMA.4 ,6 ,32 CD16 and MPO are expressed in a subset of HLA-DR- CD15+ CD66b+ myeloid cells, mostly including activated neutrophils.4 ,6 Indeed, numbers of IL-17+ cells also correlated with those of infiltrating polymorphonuclear cells (see online supplementary figure S1D).
We wondered whether the positive impact of these populations might mask the negative prognostic significance of IL-17+ cells. However, also in cases stratified for CD16, MPO or CD8 expression, no effect of IL-17 on survival was found (see online supplementary figure S2).
CRC-infiltrating IL-17+ cells consist of Th17 cells
We next assessed phenotypes of CRC-infiltrating IL-17+ cells, in freshly isolated clinical specimens, by flow cytometry (figure 2). IL-17 production was exclusively observed within CD3+ T cells, whose large majority expressed CD4, CD45RO and CCR6 (figure 2A, B). Interestingly, 27±24% of infiltrating IL-17+ cells also expressed Foxp3 (figure 2A, B). Sporadically, IL-17+ CD8+ T cells were detected (figure 2A, B).

Colorectal cancers (CRC)-infiltrating interleukin (IL)-17+ cells are polyfunctional Th17. Single cell suspensions from freshly excised clinical specimens of CRC and corresponding tumour-free colonic mucosa (Ctr) and peripheral blood mononuclear cells from healthy donors (PBMC HD) or patients with CRC (PBMC Pz), were incubated with phorbol 12-myristate 13-acetate (PMA)/ionomycin/brefeldin for 5 h. Surface staining for specific cell population markers and intracellular staining for Foxp3 and cytokines was then performed. (A) Representative flow cytometric analysis of CRC infiltrates stained for IL-17 and the indicated cell-specific markers. Tumour-infiltrating cells are gated based on physical parameters, as defined by analysis of autologous PBMC. (B) Frequencies of cells positive for the indicated markers within gated IL-17+ cells. Means are indicated by bars. Numbers of samples evaluated for each marker are indicated in parentheses. (C) Frequencies of IL-17+ cells on gated CD3+ T cells obtained from PBMC HD or PBMC CRC and single cell suspensions from freshly excised clinical specimens of Ctr and corresponding CRC. Means are indicated by bars. Numbers of samples evaluated for each marker are indicated in parentheses. Statistical significance was assessed by t test. (D) Representative flow cytometric analysis of intracellular cytokine staining on gated CRC-infiltrating CD3+CD4+ cells. (E) Frequencies of cells positive for the indicated cytokines gated on CD3+CD4+ IL-17+ T cells. Means are indicated by bars. Numbers of samples evaluated for each cytokine are indicated in parentheses. GM-CSF, granulocyte-macrophage colony stimulating factor; IFN, interferon; TNF, tumour necrosis factor.
Large proportions of γδT cells and CD66b+ neutrophils (up to 24±5% and 13±9%, respectively) were detectable, but only a minor fraction (<1%) of those cell types showed evidence of IL-17 production. CD56+NK, Vα24/Jα18TCR+NKT and CD3−CD127+ LTi-like cells were detected in limited numbers and did not include significant fractions of IL-17+ cells (figure 2A, B). Evaluation of absolute cell numbers confirmed that the large majority of IL-17+ cells were CD3+ cells (see online supplementary figure S3). Thus, CRC-infiltrating, IL-17-producing cells mainly consist of memory Th17.
Th17 frequencies were significantly higher in tumours than in corresponding healthy tissues or autologous PBMC, whereas no significant difference was observed between PBMC of patients with CRC and HD (figure 2C).
Importantly, a significant fraction of CRC-infiltrating Th17 also produced TNF-α, IL-21, IL-22, GM-CSF, IFN-γ, and IL-8 (figure 2D, E). Th17 clones expanded from CRC-infiltrating CD4+ T cells (CRC-Th17) also released TNF-α, IL-21, IL-22, GM-CSF, IFN-γ and IL-8, in addition to IL-17 (see online supplementary figure S4), thus indicating that CRC-infiltrating Th17 are polyfunctional effector cells.
CRC-Th17 mediate protumorigenic effects in an IL-17-dependent manner
The lack of association between IL-17-producing cells and unfavourable clinical outcome was an unexpected finding considering the suggested protumorigenic activity of IL-17.7 ,8
This prompted us to investigate the functional properties of CRC-Th17.
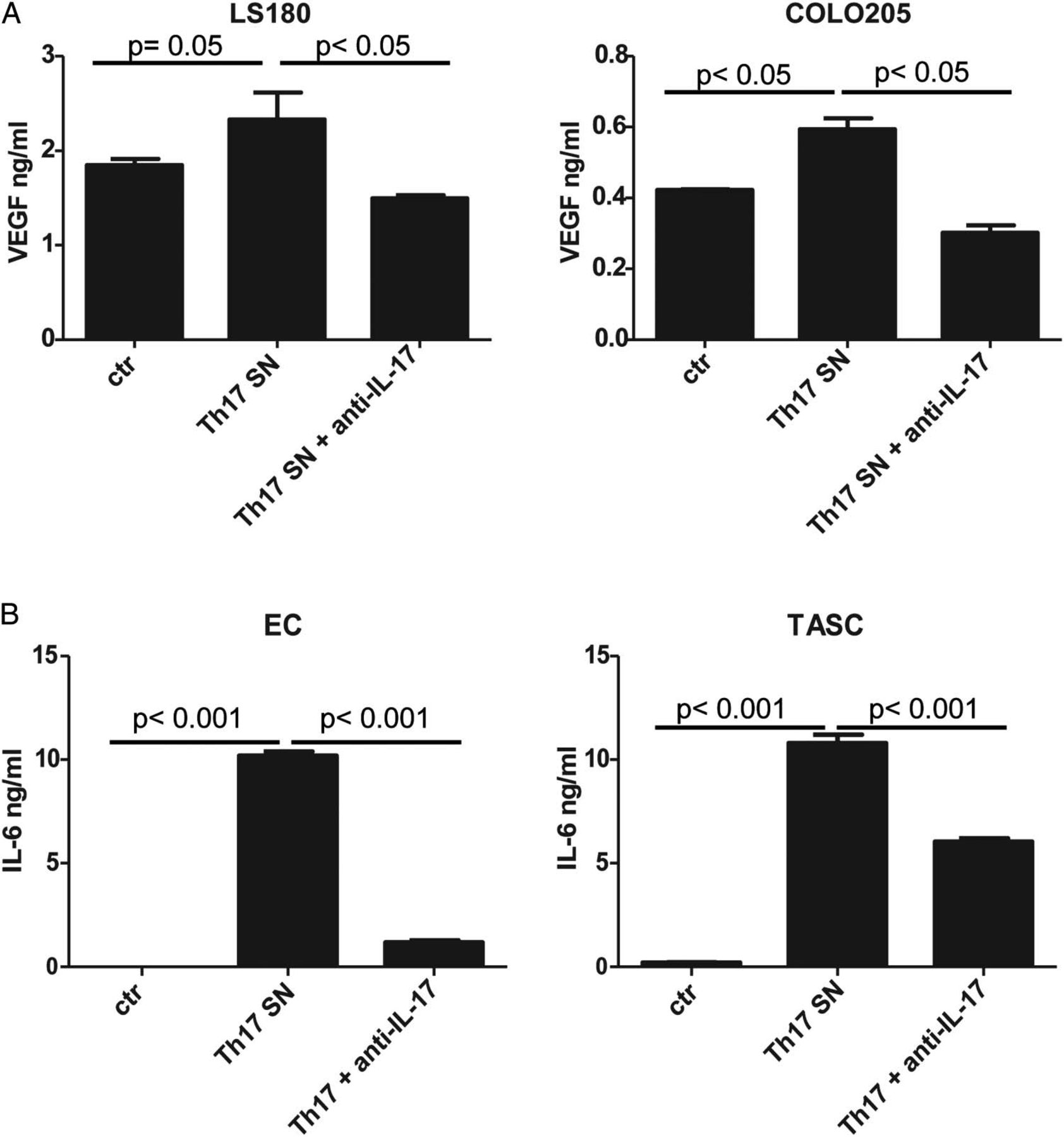
We conditioned CRC, endothelial cells (EC) and TASC with supernatants obtained from CRC-Th17 clones or bulk populations. No significant effects on cell proliferation were detected (see online supplementary figure S5). VEGF production by CRC cells was slightly increased by Th17 supernatants and it was inhibited upon IL-17 neutralisation (figure 3A and see online supplementary figure S6A). IL-6 release by EC and TASC was strongly induced by Th17 supernatants in an IL-17-dependent manner (figure 3B and see online supplementary figure S6B). Thus, IL-17 released by CRC-Th17 cells appears to mediate protumorigenic effects mainly by acting on EC and TASC.

Colorectal cancers (CRC)-Th17 mediate protumorigenic effects in an interleukin (IL)-17-dependent manner. Endothelial cells (EC), tumour-associated stromal cells (TASC) and CRC cell lines (LS180, COLO205) were conditioned for 24 h with CRC-Th17 supernatants untreated (Th17 SN) or pretreated with anti-IL-17 neutralising antibodies (Th17+ anti-IL-17). Vascular endothelial growth factor (VEGF) (A) or IL-6 (B) release was measured in culture supernatants by ELISA. Statistical significance was analysed by one-way analysis of variance. Data refer to experimental triplicates from two independent experiments performed with supernatants from two different clones. Ctr, healthy colonic tissue.
CRC-Th17 promote neutrophil recruitment and activation
We next investigated the molecular background underlying the association between Th17 and CD16+MPO+ myeloid cells or CD8+CTLs. The phenotypic analysis ruled out the possibility that neutrophils or CTLs could be the IL-17 producers. Alternatively, we investigated whether Th17 cells may recruit these cell populations into tumour tissues either directly or by eliciting chemokine release by stromal cells.33 ,34
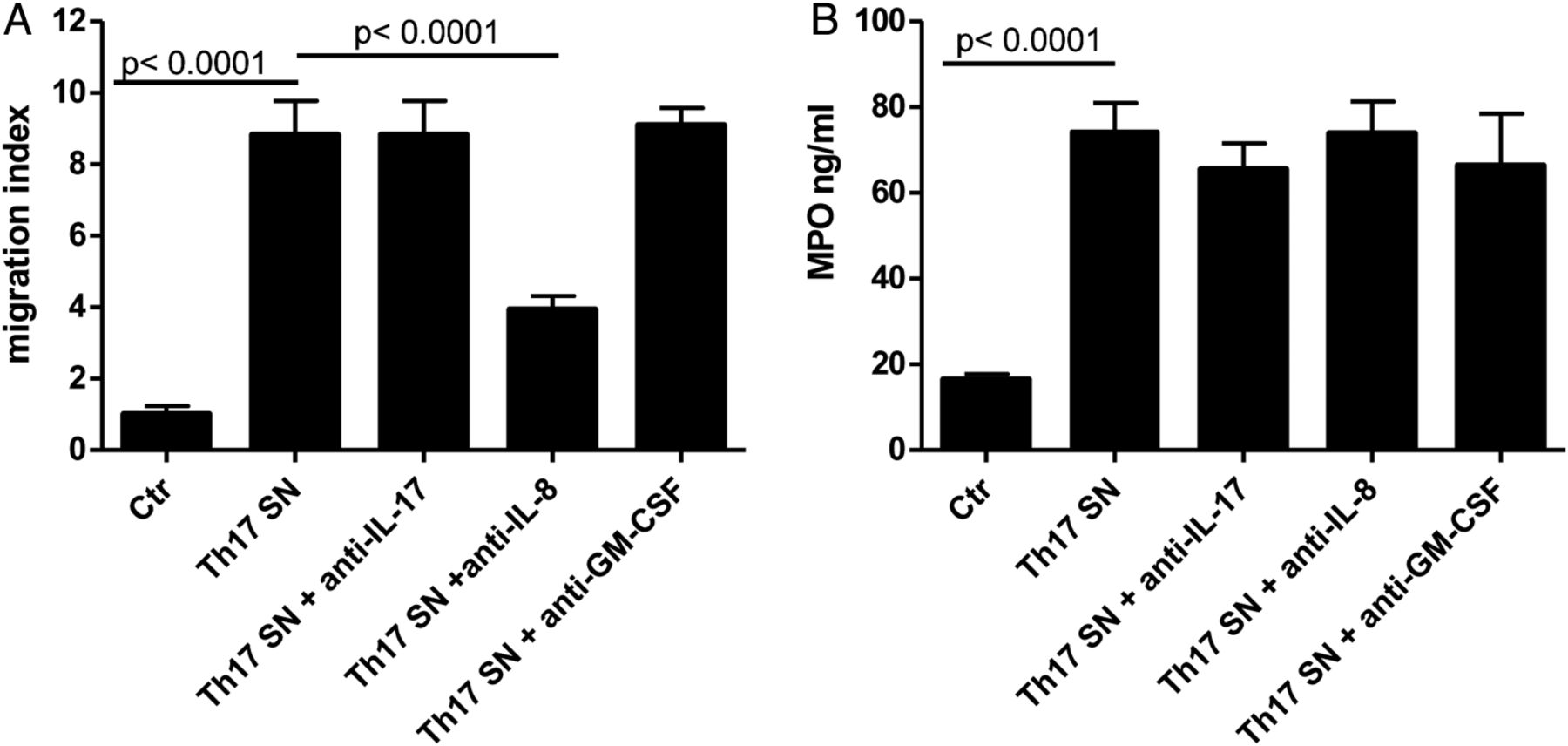
Neutrophils are known to migrate in response to IL-8. Indeed, supernatants from CRC-Th17 clones and bulk populations induced vigorous neutrophil migration in an IL-8-dependent manner (figure 4A and see online supplementary figure S7A, respectively), whereas other Th17 cytokines, including IL-17 and GM-CSF, appeared to play no role in this phenomenon (figure 4A and see online supplementary figure S7B). Interestingly, exposure of neutrophils to Th17 supernatants also significantly enhanced MPO release, independently of IL-17, IL-8 or GM-CSF (figure 4B and see online supplementary figure S7C). Thus, CRC-Th17 might directly promote neutrophil recruitment and activation into CRC.

Colorectal cancers (CRC)-infiltrating Th17 favour recruitment and activation of neutrophils. (A) Migration of neutrophils, purified from blood of healthy donors, towards control medium (Ctr), supernatants of Th17 clones expanded from CRC-infiltrating cells (Th17 SN) or Th17 supernatants pre-treated with anti-interleukin (IL)-17 (Th17 SN+ anti-IL-17), anti-IL-8 (Th17 SN+ anti-IL-8), or anti-granulocyte-macrophage colony stimulating factor (anti-GM-CSF antibodies) (Th17 SN+ anti-GM-CSF), was evaluated after 90 min incubation by flow cytometry. Data refer to experimental triplicates from three independent experiments performed with supernatants from three different clones. Means±SD are depicted. Statistical significance was assessed by one-way analysis of variance (ANOVA). (B) Myeloperoxidase (MPO) release by neutrophils exposed to control medium (Ctr), Th17 supernatants (Th17 SN) or Th17 supernatants pretreated with anti-IL-17, IL-8 or GM-CSF antibodies was assessed after 4 h incubation by ELISA. Means±SD are depicted. Data refer to experimental triplicates from three independent experiments performed with supernatants from three different clones. Statistical significance was assessed by one-way ANOVA.
CRC-Th17 favour CTL recruitment by triggering chemokine release from EC
In line with previous findings,35 circulating and tumour-infiltrating CTLs were found to express CCR5 and CXCR3 receptors (figure 5A), enabling them to respond to CCL5 and CXCL9/CXCL10, respectively.

Th17 cells favour recruitment of CD8+ T cells by triggering chemokine release from endothelial cells (EC). (A) Peripheral blood cells (PB, n=7) and tumour cell suspensions from patients with colorectal cancer (CRC) (n=12) were surface stained for CD8 in combination with the indicated chemokine receptors. Left panels: representative flow cytometric analysis. Right panels: percentages of CD8+ cells expressing the indicated chemokine receptors. Means±SD are depicted. (B) The expression of the indicated chemokine genes was assessed on CRC samples (n=21) by quantitative PCR. Expression levels relative to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) are depicted. (C) Endothelial, tumour and stromal cells were sorted from cell suspensions derived from CRC specimens by flow cytometry, based on CD31, EpCAM and CD90 expression, respectively. mRNA levels of the indicated chemokine genes were assessed in sorted cells by quantitative PCR. Gene expression levels relative to GAPDH are depicted. Data refer to analysis of one representative sample out of four. (D) Chemokine release by HMEC cells untreated (EC) or exposed overnight to recombinant interleukin-17 (rIL-17; 50 ng/mL) (EC+IL-17) or to Th17 clone supernatants (EC+Th17 SN), was measured by ELISA. Chemokine content in Th17 SN was also assessed as control. Data refer to experimental triplicates from three experiments performed with three different clones from one patient. Means±SD are depicted. Statistical significance was assessed by one-way analysis of variance (ANOVA). (E) Migration of CD8+ T cells, purified from peripheral blood mononuclear cells (PBMC) of healthy donors, towards supernatants of HMEC cells untreated (EC), or exposed to rIL-17 (50 ng/mL) (EC+IL17) or Th17 supernatants (EC+Th17), was assessed after 90 min of incubation by flow cytometry. Migration towards Th17 supernatants (Th17 SN) was also assessed as control. Data refer to experimental triplicates from three experiments performed with three different clones from one patient. Means±SD are depicted. Statistical significance was assessed by one-way ANOVA. (F and G) Chemokine contents (F) and cytotoxic T lymphocyte (CTL) chemoattraction capacity (G) of supernatants from HMEC cells untreated (EC), exposed overnight to Th17 supernatants (EC+Th17 SN), or exposed to Th17 supernatants pretreated with anti-IL-17 (EC+Th17+anti-IL-17) or anti-tumour necrosis factor α (anti-TNF-α) antibodies (EC+Th17+anti-TNF-α). Data refer to three experiments performed with supernatants of Th17 bulk populations derived from three different samples. Means±SD are depicted. Statistical significance was assessed by one-way ANOVA.
These chemokines were all expressed on whole CRC tissues (figure 5B). Upon sorting of tumour, stromal and EC from specimens, CCL5 and CXCL10 were expressed at the highest level within the endothelial compartment (figure 5C), whereas no CXCL9 was detected. EC exposure to CRC-Th17 supernatants strikingly enhanced their ability to release CCL5 and CXCL10 (figure 5D) and to promote CTL migration (figure 5E). Notably, these effects were not elicited by IL-17, nor were they inhibited after IL-17 neutralisation, but instead appeared to depend on Th17-derived TNF-α (figure 5F, G and see online supplementary figure S8). Thus, Th17 may favour CTL recruitment into CRC by triggering chemokine release from tumour-associated EC.
CRC-Th17 directly attract CTLs in vitro
Intriguingly, we observed that supernatants from CRC-Th17 bulks and clones significantly induced CTL migration also in the absence of EC (figures 5E, 6A and see online supplementary figure S9A), thus suggesting that Th17 may release chemoattractants directly acting on CTLs.

Th17 cells directly attract cytotoxic CD8+ T cells. (A) Migration of CD8+ T cells towards control medium (Ctr) or supernatants of two different colorectal cancer (CRC)-Th17 clones (Th17.15 SN and Th17.71 SN), was assessed after 90 min incubation by flow cytometry. (B) Chemokine release by Th17 clones (Th17.15 and Th17.71), activated with plate bound anti-CD3 and soluble anti-CD28 antibodies for an overnight period, was measured by ELISA. Means±SD of experimental triplicates are depicted. (C) Migration of CD8+ T cells towards Th17 clone supernatants depleted of CCL5 (Th17 SN−CCL5), CCL20 (Th17 SN−CCL20) or both (Th17 SN−CCL5−CCL20), relative to control (migration towards untreated Th17 supernatants). (D) Migration of CD8+ T cells towards Th17 clone supernatants (Th17 SN) or towards low or high doses of recombinant CCL20 (300 and 1000 ng/mL, respectively) and CCL5 (60 and 200 ng/mL, respectively). (A–D) Means±SD from experimental triplicates are depicted. One representative experiment out of two is shown. Statistical significance was assessed by one-way analysis of variance (ANOVA). (E) Representative flow cytometric analysis of CRC infiltrates stained for CD8 and CCR6 markers (left panel). Frequencies of CCR6+CD8+ T cells were measured by flow cytometry in peripheral blood mononuclear cells of healthy donors (PBMC HD n=7) or patients with CRC (PBMC Pz n=12) and in single cell suspensions from freshly excised clinical specimens of tumour-free colonic mucosa (Ctr) and corresponding CRC (n=12). Means are indicated by bars. Statistical significance was assessed by t test. (F) Left and middle panels: representative flow cytometric analysis of CRC-infiltrating CD8+ T cells stained for CCR6, CCR5 and TIA-1. Right panel: frequencies of CCR6+ CCR5+ cells within TIA-1- or TIA-1+ CRC-infiltrating CD8+ T cells (n=7).
In CRC-Th17 supernatants we detected significant amounts of CCL5, in addition to CCL20, a known Th17-derived chemokine (figure 6B and see online supplementary figure S9B). CTL migration was marginally impaired by CCL5 depletion, whereas it was markedly reduced upon CCL20 removal (figure 6C). However, CCL5 increased CTL migration to low CCL20 concentrations (figure 6D), thus revealing a synergism between these two chemokines. CCR6, the receptor for CCL20, was expressed on a subset of CRC-infiltrating CTLs (figure 6E). Frequencies of CCR6+CTLs were significantly higher in CRC than in autologous control tissues or PBMC (figure 6E). Importantly, most CCR6+CTLs (up to 62±10%) also expressed CCR5 (figure 6F). Furthermore, CCR5+CCR6+CTLs were positive for TIA-1 (figure 6F), a cytotoxic granule-associated, RNA-binding protein whose expression by CRC-infiltrating CTLs is associated with survival advantage.28 Thus, through CCL5 and CCL20 CRC-Th17 target highly cytotoxic T cells.
CRC-Th17 cells promote CTL recruitment into CRC tissues
We next assessed whether Th17-mediated effects might be relevant for CTL recruitment into tumours.
We engineered tridimensional tumour-like tissues by culturing HT29 cells on collagen scaffolds in perfused bioreactors31 ,36 (see online supplementary figure S10A, B). CRC-Th17 were added into the system under perfusion, allowing their localisation in proximity of tumour nests (see online supplementary figure S10C) and were subsequently activated (figure 7A). Perfusion was then stopped and CTLs were applied. After an overnight incubation, tumour infiltration by CTLs was evaluated (figure 7B, C and see online supplementary figure S10D). CTLs were found in tumour-like tissues, in proximity of tumour cells (figure 7B) and their frequencies were significantly higher in the presence of activated CRC-Th17 (figure 7C).

Th17 cells promote recruitment of cytotoxic CD8+ T cells into colorectal cancer (CRC) tissues. (A) CRC-Th17-infiltrating engineered tumour tissues (see online supplementary figure S10) were left untreated or were activated by adding CytoStim to the perfusion medium. Culture media were collected 20 h later and chemokine contents were assessed by ELISA. Statistical significance was assessed by Mann–Whitney test. (B and C) Three hours after Th17 activation, the perfusion in the bioreactor was stopped and CD8+ T cells were added to the system. After an overnight period, scaffolds were removed and tumour infiltration by CD8+ T cells was evaluated by immunofluorescence analysis upon staining with CD4- and CD8-specific antibodies (B) and by flow cytometry upon staining of single cell suspensions with EpCAM-, CD4- and CD8-specific antibodies (C). Percentages of CD8+ cells in tumour tissues infiltrated by resting or activated Th17 cells are reported. Means±SD from three experimental replicates performed with one Th17 clone are depicted. Statistical significance was assessed by Mann–Whitney test. (D) CSFE+CD8+ T cells were adoptively transferred in tumour bearing mice alone or together with equal numbers of CRC-Th17 (four mice/condition). Absolute numbers of CD8+CSFE+ T cells were evaluated by flow cytometry upon staining of tumour and Th17 cells with anti-EpCAM and anti-CD4 antibodies, respectively. Statistical significance was assessed by Mann–Whitney test.
Accordingly, when CTLs were adoptively transferred in tumour-bearing mice together with Th17, significantly higher numbers of tumour-infiltrating CTLs were detected as compared with mice transferred with CTLs alone (figure 7D). Thus, CRC-Th17 directly attract CTLs into tumours.
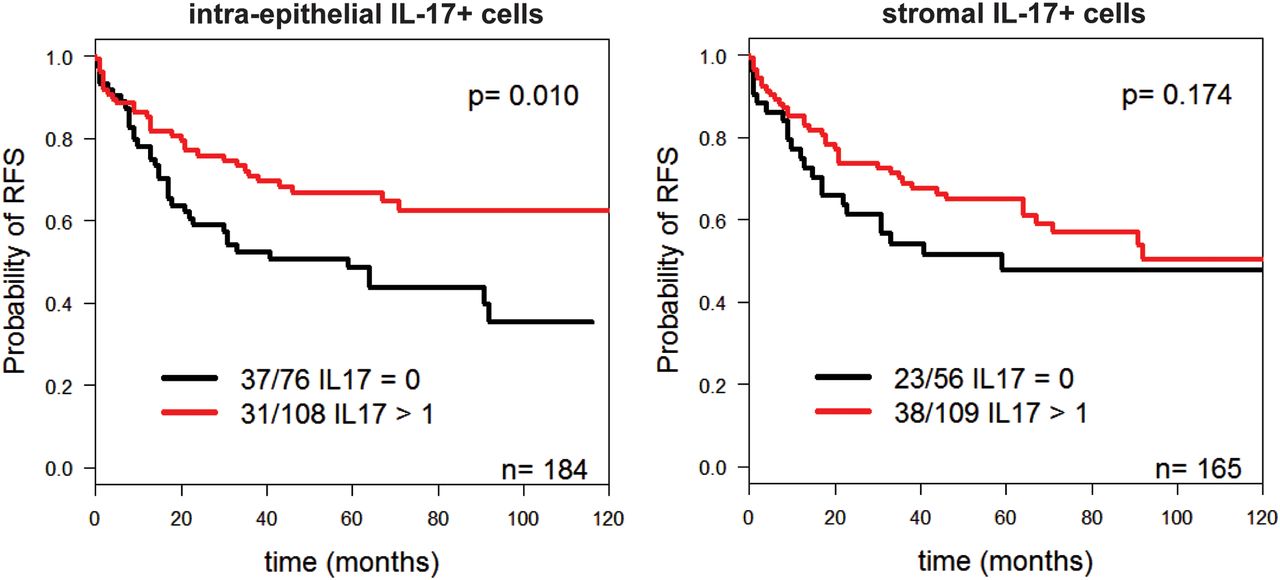
These findings suggested that CRC-Th17 localised into the tumour nests might play a beneficial role by recruiting CTLs close to tumour cells. Indeed, when we evaluated the prognostic significance of IL-17+ cells according to their localisation, the presence of intraepithelial, but not of stromal IL-17+ cells, positively correlated with absence of local recurrence and prolonged relapse-free survival (see online supplementary table S2 and figure 8).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Intra-epithelial localisation of interleukin (IL)-17+ cells correlates with prolonged relapse-free survival (RFS) survival. Kaplan–Meier curves illustrating RFS probability according to infiltration by IL-17+ cells within the epithelial (left panel) or stromal (right panel) compartment. Numbers of deaths/total cases within each category are indicated. Statistical significance was assessed by log-rank test.
Discussion
The goal of our study was to investigate the clinical relevance of CRC-infiltrating, IL-17-producing cells in a large patient cohort and to characterise in detail their phenotypes and functional properties. Upon analysis of >1000 primary CRC cases, we found that CRC-infiltrating IL-17+ cells are not themselves predictive of patient survival but, their prognostic significance rather appears to depend on their localisation within CRC tissue. Indeed, intraepithelial, but not stromal IL-17+ cells, were associated with improved prognosis. Ex vivo analysis showed that tumour-infiltrating, IL-17-producing cells mostly consist of polyfunctional Th17, releasing a spectrum of cytokines and chemokines, in addition to IL-17. Through IL-17, CRC-Th17 triggered the release of protumorigenic factors mainly by tumour-associated stroma. However, by releasing additional chemokines, they also appeared to contribute to the recruitment of beneficial effector cells into tumours. In particular, through IL-8 they promoted the recruitment of CD16+ MPO+ neutrophils. Most importantly, through CCL5 and CCL20, they attracted cytotoxic CTLs into tumour nests. Accordingly, in our patient cohort, CRC infiltration by Th17 cells correlated with that by neutrophils and CTLs.
Based on these findings, the lack of association between total numbers of Th17 cells and prognostic significance is not surprising. Indeed, the potentially negative impact of IL-17 is probably counterbalanced by recruitment of beneficial effector cells. In previous reports, however, an association between total IL-17+ infiltrates and unfavourable clinical outcome was observed.7 ,8 Although a few differences in the protocol used in our study as compared with others must be acknowledged (ie, in our work IL-17 positivity was assessed by three independent observers, whereas in the study by Tosolini et al8 the TMA staining was quantified by image software), the discrepancy between ours and previous studies probably relies on the different numbers of cases evaluated (527 and <2008 vs 1148). Notably, we obtained comparable results upon TMA staining with two different IL-17-specific antibodies.7 ,8
Importantly, our study reveals a previously unrecognised positive prognostic significance of intraepithelial Th17 cells, suggesting a critical contribution of these cell subsets to the recruitment of CTLs and neutrophils into tumour nests.
IL-17-production by CD4+ cells has been previously reported in CRC.37–39 However, whether other cell types may also contribute to IL-17 release within CRC tissues remained to be assessed. We observed that most IL-17+ cells are CD3+ CD4+ T cells, expressing CD45RO and CCR6 and including a subset of Foxp3+ cells. In contrast, no significant IL-17 production was seen within other cell subsets, including CTLs and innate lymphoid and myeloid cell populations.
IL-17-producing Foxp3+ cells were previously described within CRC tissues,37 ,38 ,40 but their functional role remains unclear. Remarkably, CRC infiltration by Tregs correlates with prolonged patient survival.2 ,3
A recent study reported γδT cells as a major IL-17 source in human CRC.41 We also observed considerable percentages of γδT cells within CRC infiltrates, but only a negligible fraction of them showed IL-17 production ability. However, the patients examined in the study by Wu et al41 and in our study belong to two different ethnic and geographical groups (ie, China and Switzerland, respectively), therefore probably differing in the composition of their gut flora.42 This may be of relevance when considering cell populations, such as γδT and Th17 cells, whose function is modulated by defined microbiota.22 ,43 ,44 Thus, this discrepancy may be due to the presence, in the patients evaluated, of distinct gut microbial species, possibly driving preferential expansion of IL-17-producing γδT or Th17 cells.
The ability of Th17 to recruit effector cells appears to be dependent on their capacity to release cytokines other than IL-17. Indeed, Th17 supernatants promoted neutrophil recruitment in an IL-8-dependent fashion and enhanced secretion of MPO, a lysosomal enzyme possibly involved in the mechanisms underlying the favourable effect of MPO+ neutrophils in CRC.4 In addition, CRC-Th17 supernatants activated tumour-associated EC to release CCL5 and CXCL10, attracting CTLs. Importantly, tumour-infiltrating Th17 proved capable of directly recruiting highly cytotoxic TIA-1+ CCR5+ CCR6+ CTLs through own production of CCL5 and CCL20. Interestingly, CCL5 and CCL20 appeared to synergise. Although CCL5 could not itself induce CTL migration, it could increase CTL response to low CCL20 concentrations.
In a melanoma mouse model, Th17 have been previously shown to favour tumour infiltration by CTLs, ultimately mediating tumour eradication.45 ,46 These in vivo models, however, could not discriminate between direct and indirect effects of Th17 on CTL recruitment. Therefore, to assess the relevance of the direct Th17-mediated effects on CTLs, we took advantage of an engineered tridimensional tumour tissue infiltrated by Th17 cells, in the absence of EC. Furthermore, we adoptively transferred human CTLs and Th17 in immunodeficient mice bearing human tumour xenografts. Strikingly, in both experimental systems, the presence of activated Th17 cells markedly enhanced CTL recruitment into the tumour.
These data suggest that Th17 cells may promote tumour infiltration by CTLs through a double axis: on the one hand, they might favour EC activation leading to CTL recruitment from the blood stream, on the other, they might directly guide the positioning of CTLs in proximity of tumour cells. Altogether our data reveal a positive contribution of Th17 cells to beneficial anti-tumour immune responses developing in CRC and underline their pleiotropic function resulting from the production of a broad spectrum of cytokines and chemokines beyond IL-17.
This may, at least partially, explain the discrepancies between results obtained from studies evaluating the effects of IL-17 or IL-17 signalling only and those examining functions of the whole Th17 cell subset.47–50
Our findings have important clinical implications. Indeed, the positive contribution of Th17 to anti-tumour immune responses should not be disregarded when developing new IL-17/Th17 targeted treatments in CRC, possibly resulting in impaired tumour infiltration by beneficial effector cells.
Acknowledgments
We thank Giulio C Spagnoli, Ed Palmer, Daniela Finke, Michael Heberer, Marco Lepore and Mariacarla Andreozzi for critical discussion and review of the manuscript; Thèrese J Resink for human microvascular endothelial cells (HMEC); Diego Calabrese for his advice on immunofluorescence studies; and Ivan Martin for his support with bioreactor-based cultures.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online supplement
Footnotes
Contributors FA and GI conceived and designed the experiments, analysed and interpreted results, obtained funding, wrote the manuscript. FA, MGM, CH, EC, VM, VG, and JH performed and analysed the experiments. XH, RAD, MZ, MA, MB and RR contributed to the conception of research and data collection. AL, IZ, LTe, LTo, PZ, SE-C and FT contributed to the conception of research, and collection, analysis and interpretation of data. IZ and SE-C performed the statistical analysis. EP contributed to the conception of research, experimental design, data analysis and interpretation and writing of the manuscript. DO contributed to the conception of research, data collection and financial support.
Funding Swiss National Science Foundation (SNF 310030-127490, PP00P3-133699, and PP00P3-159262), Freiwillige Akademische Gesellschaft Basel and Department of Surgery, Basel University Hospital.
Competing interests MGM is employed at Cellec-Biotek AG.
Patient consent Obtained.
Ethics approval Ethikkomission Beider Basel/Ethik Kommission Nord-West Schweiz (EKBB/EKNZ).
Provenance and peer review Not commissioned; externally peer reviewed.